Epifyttiske alger på Laminaria hyperborea (Gunn.) Foslie kartlagt langs en dybdegradient fra Skipsholmen, mørekysten.
|
|
|
- Endre Thomassen
- 4 år siden
- Visninger:
Transkript
1 Epifyttiske alger på Laminaria hyperborea (Gunn.) Foslie kartlagt langs en dybdegradient fra Skipsholmen, mørekysten. av Hanne Vigander
2 FORORD Denne oppgaven er gjennomført ved avdeling for marin botanikk, Biologisk Institutt, Universitetet i Oslo. Oppgaven er en del av et større tareskogsøkologiprosjekt. Prosjektet har studert tareskogen og dens økologi i området rundt Finnøy på mørekysten. En stor takk til professor Stein Fredriksen som har vært min gode veileder gjennom hele oppgaven! Takk Stein, jeg er veldig takknemlig for all hjelp du tålmodig har gitt. Takk også til professor Jan Rueness som alltid har vært tilgjengelig for faglige spørsmål og hjelp på laben. Takk til seniorforsker Hartvig Christie som sammen med Stein Fredriksen hjalp til med innsamlingen av materialet jeg har arbeidet med. Takk til professor Tom Andersen for hjelp med programvaren R. Din hjelp har spart meg tid og frustrasjoner. Takk til familien for god hjelp til korrekturlesning. Til slutt vil jeg takke alle på avdelingen for godt humør og muntre kommentarer som gjør det til et hyggelig sted å være! Hanne Vigander Oslo, 6. august 2007 i
3 SAMMENDRAG Hensikten med oppgaven har vært å kartlegge epifytter på stipes og hapter av Laminaria hyperborea (Gunn.) Foslie langs en dybdegradient. Til sammen ble 25 planter samlet inn 6. juni 2006 ved Skipsholmen utenfor Finnøy på mørekysten. Denne lokaliteten er preget av høy eksponeringsgrad og høy artsrikhet. Fem planter Laminaria hyperborea ble samlet fra hvert av fem dyp: 5, 10, 15, 20 og 25 meters dyp. Plantene ble målt og aldersbestemt. Det ble også foretatt målinger av lysinnstråling og temperatur på 3 dybder. Fire av de fem plantene fra hvert dyp ble systematisk gjennomgått, og epifyttene identifisert så langt mulig. De vanligste algene viste seg å være Phycodrys rubens, Ptilota gunneri, Polysiphonia stricta, Bonnemaisonia hamifera og Rhodochorton purpureum. Totalt ble det funnet 50 taxa epifytter fordelt på 38 rødalger, 9 brunalger og 3 grønnalger. Nullhypotesen, ingen signifikant forskjell på antall arter epifytter mellom dypene, har på bakgrunn av statistiske analyser blitt forkastet. Det ble registrert færrest antall arter på 25 meters dyp. Her var det signifikant færre arter enn på alle andre dyp unntatt 5 meter. Flest arter ble registrert på 15 meters dyp, men allikevel ikke signifikant flere arter enn på 10 og 20 meters dyp. Det viste seg at de fleste arter var til stede langs mer eller mindre hele transektet. Det er kun få arter som viser en sonering. Eksempler er Palmaria palmata som hovedsakelig er funnet på 5 meters dyp. Pterothamnion plumula og Brongniartella byssoides har jeg hovedsakelig funnet i den andre enden av transektet, fra 15 meter og nedover. Noen arter er også kun funnet på de midtre dypene. De gjennomførte målingene viser at det er små forskjeller i temperatur langs transektet. Det er også små forskjeller i lysmengde som trenger gjennom dekkvegetasjonen (canopy) på de forskjellige dypene. Lys og temperatur er sannsynligvis derfor ikke de viktigste forklaringsvariablene for forskjellene i antall arter mellom dyp. Hva som er de viktigste årsakene til variasjonen er fortsatt ubesvart. ii
4 INNHOLDSFORTEGNELSE FORORD... i SAMMENDRAG... ii 1 INNLEDNING Generelt om stortare, Laminaria hyperborea (Gunn.) Foslie og epifytter Gradienter Betydningen av epifyttdiversitet Målsetning MATERIALE OG METODER Stedsbeskrivelse Feltarbeidet Laboratoriearbeidet RESULTATER Kvantitativ del Antall arter Biomassedata Lysdata Temperaturdata Forsøk på å forklare variasjon i antall arter Systematisk del PLANSJER DISKUSJON Sammenligning med Sørlie Sammenligning med andre undersøkelser Diskusjon rundt variasjon i artssammensetning med dypet Usikkerhet rundt aldersbestemmelse Andre kommentarer Avsluttende konklusjoner REFERANSER VEDLEGG iii
5 1 INNLEDNING 1.1 Generelt om stortare, Laminaria hyperborea (Gunn.) Foslie og epifytter En epifytt er en organisme som vokser på overflaten av en plante eller en alge (Graham & Wilcox, 2000). Det ligger ikke i dette nødvendigvis en form for parasittisme, organismen benytter kun planten/algen som substrat å vokse på. Det betyr ikke at den ikke kan ha negativ betydning for vertsplanten (den vil for eksempel som regel skygge for lys eller gjøre vertsplantens opptak av næringsstoffer mindre effektivt). I denne oppgaven vil jeg kun se på den fastsittende floraen på stipes og hapter av stortare. Jeg vil ikke skille mellom hvorvidt planten vokser som en primær epifytt (dvs. sitter direkte på stipes) eller om den vokser som sekundær epifytt (dvs. vokser på en plante som igjen vokser på stipes). Dette fordi jeg er interessert i hele det floristiske påvekstsamfunnet på stortare og hvilke arter som er til stede. Det er for eksempel uten betydning for høyere trofiske nivåer hvorvidt arten er primær eller sekundær epifytt, det er tilstedeværelse eller ikke som er avgjørende. Stortarestilkens vekstsone sitter i overgangen mellom stipes og lamina. Den yngste delen av stipes nærmest lamina er glatt, men etter hvert får den vorteaktige utvekster og blir ru. Når dette skjer, får epifytter generelt lettere feste. Det er derfor en rik både flora og fauna av epifytter på stortarestilken. Likeledes er det et rikt både dyre- og planteliv i tilknytning til hapterene. Både flora og fauna er i sublittoralsonen begrenset av plass. Vanlig voksetetthet for stortare er ca 10 planter pr kvadratmeter (Christie et al., 1998). Stortaren utgjør derfor en betydelig økning av substrat som fastsittende organismer kan slå seg ned på. Stortaren med epifytter utgjør igjen et habitat for mobil fauna. Christie et al. (2003) fant i gjennomsnitt individer mobil fauna (særlig amfipoder og gastropoder) i tilknytning til hver stortareplante. 1
6 1.2 Gradienter I denne oppgaven har jeg sett på artssammensetning av epifytter langs en dybdegradient. En dybdegradient kan deles opp i flere gradienter av abiotiske faktorer som alle har betydning for hvor makroalgene kan vokse. Den mest åpenbare (og kanskje den mest studerte) gradienten er lysgradienten. I denne sammenhengen menes det fotosyntetisk aktive lyset (PAR). Lyset svekkes som kjent eksponensielt med dypet. Den nedre dybdegrensen for hvor en art vokser er derfor i stor grad bestemt av artens grad av lyskrav (Lüning, 1990 s. 5-7). Det er ikke bare dybden som avgjør hvor mye lys epifyttene får, hvor mye dekkvegetasjonen (canopyen) skygger spiller også inn, se avsnitt 4.3. Også andre sider av innstrålingen kan ha betydning for sonering. I hvor stor grad en art kan tåle UVB-stråling kan påvirke artens øvre voksegrense (Bischof et al., 1998). Alger har i likhet med andre planter mekanismer for å beskytte seg mot UVB-stråler. Stoffer som inngår i disse, finnes i større grad hos arter som vokser i litoralsonen enn skyggetilpassede mer dyptvoksende alger (Larkum & Wood, 1993). I følge Larkum og Wood (1993) er det enighet om at skadelige doser UVB-stråler kan trenge ned til 10 meters dyp. Dermed er også eksponering for UVB-stråler en faktor som kan forklare sonering av artene langs en dybdegradient. På et sted som ligger eksponert til vil det mekaniske stresset plantene utsettes for ved overflaten være betydelig. Turbulens og det mekaniske stresset avtar med dypet. Stresset vannbevegelsen påfører algene varierer ikke bare med hvor værhard lokaliteten er ved overflaten, men også forholdene under vann. For eksempel vil vannbevegelsen være kraftigere der kysten er bratt enn der det er langgrunt (Lewis, 1964 s. 30). For å kunne vokse eksponert kreves enkelte tilpasninger, for eksempel et sterkt festeorgan. Det er også en fordel med tallus som gir minst mulig risiko for å bli ødelagt eller revet bort av de turbulente vannmassene. Mulige tilpasninger til dette er å være liten eller ha en fasong som gir minst mulig turbulens (Lewis, 1964 s. 277). 2
7 Summen av disse abiotiske gradientene vil påvirke antall epifyttiske arter pr. tareplante. Hvor mange arter epifytter som er til stede, og særlig hvilke arter som er til stede, får igjen konsekvenser for de neste nivåene i næringskjeden. 1.3 Betydningen av epifyttdiversitet Hvordan epifytter påvirker økosystemet de er en del av har vært studert av mange. For eksempel har Parker et al. (2001) studert hvordan plantediversitet og artssammensetning kan påvirke epifaunaen. De fant at lite av variansen i den bevegelige epifaunaens diversitet kan forklares av plantediversitet. Derimot konkluderer de med at samfunnsstrukturen i faunaen er mer påvirket av spesielle egenskaper hos plantene, dvs. hvilke plantearter som er til stede, snarere enn hvor mange plantearter som er til stede. Studien ser ikke spesifikt på epifytter, men har i sine eksperimenter sett på makroalger med forskjellig struktur (Gracilaria, Ceramium og Ulva) og to arter sjøgress. Norderhaug (2004), derimot, har sett på amfipodetetthet på fire forskjellige rødalgeepifytter. Han konkluderte med at næringsverdi av epifyttene ikke forklarer forskjeller i amfipodetetthet. Viktigere er epifyttens strukturelle kvaliteter da forskjellige farger og strukturer kan spille en rolle både som beskyttelse mot predasjon og som beskyttelse mot fysisk eksponering. De omtalte studiene viser at det ikke bare er den totale epifyttbiomassen som spiller en rolle for samfunnets diversitet, men også hvilke arter som er til stede. Artskunnskap og kartlegging av artssammensetning er derfor viktig for å studere økosystemene langs kysten. Kunnskap om stedsspesifikk artssammensetning er også viktig for å fange opp endringer over tid. Kartlegging av arter er derfor også viktig i et konserveringsperspektiv. 1.4 Målsetning Målsetningen med oppgaven har vært å systematisk kartlegge epifytter funnet på stortare langs en dybdegradient på den beskrevne lokaliteten. Ettersom få har sett på endringer av 3


8 epifyttsammensetning med dypet, har hensikten vært å kartlegge eventuelle endringer i antall arter og artssammensetning langs gradienten. Nullhypotesen for dette arbeidet har vært at det ikke eksisterer en forskjell i gjennomsnittlig antall arter mellom dypene. Ettersom fokus på denne oppgaven har vært artssammensetning langs en dybdegradient har det ikke blitt gjort flere innsamlinger på andre steder eller til andre tider på året for sammenligning. 2 MATERIALE OG METODER 2.1 Stedsbeskrivelse Undersøkelsen er gjort på tare samlet inn fra sørspissen av Skipsholmen utenfor Finnøy på mørekysten, se figur 1. GPS-koordinater for lokaliteten for innsamlingen er N62º E6º Skipsholmen ligger eksponert til, det er få skjær utenfor dette. Eksponeringsgraden er derfor høy. NIVA (Norsk Institutt for Vannforskning) har laget en modell for eksponering i området. I denne modellen ligger eksponeringsgraden på lokaliteten for innsamlingen midt i kategorien veldig eksponert. Ålesund Skipsholmen Figur 1: Kart som viser Skipsholmens plassering langs norskekysten Denne modellen er kompatibel med EUNIS systemet, slik at lokaliteten også vil være klassifisert veldig eksponert etter det europeiske klassifiseringssystemet. Rundt Skipsholmen viste det seg under innsamlingen at nedre voksegrense for stortare er på 25 meters dyp. Til sammenligning kan stortare i klart kystvann vokse helt ned til 36 meters dyp (Moen & Svensen, 2004 s. 32), men dette er ikke vanlig. 4
9 Generelt blir denne delen av kyststrekningen regnet for å være et artsrikt område. Her finner man lavere temperaturforskjeller mellom sommer- og vintertemperaturer enn man gjør lenger sør. I henhold til Sætre (1973) er maksimal sommertemperatur i Hustadvika (som ligger langs samme strekning på mørekysten) 13,7 grader i august og laveste vintertemperatur 4,6 grader i februar og mars. Dette kan være en av årsakene til at flere arter som er regnet som sjeldne i Norge opptrer her (se mer om dette i avsnitt 4.5). På denne strekningen finner man også en overlapp mellom sørlig utbredelse for nordlige arter og nordlig utbredelse for sørlige arter som bidrar til artsdiversiteten. 2.2 Feltarbeidet Stortaren ble innsamlet 6. juni 2006 som en del av et større feltarbeid. Taren ble samlet ved dykking utført av Stein Fredriksen og Hartvig Christie. De hentet 5 tilfeldig valgte stortarestilker fra hvert av 5 dyp: 5 m, 10 m, 15 m, 20 m og 25 m. Stortaren ble skåret løs fra substratet, og på hvert dyp puttet i lerretposer slik at ikke epifytter skulle gå tapt under innsamling og transport. Tilbake på bryggen ble lamina fjernet og følgende målinger foretatt: stipeslengde og diameter av stipes både ved topp og bunn, hapterhøyde og - diameter. I tillegg ble stortarens alder registrert ved å telle årringer på et tverrsnitt ved overgangen mellom stipes og hapter (Kain, 1963). Stipes ble skrapt med kniv og kalkalgene skåret av slik at all påveksten ble samlet. Påveksten ble lagt på merkede flasker. Hver plante ble behandlet for seg. Hapter ble skåret i biter og lagt på flasker. Alle prøvene ble fiksert med formalin (2 %). Et alternativ til denne metoden kunne vært å dele stipes i biter og legge bitene på flaske. Ettersom hovedfokus på oppgaven er arter langs dybdegradienten, og ikke hvor på stipes artene vokser, ville ikke dette ha tilført oppgaven vesentlig informasjon. Dette alternativet ble derfor valgt bort av plass- og transporthensyn. Det ble satt ut lysloggere som logget lysinnstråling og temperatur langs transektet der prøvene ble hentet. Disse logget innstrålingen og temperatur hvert 10. minutt over 4 5
10 døgn. Brikkene ble satt ut på 3 dybder: øverst (ca. 5 meters dyp), på midten (ca. 10 meters dyp) og nederst (ca. 25 meters dyp) i transektet. På hvert dyp ble det festet en brikke over dekkvegetasjonen og en brikke lenger nede under dekkvegetasjonen. Brikkene var festet til en snor som ble forankret. Dessverre var den brikken som skulle måle lys over dekkvegetasjonen nederst i transektet montert opp ned. Dermed var det kun 5 brikker som logget korrekte lysdata og ikke Laboratoriearbeidet Tilbake i laboratoriet ble totalt fire av fem planter fra hvert dyp studert. Nytten av å gjennomgå den siste planten ved hvert dyp ble vurdert som lav, se figur 2 side 9. Denne ble derfor utelatt. Etter å ha vasket ut formalinen ble en og en prøve systematisk gjennomgått i lupe og lysmikroskop. Alle arter i prøven ble artsbestemt i den grad det lot seg gjøre. Et mindretall av artene ble funnet med reproduktive strukturer. Overvekt av små individer og fravær av reproduktive strukturer vanskeliggjorde, og i noen tilfeller umuliggjorde, artsbestemmelsen. Flere individer er derfor ikke bestemt helt til art. Ettersom prøvene var fiksert med formalin kunne jeg heller ikke benytte meg av dyrkningsforsøk eller krysningsforsøk i tilfeller der dette kunne vært metoder for å komme frem til sikrere artsbestemmelse. I enkelte tilfeller hadde jeg i arbeidet med artsbestemmelse behov for å farge cellekjerner hos et individ. Dette ble gjort ved å varme opp algematerialet i en haematoxylin-løsning etter Wittmanns metode (Wittmann, 1965). Kalkalger som ikke kunne bestemmes uten snitting ble lagt til side ved gjennomgang av prøvene for deretter å bli behandlet for seg til slutt. Disse ble først avkalket med saltsyre (1N). Deretter ble en frysemikrotom benyttet. Skorpen ble frosset fast i ferskvann og snitt på mellom 30 µm og 50 µm skåret. De fleste kalkalgene var små individer som falt av tarebiten den satt på ved avkalking. Dette gjorde at det var vanskelig å få snitt som kunne 6
11 benyttes til artsbestemmelse, og ofte manglet også snitt av konseptakler. Kun en liten andel av skorpene kunne derfor artsbestemmes. Ettersom et så lite utvalg av kalkalgene ble bestemt, har jeg valgt å holde dem utenfor i statistikken over antall arter pr. dyp. De er derfor ikke inkludert i oversikten i vedlegg 1, men omtales allikevel i avsnitt 3.2, resultater - systematisk del. Til artsbestemmelse er følgende hovedverk benyttet: i arbeidet med rødalgene har de 5 bindene i volum 1: Rhodophyta i serien Seaweeds of the British Isles vært viktige. Rosenvinge (1909) har vært mye brukt til bestemmelse av de uniseriate rødalgene. Generelt har også Norsk Algeflora av Rueness (1977) vært flittig benyttet. I tillegg har A handbook of the British seaweeds (Newton, 1931) og Meeresalgen von Helgoland (Kornmann & Sahling, 1977) vært benyttet som oppslagsverk. Disse nevnte verkene er benyttet til beskrivelsene av artene uten at det nødvendigvis er referert til dem ytterligere. Der annen litteratur er benyttet i tillegg eller i stedet er dette referert til i teksten. Algers systematikk er under stadig revisjon og endring etter hvert som nye molekylære undersøkelser foretas. Flere alternative systematiske inndelinger eksisterer. Jeg har valgt å følge systematikken fra i denne oppgaven. Algaebase gjør ikke noe forsøk på å sortere etter slektskap på ordensnivå. Ordnene er derfor i denne oppgaven også presentert i alfabetisk rekkefølge. Det finnes belegg av alle identifiserte arter, enten i form av corn-sirup mikroskopipreparater, herbarieark, bilder eller en kombinasjon av disse. Det er kun noen arter der individene har vært store nok til at det har vært noen mening i å presse planten for å lage herbarieark. De fleste arter er derfor dokumentert ved hjelp av bilder og mikroskopipreparater. Bildene ble tatt med et Nikon digitalkamerasystem for mikroskopi, modell DS-5M. Bildene som er tatt i mikroskop er tatt med en kjent forstørrelse og systemet tillater da at en skala legges på bildet. Denne har dessverre ofte blitt utydelig, og ny skala er derfor lagt over manuelt uten at det har latt seg gjøre å fjerne den automatisk genererte skalaen. Bildene som er tatt i lupe er tatt uten en kjent forstørrelse ettersom denne er trinnløs. Disse bildene mangler derfor dessverre skala. 7
12 Når alle prøvene var gjennomgått, ble samtlige prøver av stipes satt i tørkeskap og tørket slik at jeg fikk et mål på tørrvekt av biomasse epifytter. At dette skulle gjennomføres ble bestemt i etterkant av gjennomgangen av prøvene. Jeg har derfor ikke tatt hensyn til at biomassen skulle veies, hverken under innsamling eller gjennomgangen på laboratoriet. I tillegg hadde jeg ingen mulighet for å skille ut påvekst av bryozoer som Electra pilosa, som i mange prøver kunne være omfattende. Selv om biomassemålene på grunn av dette blir mer unøyaktige, gir de allikevel en idé om fordelingen av epifyttbiomassen med dypet. For hver prøve har jeg gjort en subjektiv vurdering av hvor vanlig hver art er i prøven. Jeg har benyttet en skala fra 1 til 4. Arten er tilegnet en verdi etter følgende vurdering: biomassemessig dominerende = 4, vanlig = 3, spredt = 2, enkeltfunn i prøven = 1. Graderingen er min subjektive vurdering og utført for å få et inntrykk av hvor viktig arten er på stedet. En fullstendig tabell finnes som vedlegg 1. Identifiserte taxa ble registrert pr. prøve pr. dyp slik at jeg til slutt kunne telle opp hvor mange taxa som gjennomsnittlig var til stede på hver plante på de forskjellige dypene. Standardavvik og konfidensintervall rundt dette ble beregnet i Excel. Dermed kunne jeg teste hvorvidt observerte forskjeller i antall arter på de forskjellige dypene var signifikant forskjellig på 99 % signifikansnivå. Vanligvis benytter man 95 % signifikansnivå. Fordi jeg her foretar multiple sammenligninger har jeg benyttet Bonferroni s metode som vil si at man korrigerer α i forhold til antall sammenligninger som gjøres (Crawley, 2002 s. 274). Ettersom jeg foretar 5 sammenligninger blir dermed α = 0,01. Fordi jeg har få prøver ønsket jeg å få resultatet av denne testen bekreftet av flere tester. Jeg har derfor i tillegg utført variansanalyse, se vedlegg 6 og Tukeys HSD-test for multiple sammenligninger, se vedlegg 5. Begge disse testene er utført i programvaren R. Jeg har også benyttet R til å lage en enkel regresjonsmodell for å se om det er mulig å si noe om hvilke variabler som forklarer variasjonen i antall arter epifytter med de dataene jeg har. Jeg har valgt først å teste en modell med samtlige variabler og interaksjoner for 8
13 så å fjerne den minst signifikante variabelen helt til alle variabler er signifikante (baklengs seleksjon). Denne metoden anbefales av Crawley (2002 s.452). Ettersom plass som nevnt er en begrensende faktor for epifytter ville jeg beregne overflate på stipes for å kunne teste denne variabelen i modellen. Stipesoverflate ble beregnet etter formelen: overflate = ( omkrets topp + omkrets bunn) høyde 2 Resultatene av beregningen finnes i vedlegg 3. Når man skal teste en regresjonsmodell, finnes en tommelfingerregel som sier at man ikke skal teste en modell med flere variabler enn antall datapunkter (n) / 3 (Crawley, 2002 s. 442). Med alle variablene jeg har, inkludert interaksjoner, overstiger jeg dette. Stipesoverflate og stipeslengde er sterkt korrelerte størrelser. For å redusere antallet variabler, laget jeg en modell med hver av dem. Modellene er ellers like. Deretter valgte jeg den beste av de to modellene basert på AIC (Akaike Information Criterion) (Crawley, 2002 s. 316). Dette viste seg å være modellen som inkluderer stipeslengde. 3 RESULTATER 3.1 Kvantitativ del De fleste arter ble oppdaget ved gjennomgang av første plante fra alle dyp. Ingen nye arter ble oppdaget ved gjennomgang av fjerde plante, se figur 2. Det ble derfor bestemt at det ikke var nødvendig å gå gjennom plante fem fra alle dyp da det ville tatt lang tid og sannsynligvis gitt lite ekstra informasjon. Antall observerte arter Plante Figur 2: Totalt antall observerte epifytter plottet mot antall gjennomgåtte planter fra hvert dyp. 9
14 3.1.1 Antall arter Totalt har jeg i løpet av gjennomgangen registrert 50 taxa (inkludert kalkalgene). Det er en stor overvekt av rødalger representert. Hele 76 % (dvs. 38 taxa) av de registrerte epifyttene er rødalger, 18 % (dvs. 9 taxa) er brunalger og 6 % (dvs. 3 taxa) grønnalger, se figur 3. To faktorer som varierer med antall arter på plantene er plantenes alder og stipeslengde. Stipeslengde og alder er igjen korrelerte. Figur 4, 5 og 6 viser sammenhengen mellom disse tre størrelsene i mitt materiale. Brunalger 18 % Grønnalger 6 % Rødalger 76 % Figur 3: grafisk fremstilling av fordelingen av epifytter mellom rødalger, grønnalger og brunalger A n t a ll a r t e r e p if y t t e r A n t a ll a r t e r e p if y t t e r Alder Figur 4: antall arter epifytter på en tarestilk plottet mot tarestilkens alder for de 20 gjennomgåtte plantene Stipeslengde (cm) Figur 5: antall arter epifytter på en tarestilk plottet mot tarestilkens lengde for de 20 gjennomgåtte plantene. 10
15 S t ip e s le n g d e ( c m ) Alder Figur 6: tarestilkens lengde plottet mot tarestilkens alder for de 25 samlede plantene. Figur 4 viser at det er en positiv sammenheng mellom antall arter og stortarestilkens alder. Figur 5 viser at det også er en positiv sammenheng mellom antall arter og stortarens stipeslengde. Begge deler er forventet. På en eldre stilk har epifyttene hatt lengre tid til å slå seg ned, og på en større stilk er det plass til flere individer slik at det er større sjanse for å finne flere arter på stilken. Figurene sier derimot ikke noe om årsakssammenheng. Ettersom stortarens alder og stipeslengde også er korrelert (figur 6), kan man heller ikke si noe om det er en eller begge faktorer som egentlig påvirker antallet epifytter. Som nevnt varierer antall arter epifytter fra plante til plante (figur 7). Færrest arter ble funnet på plante nr. 2 fra 25 meters dyp. Denne hadde kun 12 arter epifytter. Flest arter registrerte jeg på plante nr. 2 fra 10 meters dyp. Denne hadde 32 arter epifytter. Antall arter Plante 1 Plante 2 Plante 3 Plante meter 10 meter 15 meter 20 meter 25 meter Dybde Figur 7: Antall arter registrert pr plante. I denne oversikten er de skorpeformede holdt utenfor. 11
16 Antall arter varierer også veldig med dypet. Figur 7 gir et tydelig inntrykk av at det var færre arter på 5 meter og 25 meter enn på de tre midterste dypene. Dette bekreftes av figur 8: 35 Antall arter Figur 8: Gjennomsnittlig antall arter pr. dybde med konfidensintervall tegnet inn, α =0, meter 10 meter 15 meter 20 meter 25 meter Dybde Standardavvikene for antall epifytter varierer mellom dypene fra 1,41 planter (på 15 meters dyp) til 4,21 planter (på 20 meters dyp). Figur 8 viser at det i gjennomsnitt er signifikant færre arter på plantene på 5 og 25 meter enn på 10 og 15 meter. På 25 meter er det også signifikant færre arter enn på 20 meter. Ettersom konfidensintervallene for de tre midterste dypene overlapper, kan derimot ikke ett av dypene sies å ha signifikant flest arter. Tilsvarende kan det her ikke skilles mellom 5 meter og 25 meter når det gjelder hvilket dyp som har signifikant færrest arter. Ettersom jeg har få prøver har jeg villet være på den sikre siden og teste dette resultatet med en annen modell. Jeg har derfor utført en variansanalyse i R. Nullhypotesen, alle gjennomsnitt er like, må i følge analysen forkastes (p = 0, ), vedlegg 6. Deretter utførte jeg en Tukey HSD test ettersom dette er en av de mest konservative testene for multiple sammenligninger. Denne testen gir samme konklusjon som multiple sammenligninger justert med Bonferroni s metode, se vedlegg 5. 12
17 Størst variasjon innenfor ett dyp fant jeg på 20 meter, hvilket gir seg utslag i et større konfidensintervall på dette dypet enn for de andre dypene. Minst variasjon mellom plantene ble registrert på 15 meters dyp Biomassedata Den grafiske fremstillingen av tørrvekten (figur 9) viser at biomassen av epifytter på stipes også varierer med dypet. Klart minst biomasse ble registrert på 25 meters dyp. Her var samtlige individer små. Mest biomasse ble funnet på 10 og 15 meters dyp. Her fantes individer av flere arter, for eksempel Phycodrys rubens og Ptilota gunneri, som hadde vokst seg store relativt sett i forhold til individstørrelse generelt i mine prøver. Biomassedataene er kun benyttet videre som en forklaringsvariabel i modellen over variasjon i antall arter, se avsnitt Tørrvekt (gram) meter 10 meter 15 meter 20 meter 25 meter Dybde Plante 1 Plante 2 Plante 3 Plante 4 Figur 9: Tørrvekt av epifytter på stipes for alle 20 gjennomgåtte planter. Det er en mulighet for at biomassen for plante 2, 10 meter er feil. Se mer om det i avsnitt 4.5. Biomassedata finnes i vedlegg nr Lysdata Figur 10 viser at lysmålingen på grunt vann over dekkvegetasjonen (canopy) skiller seg ut fra de andre målingene i den forstand at vesentlig mer lys trenger ned hit enn til de andre lysmålerne. Av de andre 4 stedene det ble målt lysinnstråling disse dagene var det medium dybde over dekkvegetasjonen som fikk mest lys. På grunt vann under 13
18 dekkvegetasjonen ble det målt like lite lys som på dypt vann. 5m - over canopy 5m - under canopy 10m - over canopy 10m - under canopy 25m - under canopy Lys / lux til 9. juni Figur 10: Lysdata fra 5 dybder registrert på Skipsholmen hvert 10. minutt 6. til 9. juni Temperaturdata 5m - over canopy 5m - under canopy 10m - over canopy 10m - under canopy 25m - over canopy 25m - under canopy Temperatur/celsius 9,8 9,6 9,4 9,2 9 8,8 8,6 8,4 8, til 9. juni Figur 11: Temperaturdata fra 6 dybder registrert på Skipsholmen hvert 10. minutt 6. til 9. juni. 14
19 Skipsholmen ligger som tidligere nevnt eksponert til. Vannmassene er derfor hele tiden under omrøring. Dette er en viktig årsak til at temperaturmålingene som ble gjort viser minimale forskjeller i temperatur med dybden (se figur 11). Målingene gjort på grunt vann og medium dybde ligger generelt mellom 9 og 9,5 grader. Det dype vannet skiller seg ut ved å være litt kaldere, men bare marginalt. Her varierer målingene stort sett mellom 8,5 og 9 grader, dvs. en halv grad kaldere. Ettersom forskjellene i temperatur viser seg å være så marginale kan temperatur ikke være en faktor som forklarer forskjeller i artssammensetning på denne lokaliteten og dataene ble følgelig ikke benyttet videre Forsøk på å forklare variasjon i antall arter For å gjøre et forsøk på å finne forklaringsvariable for antall arter epifytter satte jeg opp følgende regresjonsmodell i programvaren R: lm(ant.arter ~ (dyp + alder + stipeslengde + biomasse)^2) Denne modellen tester de fire variablene dyp, alder, stipeslengde og biomasse, og samtlige interaksjoner mellom disse. Verdiene for antall arter finnes i vedlegg 1, verdiene for de andre variablene finnes i vedlegg 3. Når alle ikke-signifikante variabler er fjernet, står jeg igjen med modellen: lm(formula = ant.arter ~ dyp + alder + stipeslengde + biomasse + dyp:stipeslengde + alder:stipeslengde + alder:biomasse) For fullstendig utskrift av modellen, se vedlegg 6. I vedlegg 6 finnes også et plot av residualene som viser at disse er tilnærmet normalfordelte. Modellen viser at 6 variabler og interaksjoner er signifikante (alder er ikke signifikant, men er med fordi interaksjoner med alder er signifikante) i denne modellen. Disse sier derfor noe om variasjonen i antall arter. Til sammen forklarer variablene nesten 74 % av variasjonen. 15
20 3.2 Systematisk del I denne delen presenteres systematisk alle artene jeg har funnet og artsbestemt i løpet av gjennomgangen av prøvene. For hver art som presenteres angis nøkkelkarakterer, referanse til belegg, forekomst, fertile strukturer funnet i prøvene, og til slutt eventuelle kommentarer. Phylum RHODOPHYTA Klasse COMPSOPOGONOPHYCEAE Orden ERYTHROPELTIDALES Familie Erytrotrichiaceae Erythrocladia irregularis Rosenvinge Beskrivelse: Tallus består av krypende cellerekker som danner en enlaget skive på inntil 100 µm i diameter. Cellerekkene utløper fra et sentrum og kan danne alt fra en regelmessig, relativt rund skive til sprikende grener. Cellene er ovale og har en veggstilt kloroplast med pyrenoide. Endecellene er ofte gaffeldelte. Belegg: Plansje 1, bilde a) og b). Preparat 1 og 2. Forekomst: Arten er funnet epifyttisk på forskjellige arter, for eksempel Sphacelaria sp., Spermothamnion repens, og Polysiphonia fibrillosa. Den er funnet i prøver fra alle dyp. Fertile strukturer: Ikke funnet. Kommentarer: Arten er sannsynligvis vanligere i prøvene jeg har gjennomgått enn det mine registreringer skulle tilsi. Ettersom den kun kan ses i mikroskop, blir det litt tilfeldig hvor ofte man kommer over den i en prøve. Porphyropsis coccinea (J. Agardh ex Areschoug) Rosenvinge Beskrivelse: Kun et cellelag tykt tallus med blekt rosa farge. Cellene er små, kun 4-7 µm i diameter, og ligger i regulære rekker. Belegg: Plansje 1, bilde f). Preparat 3. Forekomst: Arten er funnet på alle dyp, og har i flere prøver vært veldig vanlig. Den har forekommet oftest på hapter, men har også vært funnet på andre rødalger på stipes. Fertile strukturer: Ikke funnet. 16

 og c). Preparat 4 og 5. Forekomst: Funnet i flere prøver på 10 meter og 15 meters dyp.")
21 Klasse FLORIDEOPHYCEAE Orden ACROCHAETIALES Familie Acrochaetiaceae Acrochaetium minimum F. S. Collins Beskrivelse: Tallus består av uniseriate, grenete filamenter som vokser endofyttisk i celleveggen hos blant annet arter av Polysiphonia. Filamentene er rikt forgrenete og danner et nettverk. Cellene er delvis sylindriske, 8-14 µm lange og 2-4 µm brede. Kloroplasten er parietal og danner en sylinder langs celleveggen. Monosporer er enten fastsittende på filamentene eller på encellete stilker. Belegg: Plansje 1, bilde b) og c). Preparat 4 og 5. Forekomst: Funnet i flere prøver på 10 meter og 15 meters dyp. Kun ett funn fra 20 meters dyp. Arten er i mine prøver med andre ord vanlig der Polysiphonia fibrillosa er vanlig. Fertile strukturer: Monosporangier funnet, se bilde c) på plansje 1. Kommentarer: Denne arten er sannsynligvis vanligere i mine prøver enn det jeg har kunnet dokumentere. På grunn av algens størrelse er den vanskelig å legge merke til, selv i mikroskop på lavere forstørrelser. Den blir ytterligere vanskelig å oppdage av at den har a) samme farge som vertsplanten. Ikke mange arter har så små celler og danner et nettverk i celleveggen på denne måten. Voksemåte og cellestørrelse var derfor vesentlige karakterer i artsbestemmelse av dette funnet. Figur 12 a) viser arten vokse i celleveggen av Polysiphonia fibrillosa. Denne figuren viser også monosporangium på vei ut (se pil). Mine prøver er fikserte, og observasjoner av kloroplastene er derfor noe usikre. Det man kan se av kloroplaster stemmer allikevel med beskrivelsen over, se figur b) 10 µm 10 µm Figur 12 a) A. minimum i celleveggen av Polysiphonia fibrillosa. Monosporangium merket med pil. b) Forgreningsmønster og veggstilte kloroplaster hos A. minimum 17
22 12 b). Mine observasjoner stemmer også godt overens med beskrivelse og tegninger av arten hos Rosenvinge (1909 s ) (beskrevet som Chantransia emergens). I følge litteraturen trives arten i Polysiphonia (Dixon & Irvine, 1977 s. 94). Etter at jeg ble oppmerksom på den, har jeg derfor lett etter den like mye i P. stricta som i P. fibrillosa. Leting i P. fibrillosa ga ofte resultater, men jeg fant den aldri i P. stricta. Heller ikke i de prøvene der den var til stede i P. fibrillosa. Rosenvinge beskriver derimot arten fra P. stricta (Rosenvinge, 1909 s.129). Det har lenge vært mange uavklarte spørsmål vedrørende systematikken rundt ordenen Acrochaetiales. Garbary går så langt som til å omtale familien Acrochaetiaceae som taksonomisk kaos (Garbary, 1978). Harper og Saunders (2002) foretok derfor molekylære analyser med det formål å belyse slektskap mellom Acrochaetiales og nærstående ordner, slektskap mellom de to grenene i Acrochaetiales (Acrochaetiales I og II, se Harper og Saunders (1998)) og kartlegge hvilke morfologiske karakterer som kan være nyttige for å skille mellom disse. På bakgrunn av de molekylære undersøkelsene, argumenterer Harper og Saunders (2002) for at ordenen Acrochaetiales burde begrenses til den opprinnelige Acrochaetiales I og at Acrochaetiales II burde plasseres i en egen, ny orden, Colaconematales. Videre beskriver forfatterne de morfologiske trekkene som skiller disse ordenene. Arter som reproduserer med monosporangier i Acrochaetiales skal ha en enkel, stjerneformet kloroplast lokalisert langs midtaksen. Arter i ordenen Colaconematales, derimot, skal ha en eller flere parietale plastider av varierende form, men aldri stjerneformet. Benytter man disse kriteriene virker det sannsynlig at A. minimum skulle ha vært plassert i Colaconematales. Arten er i Algaebase plassert i slekten Acrochaetium, men den har av andre tidligere blitt plassert i slekten Colaconema. Basert på Harper og Saunders (2002) undersøkelser av Acrochaetiales kan man stille spørsmålstegn ved Algeabases plassering av denne arten i Acrochaetium. 18
23 Rhodochorton purpureum (Lightfoot) Rosenvinge Beskrivelse: Tallus består av krypende og opprette uniseriate, grenete filamenter. Det opprette systemet blir inntil 1 cm høye og er µm brede. Hver celle inneholder flere kloroplaster. Disse kan være skiveformete til båndformete og mangler pyrenoide. Tetrasporangiene er korsdelte og sitter i knipper. Belegg: Preparat 6 og 7. Forekomst: Funnet i samtlige prøver med unntak av en prøve på 5 meter. Funnet både på stipes og på hapter, og er en av de vanligste artene i prøvene. Fertile strukturer: Tetrasporangier funnet. Kommentarer: I flere av funnene som jeg har bestemt til R. purpureum har jeg bare kunnet se en kloroplast i mange av cellene, mens det har sett ut som om noen celler i samme celletråd hadde flere kloroplaster. I følge Dixon og Irvine (1977 s. 107) har cellene først kun en kloroplast. Denne fragmenteres med alder til flere. Dette kan forklare min observasjon. Acrochaetium-lignende, ubestemte I samtlige prøver fantes det Acrochaetium-lignende individer som ikke lot seg bestemme grunnet for få karakterer. Orden BONNEMAISONIALES Familie Bonnemaisoniaceae Bonnemaisonia hamifera Hariot, Trailliella-fase Beskrivelse: Trailliella-fasen består av uniseriate, forgrenete filamenter. Forgreningen er uregelmessig og sidegrenen sitter ofte vinkelrett på hovedgrenen. I øverste ende av en celle dannes det ofte en kjertelcelle. Denne er uten farge og har en karakteristisk, trekantet form. Belegg: Preparat 8. Forekomst: Arten er vanlig i alle prøver på samtlige dyp. Fertile strukturer: Ikke funnet. 19
24 Kommentarer: Tetrasporofyttstadiet av B. hamifera har tidligere blitt beskrevet som en egen art under navnet Trailliella intricata. Trailliella-navnet har blitt hengende igjen og benyttes fortsatt for å beskrive tetrasporofytten hos B. hamifera. I mange individer jeg har sett på av denne arten har det vært få, i noen nesten ingen, kjertelceller. Jeg har ikke kommet over litteratur som forklarer dette fenomenet. Orden CERAMIALES Familie Ceramiaceae Aglaothamnion priceanum Maggs, Guiry & Rueness Beskrivelse: Fra den opprette hovedaksen utgår det et sideskudd fra hver celle, regelmessig avvekslende til den ene og den andre siden. De første to cellene i sideskuddet bærer gjerne en adaksial (vender opp og inn mot hovedaksen) forgrening, den tredje en abaksial (vender ned og ut fra hovedaksen) forgrening og deretter avvekslende forgrening. Sideskuddene ligger i ett plan slik at planten får et flatt uttrykk. Cellene er uninukleære. Belegg: Plansje 1, bilde e). Preparat 9. Forekomst: Kun ett individ er funnet. Det ble funnet i en prøve fra 15 meters dyp. Fertile strukturer: Ikke funnet. Kommentarer: Denne arten beskrives for første gang i en artikkel fra 1991 (Maggs et al., 1991). I følge denne artikkelen kan denne arten ligne A. bipinnatum i morfologi og generelt utseende. Artene kan blant annet skilles på at A. bipinnatum generelt ikke har forgrening på de første 1-2 cellene av sidegrenene, mens A. priceanum har forgrening fra første celle (Maggs & Hommersand, 1993). I tillegg kommer algenes respektive voksesteder som ytterligere sannsynliggjør at mitt funn dreier seg om A. priceanum. Denne vokser i følge Maggs og Hommersand (1993) på områder som er moderat til ekstremt eksponert. A. bipinnatum derimot trives best der det er veldig beskyttet til moderat eksponert. 20
. Preparat 10 og 11.")
25 Antithamnionella floccosa (O.F. Müller) Whittick Beskrivelse: Hovedaksen er avvekslende grenet, mens smågrenene er motsatt grenet. Forgreningen ligger i et plan, slik at plantene får en fjærform. Kjertelceller, som ellers er typisk for mange nærstående arter, er helt fraværende. Belegg: Plansje 1, bilde d). Preparat 10 og 11. Forekomst: Ett veldig lite individ funnet på 5 meter, ellers kun funnet i to prøver på 20 meter. Fertile strukturer: Ikke funnet. Callithamnion tetragonum (Withering) S.F. Gray Beskrivelse: Tallus består av en hovedakse med en sidegren fra hver celle. I yngre deler av planten er forgreningen avvekslende, senere sitter sidegrenene gjerne i spiral. Som hos andre arter i slekten har C. tetragonum flere kjerner i hver celle. Barkdannelse i proksimale deler av tallus. Tetraedrisk delte tetrasporangier som alltid sitter på oversiden av sidegrenene (adaksialt - nærmest hovedaksen). Belegg: Plansje 2, bilde a). Preparat 12, 13 og 14. Forekomst: En veldig vanlig art i mine prøver, den er funnet på alle dyp unntatt 25 meter. Fertile strukturer: Tetrasporangier funnet på de fleste individer. 100 µm Figur 13: Individ av Callithamnion tetragonum der cellekjernene er farget. Figuren viser flere cellekjerner i hver celle. Kommentarer: Denne arten er veldig vanlig i mine prøver. De fleste av individene er små, bare noen millimeter lange og den typiske formen med broddspisse sideskudd samlet i knipper er ennå ikke veldig tydelig. For å kunne bestemme disse individene til Callithamnion ble derfor cellekjerner farget slik at det ble tydelig at det fantes flere kjerner i hver celle, se figur 13. Dette for å unngå å forveksle disse små individene med arter av Aglaothamnion hvor hver celle kun har en kjerne. 21
26 Individer av denne arten satt gjerne ikke direkte på L. hyperborea, men epifyttisk på andre rødalgeepifytter. Slekten Ceramium Roth Slekten består av buskaktige alger. Store aksialceller er ved leddene omgitt av periaksialceller. Barkkledning varierer fra å være begrenset til leddene til å være heldekkende. Forgreningen er som regel gaffelgrenet. Antall periaksialceller, grad av barkkledning og antall ledd mellom forgrening er noen av de viktigste morfologiske karakterene ved artsbestemmelse. Ceramium-arter er morfologisk variable noe som har ført til det Maggs og Hommersand (1993) kaller taksonomisk kaos blant de artene uten pigger. Særlig har de artene der aksialcellene er helt dekket av bark blitt forvekslet under under navnet C. rubrum (Hudson) C. Agardh. Maggs et al. (2002) belyser artsinndelingen i dette C. rubrumkomplekset ved hjelp av molekylære undersøkelser. I følge denne artikkelen er antall segmenter mellom forgreningene et av de viktigste morfologiske karakteristika for å skille mellom artene i C. rubrum-komplekset. Ceramium shuttleworthianum (Kützing) Rabenhorst Beskrivelse: Denne Ceramium-arten er lett å kjenne igjen på den trekantete piggen som dannes i hvert barkbelte, se plansje 2, bilde b). Den har 4 periaksialceller. Forgrening skjer hvert 6-10 segment. Barklaget er begrenset til leddene. Belegg: Plansje 2, bilde b). Preparat 15. Forekomst: Funnet kun i tre prøver, to fra 10 og en fra 15 meters dyp. Fertile strukturer: Ikke funnet. Kommentarer: Arten vokser epilithisk, epizoisk eller epifyttisk i litoralsonen, aldri sublittoralt (Maggs & Hommersand, 1993). Rueness skriver at arten forekommer i nedre del av litoralsonen (Rueness, 1977). Mine funn av denne arten ble som nevnt gjort fra 10 og 15 meters dyp. Dette stemmer med andre ord ikke med beskrevne voksesteder for arten. Arten har et karakteristisk utseende, og det er ikke tvil om artsbestemmelsen. 22
27 Individet fra 15 meter var fargeløst og dødt og har sannsynligvis vokst et annet sted. Det kan stilles spørsmål ved hvorvidt ikke det samme gjelder de to individene på 10 meter også. Et av disse var løst og kan ha vokst et annet sted. Det andre satt fast, men i en geleaktig klump slik at det ikke var mulig å se hvorvidt det hadde spirt epifyttisk på dette dypet eller om det hadde løsnet fra et voksested høyere oppe, sunket ned, og satt seg fast i denne geleaktige klumpen. På bakgrunn av artens beskrevne voksested i litteraturen vil jeg anslå det sistnevnte som mest sannsynlig. Ceramium virgatum Roth Beskrivelse: Hos Maggs et al. (2002) finner man følgende beskrivelse av arten: Arten er veldig formvariabel. Antall segmenter mellom forgreninger varierer mer enn hos andre Ceramium-arter og er som regel 7-19 segmenter. Antall periaksialceller varierer også, populasjoner er funnet med 6-7, 7-8 eller 8 periaksialceller. I følge Rueness (Upublisertb) er dette kanskje den vanligste rødalgen i Norge. Belegg: Plansje 2, bilde c). Preparat 16. Forekomst: Kun funnet i to prøver, en fra 10 meters dyp og en fra 15 meters dyp. Fertile strukturer: Ikke funnet. Kommentarer: Hos C. virgatum skal antallet segmenter mellom forgreningene variere mer enn hos C. secundatum. På grunn av individenes størrelser (små), var antallet segmenter ikke et godt trekk å skille disse artene på. Viktigere har derfor snittene vært. De fleste av disse viser 7 periaksialceller, men noen viste 6 (se bilde av snitt innfelt på bilde c) plansje 2). Også C. secundatum kan ha 7 periaksialceller, men bare C. virgatum kan også ha 6 (Maggs et al., 2002). Pterothamnion plumula (J. Ellis) Nägeli Beskrivelse: Hovedakser er gaffelgrenet, mens smågrener er motsatt forgrenet. Smågrener av siste orden utløper kun fra oversiden. Alle grener har broddspisse endeceller. Det finnes som regel rikelig med kjertelceller. Disse sitter også alltid på oversiden av smågrenene. Belegg: Plansje 2, bilde d). Preparat 17 og
 skriver at arten vokser fra ekstremt lavvann og ned til 20 meters dyp.")
28 Forekomst: Arten er funnet i flere prøver fra 15 meter, 20 meter og 25 meters dyp, men bare med en til få eksemplarer i hver prøve. Fertile strukturer: Ikke funnet. Kommentarer: Maggs og Hommersand (1993) skriver at arten vokser fra ekstremt lavvann og ned til 20 meters dyp. Det kan det virke som om det er noe som gjør at den trives dypere på Skipsholmen ettersom jeg ikke har funnet den over 15 meters dyp samtidig som jeg har funnet den flere ganger på 25 meters dyp. Ptilota gunneri P.C. Silva, Maggs & L.M. Irvine Beskrivelse: Tallus blir 5-30 cm og er fjærgrenet i flere ordner. Fra hovedaksen vokser motsatt stilte fjærgrenete skudd. Hele tallus er dekket av småcellet bark. Både karposporer og tetrasporer sitter på smågrener (se figur 14). Belegg: Preparat 19 og 20. Herbarieark 1. Forekomst: Arten er vanlig på alle dyp og er til stede i samtlige prøver. I mange prøver er den også dominerende. Fertile strukturer: Karposporer og tetrasporangier funnet. Figur 14: Ptilota gunneri med tetrasporer. Spermothamnion repens (Dillwyn) Rosenvinge Beskrivelse: Opprette, uniseriate, grenete cellerekker vokser fra krypende filamenter. De krypende cellerekkene er festet til underlaget med encellede rhizoider. Cellene er lange, 4-10 ganger lenger enn brede. Forgreningen kan være både motsatt, avvekslende og ensidig. Fertile strukturer sitter på korte sidegrener. Spermatangier, gonimoblaster og tetrasporangier kan forekomme på samme individ. Belegg: Plansje 2, bilde e). Preparat 21 og 22. Forekomst: Funnet sporadisk i mange prøver fordelt på alle dyp med unntak av 25 meter. Fertile strukturer: Karposporer funnet, se plansje 2, bilde e). 24
29 Familie Delesseriaceae Cryptopleura ramosa (Hudson) L. Newton Beskrivelse: Tallus er bladformet og kan være enten krypende eller opprett. Jeg har kun funnet den krypende formen. Denne er festet til underlaget med mange små festerøtter som dannes på undersiden av bladet (se plansje 2, bilde f)). Nervene er tynne, gjerne bare en til få celler brede. Tetrasporangier dannes i sori i utvekster fra bladkanten. Bladet har gjerne et skimmer av blått. Belegg: Plansje 2, bilde f). Preparat 23. Forekomst: Arten har vært vanlig i samtlige prøver fra 10 meter, men kun funnet i en eneste prøve på andre dyp (en prøve fra 15 meter). Jeg har kun funnet den krypende formen av arten, og nesten utelukkende har disse vokst på hapter. Fertile strukturer: Tetrasporer funnet. Kommentarer: Arten ble for første gang funnet i Norge i 1962 (Baardseth, 1974). Den omtales fortsatt som sjelden i våre farvann (Rueness, Upublisert-b). Delesseria sanguinea (Hudson) J.V. Lamouroux Beskrivelse: Fra en festeskive vokser en stilk som går over i en tydelig midtribbe i bladdelen. Sidenerver er også tydelige. Bladet er ovalt lansettformet og bladranden er helt jevn, se figur 15. På høsten slites bladet mer eller mindre vekk slik at kun midtribben blir igjen. Nye blader utvikler seg fra midtribben. Herfra dannes også de reproduktive småbladene. Belegg: Herbarieark 2. Forekomst: Arten ble kun funnet i fire prøver, en fra 10 meter, to fra 15 meter og en fra 20 meter. Fertile strukturer: Ikke funnet. Figur 15: Jevn bladrand hos Delesseria sanguinea. 100 µm 25

30 Membranoptera alata (Hudson) Stackhouse Beskrivelse: Tallus er båndformet og har en kraftig midtribbe. Forgreningen er gaffelgrenet i flere ordner. Tetrasporangier sitter jevnt fordelt langs midten av bladet. Belegg: Preparat 24. Forekomst: M. alata er en av de vanligste artene i prøvene. Den er funnet på alle dyp og i samtlige prøver med unntak av en fra 25 meter. Fra 20 og 25 meter er den funnet bare sporadisk, høyere opp er den til dels dominerende. Fertile strukturer: Tetrasporer funnet. Nitophyllum punctatum (Stackhouse) Greville Beskrivelse: Blad av denne arten er bare ett cellelag tykt. Bladet mangler nerver og stilk, gir et mykt og skjørt inntrykk og rives også lett. Det er veldig formvariabelt, hele spekteret fra et nesten udelt blad til gaffelgrenete båndformer finnes. Belegg: Plansje 2, bilde g). Preparat 25. Herbarieark 3. Forekomst: Arten er funnet i prøver fra alle dyp med unntak av 25 meter. Fertile strukturer: Ikke funnet. Kommentarer: I følge Rueness (1977) er denne arten meget sjelden i Norge. Phycodrys rubens (Linnaeus) Batters Beskrivelse: Bladaktig tallus. Gjennom bladet løper en midtribbe og sidenerver. Bladene har tannet kant (se figur 16) og blir etter hvert flikete. Tetrasporangier er samlet i utvekster fra bladkanten, eller spredt over småblad. Cystokarper dannes spredt i bladkanten eller enkeltvis i småblader. Belegg: Preparat 26. Herbarieark 4. Forekomst: Arten har vært en av de aller vanligste i min undersøkelse. Den er funnet i samtlige prøver og har på flere dyp også vært delvis dominerende (15 og 20 meter). Fertile strukturer: Både cystokarper og tetrasporangier funnet. 100 µm Figur 16: Bladrand hos et ungt blad av Phycodrys rubens. I motsetning til bladranden hos Delesseria sanguinea er denne ikke jevn. 26
31 Kommentarer: Figur 16 viser bladkanten av et ungt blad på < 1 cm. Allerede vises de taggete kantene. Sammenlign bladranden på bildet med den jevne bladranden til Delesseria sanguinea. Familie Rhodomelaceae Brongniartella byssoides (Goodenough & Woodward) F. Schmitz Beskrivelse: Hovedgrenene har en polysifon oppbygning med 7 periaksialceller. På grenene sitter det gaffelgrenete, uniseriate skudd i spiral. Dette gjør algen lett gjenkjennelig. Belegg: Plansje 3, bilde a). Preparat 27. Forekomst: Funnet i mange prøver og på alle dyp unntatt 10 meter, men aldri mer enn ett eller få individer i hver prøve. Fertile strukturer: Ikke funnet. Kommentarer: Maggs og Hommersand (1993) skriver at denne algens voksesteder varierer fra ekstremt bølgebeskyttet til ekstremt bølgeeksponert. Rueness (1977), derimot, skriver at algen foretrekker noe beskyttede steder. Dette kan være forklaringen på at jeg ikke har funnet mange individer av denne arten til tross for at den er til stede på de fleste dyp. Det kan også være en forklaring på at jeg har funnet den oftest i prøvene fra 20 og 25 meter der det er mindre eksponert. Jeg har dog ikke nok prøver til at man kan utelukke at dette kan være tilfeldig. Odonthalia dentata (Linnaeus) Lyngbye Beskrivelse: Arten har et båndformet tallus med en utydelig midtribbe. Forgreningen er uregelmessig, og randen er grovt sagtannet. Belegg: Plansje 3, bilde d). Preparat 28. Herbarieark 5. Forekomst: Det er kun funnet ett individ av arten, i en prøve fra 10 meter. Fertile strukturer: Ikke funnet. 27

32 Slekten Polysiphonia Greville Tallus består av polysifone skudd med 4-24 periaksialceller. I mine prøver har jeg kun funnet arter med 4 periaksialceller. Tetrasporangier sitter på rekke i endegrenene, cystokarper er runde eller urneformede. Spermatangiene utvikles fra basis av trichoblaster. Gametofyttene hos Polysiphonia kan være enten monøse eller diøse, men begge artene beskrevet her er diøse. Polysiphonia fibrillosa (Dillwyn) Sprengel Beskrivelse: Denne Polysiphonia-arten har 4 periaksialceller. Barkkledning i eldre deler av planten. Trichoblaster er gjerne tallrike. Belegg: Plansje 3, bilde b). Preparat 29 og 30. Forekomst: Arten er funnet i samtlige prøver på 10 meter og 15 meter, og ett enkelt individ er funnet på 20 meter. Fertile strukturer: Tetrasporofytter (som på figur 17 se tetraspore) og både hannlige og hunnlige gametofytter med enten spermatangier og cystokarper er funnet i flere prøver. 100 µm Figur 17: Polysiphonia fibrillosa med trichoblaster (stiplet pil) og tetraspore (heltrukken pil). Polysiphonia stricta (Dillwyn) Greville Beskrivelse: De 4 periaksialcellene er aldri barkkledde. Trichoblaster mangler også. Cystokarpene er urneformete og sitter på stilk. Fargen er mørkerød. Belegg: Preparat 31 og 32. Herbarieark 6. Forekomst: Arten er av de vanligste i samtlige av mine prøver. Fertile strukturer: Både hannlige gametofytter med spermatangier og hunnlige gametofytter med cystokarper er funnet i prøvene. Tetrasporer ikke funnet. Kommentarer: Algen er i følge Rueness (1977) kanskje den vanligste av våre Polysiphonia-arter. Dette stemmer som nevnt for det materialet jeg har gjennomgått. 28
33 Pterosiphonia parasitica (Hudson) Falkenberg Beskrivelse: Skuddene hos denne arten er avvekslende fjærgrenet. De består av segmenter med 8-10 periaksialceller (i følge Rueness (1977), 7-8 periaksialceller i følge Maggs og Hommersand (1993)). Bark fraværende. Belegg: Plansje 3, bilde c). Preparat 33 og 34. Forekomst: Arten er funnet i nesten alle prøver fra 10, 15 og 20 meter, men er helt fraværende fra 25 meter. Kun enkeltfunn er gjort fra 5 meter. Arten er i hovedsak funnet på hapter. Fertile strukturer: Cystokarper funnet. Kommentarer: I følge Rueness (1977) regnes denne arten for å være sjelden hos oss. Rhodomela confervoides (Hudson) P.C. Silva Beskrivelse: Hovedaksene er rikt forgrenet. I tverrsnitt er tallus rundt med en sentralcelle i midten. Denne er omgitt av 6-7 periaksialceller som igjen er omgitt av bark, se innfelt bilde plansje 3, bilde g). Dette skiller arten fra Cystoclonium purpureum som har filamentøs medulla. Arten kan også forveksles med R. lycopodioides, men denne er mye tettere besatt med korte, stive smågrener. Belegg: Plansje 3, bilde g). Preparat 35 og 36. Herbarieark 7. Forekomst: Arten er kun funnet i noen få prøver, og kun fra 10 og 15 meter. Fertile strukturer: Ikke funnet. Kommentarer: I følge Maggs og Hommersand (1993) har tre arter i slekten Rhodomela blitt rapportert fra Nord-Vest Europa, R. confervoides, R. lycopodioides og R. virgata. Ettersom arten er formvariabel finnes ulike oppfatninger om hvorvidt disse er en eller flere 3 gode arter. Dette diskuteres også hos Maggs og Hommersand (1993) som konkluderer med at R. virgata bør regnes som samme art som R. confervoides, mens R. lycopodioides bør regnes som en egen, god art. Algaebase følger denne artsinndelingen for Rhodomela, og dermed gjør også jeg det i denne oppgaven. 29

34 Orden COLACONEMATALES Familie Colaconemataceae Slekten Colaconema Batters Tallus hos alle arter i slekten utgjøres av monosifone tråder, grenet eller ugrenet. Aseksuell reproduksjon skjer hovedsakelig med monosporangier. Kloroplastene, en eller mange, er parietale. Disse kan forekomme i forskjellige former, men er aldri stjerneformete (Harper & Saunders, 2002). Beskrivelsene under er i hovedsak hentet fra Rosenvinge (1909). Colaconema daviesii (Dillwyn) Stegenga Beskrivelse: I følge Rosenvinge (1909) er dette en lett gjenkjennelig art. Sporene vokser i typiske knipper. Knippene dannes ved at smågrener deler seg opp til 3 ganger. Hvert sideskudd sitter på innsiden av det forrige. Cellene er generelt kortere enn hos mange andre nærstående arter. Hver celle inneholder en velutviklet pyrenoide som kan bli så stor at den dekker hele cellens diameter og når kloroplasten på den andre siden av cellen (se figur 18, stiplet pil). Belegg: Plansje 3, bilde h). Preparat 37 og 38. Forekomst: Arten er funnet på alle dyp med unntak av 25 meter. Den er ikke funnet mange ganger på hvert dyp. Fertile strukturer: Monosporer funnet, (se figur 18, heltrukken pil). Kommentarer: Dette er, som flere av de rød trådformete artene, en art man må lete etter. Det kan da være litt tilfeldig hvorvidt man finner den i en prøve eller ikke. Det er derfor en fare at flere av disse artene er underdokumentert, både i min oppgave og i andres. 25 µm Figur 18: Colaconema daviesii med monospore (heltrukken pil) og pyrenoide (stiplet pil). 30
35 Colaconema membranaceum (Magnus) Woelkerling Beskrivelse: De krypende filamentene vokser endozoisk i flere hydroider. Disse filamentene kan noen ganger vokse så tett at det ser ut som en skive, andre ganger vokser de løst. Hvilken form individet vokser på avhenger av hydroidearten den vokser i (Rosenvinge, 1909 s ). Cellene har flere båndformete kloroplaster. Sporangiene sitter endestilt på opprette filamenter. Belegg: Plansje 3, bilde e) og f). Preparat 39 og 40. Forekomst: Arten er funnet på hydroider fra alle dyp. Den var mindre vanlig på 25 meter enn høyere opp, noe som kan skyldes at få hydroider var til stede på dette dypet. Fertile strukturer: Korsdelte tetrasporangier funnet. Kommentarer: Denne arten er i motsetning til de andre røde trådformede (se kommentar under forrige art) lett å få øye på da den gir hydroidene en rosa farge. Orden CORALLINALES Familie Corallinaceae Corallina officinalis Linnaeus Beskrivelse: En forkalket, oppreist, leddet, motsatt fjærgrenet alge. Den er vanlig så vel i fjærepytter som i sublittoralsonen. Belegg: Plansje 4, bilde a). Forekomst: Arten er funnet på 5, 15 og 20 meter. Den er kun funnet i noen prøver, men når den først har vært til stede har det vært flere til mange individer. Funnet hovedsakelig på hapter. Fertile strukturer: Ikke funnet. Lithophyllum crouaniorum Foslie Beskrivelse: Denne arten kan danne relativt tykke skorper. Cellene har en karakteristisk diamantfasong og er horisontalt plassert på linje med hverandre. De henger sammen med sekundære poreforbindelser, se eksempler piler figur 19 a). Tetrasporangie-konseptakler er globulære i form. Gamle konseptakler begraves ofte etter hvert i tallus, se bilde d) plansje 4. 31
 Cellerekker hos L.")
. Titanoderma pustulatum (J.V.")

36 Belegg: Plansje 4, bilde d). Preparat 41. Forekomst: De avbildete individene er fra 5 og 25 meters dyp. Fertile strukturer: Konseptakler med tverrdelte tetrasporer funnet, se figur 19 b). Kommentarer: Denne arten er beskrevet som L. crouanii Foslie, men stavemåten er nå endret til L. crouaniorum Foslie i henhold til korrekt latinsk gramatikk. Figur 19: a) Cellerekker hos L. crouaniorum med sekundære poreforbindelser (eksempler merket med piler). b) Konseptakel hos L. crouaniorum med tverrdelt tetraspore. a) 25 µm b) Både denne og den påfølgende arten, T. pustulatum, er tidligere funnet å være vanlige kalkalger på stipes av L. hyperborea (Fredriksen et al., 1995). Titanoderma pustulatum (J.V. Lamouroux) Nägeli Beskrivelse: Cellene i den basale cellerekken er høye og smale og litt skråstilte (palisadeceller). Arten har konvekse konseptakler med kun 2-3 cellerekker over Figur 20: Konseptakel hos Titanoderma pustulatum. konseptaklgulvet, se figur 20. Sporangiene er som regel bisporangier. Belegg: Plansje 4, bilde d). Preparat 42 og 43. Forekomst: Individet det er tatt bilder av er funnet på 10 meters dyp. Jeg har også registrert den fra 20 meters dyp. Fertile strukturer: Konseptakler med bisporer funnet, se bilde d), plansje 4. Kommentarer: Det er påvist stor variasjon innen arten. I del 2B av Seaweeds of the British Isles har derfor Irvine og Chamberlain (1994) valgt å følge en systematikk der man deler arten T. pustulatum inn i fire varieteter, var. canellatum, var. confine, var. macrocrocarpum og var. pustulatum. Dette har man ikke valgt å gjøre i Algaebase. Ettersom jeg følger Algaebase i denne oppgaven, har jeg ikke gått videre for å finne ut 32

37 hvilke(n) varietet(er) jeg har i mine prøver. Muligens ville jeg heller ikke hatt nok informasjon til å kunne bestemme individene til varietet. Familie Hapalidiaceae Melobesia membranacea (Esper) J.V. Lamouroux Beskrivelse: Dette er en veldig vanlig, epifyttisk, forkalket skorpe. Sporangiekonseptaklene har en konkav, mørk, mangeporet plate, noe som gir dem det lett gjenkjennelige mørke midtpartiet. Belegg: Plansje 4, bilde b). Forekomst: Det er funnet flere individer i samtlige prøver med unntak av en prøve på 25 meter. Arten er hovedsakelig funnet på hapter, men også i noen prøver på epifyttene. Den har for eksempel vært vanlig på stilker av Ptilota gunneri og Phycodrys rubens. Fertile strukturer: Mangeporet sporangiekonseptakler med tetrasporer funnet. Kommentarer: Jeg har i flere prøver funnet spiringsskive av en kalkalge, se figur 21. Den eneste muligheten jeg har hatt for å bestemme disse har vært å sammenligne med tegninger av spiringsskiver i litteraturen. Mine funn stemmer veldig godt med tegningen av spiringsskiven av M. membranacea i Irvine og Chamberlain (1994 s. 198), og det er ikke usannsynlig at det er denne arten jeg har funnet spiringsstadiet av. Orden GIGARTINALES Familie Kallymeniaceae Slekten Callophyllis Kützing 50 µm Figur 21: Spiringsskive av kalkalge, muligens Melobesia membranacea. Callophyllis cristata (C. Agardh) Kützing Beskrivelse: Tallus er oppreist, bladaktig og blir 1-5 cm høy. Uregelmessige, gjentatte oppdelinger i stadig smalere fliker gjør at den kan få en viftelignende form. Cystokarper utvikles langs bladranden, se figur
 Kützing Beskrivelse: Fra en liten festeskive vokser en kort stilk, vanligvis bare 2 mm.")
38 Belegg: Herbarieark 8. Forekomst: Flere individer av arten er funnet i de aller fleste prøver og på alle dyp. Den er hovedsakelig funnet på hapter. Fertile strukturer: Cystokarper funnet, se figur µm Figur 22: Callophyllis cristata med cystokarper i bladranden. Callophyllis laciniata (Hudson) Kützing Beskrivelse: Fra en liten festeskive vokser en kort stilk, vanligvis bare 2 mm. Deretter blir tallus bladaktig og blir 3-10 cm høy. Bladet har gjerne vifteform og er gaffelgrenet eller uregelmessig oppdelt i fliker. Belegg: Preparat 44. Herbarieark 9. Forekomst: Denne arten er funnet i mange prøver og på alle dyp, dog kun et enkelt individ på 5 meters dyp. Den er funnet både på stipes og hapter, men de største individene har vokst på hapter. Fertile strukturer: Ikke funnet. Familie Phyllophoraceae Coccotylus truncatus (Pallas) M.J. Wynne & J.N. Heine Beskrivelse: Et lite, skiveformet festeorgan fester tallus til substratet. Tallus består av en rund stilkdel som går gradvis over i et flatt bladavsnitt. Ser man på bladet i snitt vil man kunne se at medulla består av celler opptil 65 µm i diameter. Cellestørrelsen avtar utover til cortex der cellene er 2-5 µm i diameter. Flere kilder oppgir arten som meget formvariabel. Belegg: Plansje 4, bilde h). Forekomst: Kun funnet i en prøve på 5 meters dyp. En utvokst plante og noen få juvenile ble funnet. Alle satt på hapter. Fertile strukturer: Ikke funnet. 34
39 Kommentarer: I følge Rueness (1998) vokser denne arten vanligst på beskyttete steder. I dette tilfellet vokste planten ut fra en dyp forsenkning i hapteren og satt på den måten mer beskyttet enn lokaliteten prøvene er hentet fra skulle tilsi. Det utvokste individet var sterkt bleket og overgrodd med bryozooer. Eksemplaret var derfor ikke lett gjenkjennelig. For å forsikre meg om at jeg ikke forvekslet individet med f.eks Palmaria palmata som også fantes i prøven skar jeg snitt av begge for sammenligning. Dermed fikk jeg også målt cellene i medulla. Snittet og cellenes mål kombinert med at individet har en flere centimeter lang stilk, gjorde at jeg bestemte dette individet til C. truncata. Orden PALMARIALES Familie Palmariaceae Palmaria palmata (Linnaeus) Kuntze Beskrivelse: Et bladaktig tallus utgår fra en liten hefteskive. Bladet deles i brede fliker og forgreningsmønsteret er gaffelgrenet eller uregelmessig. Prolifikasjoner fra bladranden er vanlig. Belegg: Herbarieark 10. Forekomst: Arten er funnet i samtlige prøver på 5 meters dyp. I tillegg er den funnet i to prøver på 10 meter og en prøve på 20 meter. Fertile strukturer: Ikke funnet. Kommentarer: Individer av arten kan bli store, opp til 0,5 meter lange. Allikevel var også individene av også denne arten små i mine prøver, og kun i en prøve (plante 4 fra 5 meters dyp) fant jeg planter over 5 cm. Her var det til gjengjeld et individ på over 20 cm. Familie Rhodophycemataceae Meiodiscus spetsbergensis (Kjellman) Saunders & McLachlan Beskrivelse: Arten har en karakteristisk basal celleskive. De krypende filamentene ligger tett (fusjon mellom celler er ikke uvanlig) og danner en skive som sprer seg i vifteform. 35
40 Kloroplastene varierer fra å være skiveformete til mer eller mindre båndformete. Sporangiene sitter gjerne enkeltvis og endestilt på de opprette skuddene. Belegg: Plansje 4, bilde e). Preparat 45 og 46. Forekomst: Arten er funnet på alle dyp. Dette til tross for at den i Dixon og Irvine (1977) er beskrevet til kun å vokse ned til 10 meter. Den er funnet både epifyttisk og i flere tilfeller epizoisk på hydroider. Fertile strukturer: Tetrasporangier funnet. Kommentarer: Denne arten er vanskelig å artsbestemme med mindre man finner celleskiven eller tetrasporangier. Flere av de individene som er identifisert til denne arten har vokst på hydroider. I disse tilfellene har jeg kunnet løsne hele hydroiden slik at celleskiven har kommet uskadet, og dermed tydelig, frem. Der individer har vokst epifyttisk og jeg har forsøkt å løsne filamenter fra underlaget har jeg stort sett ikke fått med nok av celleskiven til sikker identifikasjon og vært avhengig av at individet har vært fertilt for å kunne identifisere arten. Denne arten er derfor sannsynligvis vanligere i mine prøver enn min dokumentasjon kan tyde på. Orden PLOCAMIALES Familie Plocamiaceae Plocamium cartilagineum (Linnaeus) P.S. Dixon Beskrivelse: Tallus er flatt. Hovedaksen er avvekslende grenet. Sympodialt forgrenet, dvs. smågrener i flere ordner er ensidig stillet i serier på 2-5. Disse er alle avrundet mot aksen slik at siste ordens grener er formet som en kam. Belegg: Plansje 4, bilde f). Preparat 47. Herbarieark 11. Forekomst: Arten er kun funnet på 10 meter og 15 meter, men på disse dypene har den til gjengjeld vært vanlig. Fertile strukturer: Ikke funnet. 36
41 Orden RHODYMENIALES Familie Lomentariaceae Slekten Lomentaria Lyngbye Lomentaria clavellosa (Turner) Gaillon Beskrivelse: Tallus er hult og festet til underlaget med en liten skive. Forgreningen er uregelmessig og grenene avsmalner mot spiss og basis. Belegg: Preparat 48. Herbarieark 12. Forekomst: Arten er funnet i samtlige prøver fra og med 15 meter og nedover. Over dette har den vært sjeldnere og er bare sporadisk funnet. Den er funnet både på hapter og stipes. Fertile strukturer: Tetrasporer funnet. Kommentarer: Individer av arten kan bli inntil 40 cm høye, men er hovedsakelig funnet som kimplanter i mine prøver. Lomentaria orcadensis (Harvey) F.S. Collins Beskrivelse: Tallus er hult og uregelmessig grenet som L. clavellosa, men grenene er gjerne mer avsmalnende både mot spiss og mot basis. 2-5 cm høyde er fullt utvokst størrelse for arten. Individer av arten har i motsetning til L. clavellosa ofte horisontale utløpere fra basis. Disse utløperene er tydelig på bildet g) plansje 4. Belegg: Plansje 4, bilde g). Preparat 49. Herbarieark 13. Forekomst: Arten er kun funnet i noen prøver, men disse fordeler seg på alle dybder unntatt på 25 meters dyp. Vanligst på hapter. Fertile strukturer: Tetrasporer funnet. 37

42 Phylum CHLOROPHYTA Klasse BRYOPSIDOPHYCEAE Orden BRYOPSIDALES Familie Derbesiaceae Derbesia marina (Lyngbye) Solier Beskrivelse: Sporofytten danner 1-3 cm høye tuster av uregelmessig grenete tråder. Algen er sifonal, den har ingen ekte tverrvegger. I et filament er det derfor mange cellekjerner. Det er også mange skiveformete kloroplaster. Zoosporer dannes i sidestilte, kortstilkete sporangier. Belegg: Plansje 5, bilde a). Forekomst: Arten er funnet i mange prøver og på alle dyp. I flere av prøvene har den også vært veldig vanlig. I de fleste tilfeller har den vokst epizoisk på bryozooer på tarestilken. Fertile strukturer: Sporangier funnet i flere prøver, se figur 23. Figur 23: Derbesia marina. Sporangium med zoosporer. Familie Ostreobiaceae Ostreobium quekettii Bornet & Flahault Beskrivelse: Arten vokser i kalken til kalkdannende dyr. Filamenter er kun 2-5 µm i diameter uten tverrvegger. Filamentene kan være rette og sparsomt forgrenet, eller knudrete og rikt forgrenet. Oppsvellede partier kan forekomme. Belegg: Plansje 5, bilde b). Preparat 50. Forekomst: Denne arten er funnet i nesten alle prøver gjennomgått etter at jeg først ble oppmerksom på den. Den er funnet på alle dyp i kalken på kalkrørsmark og i bryozooer. Fertile strukturer: Ikke funnet. 38

43 Kommentarer: På grunn av artens størrelse, er det mange detaljer som er vanskelige å se i lysmikroskop. Viktig for artsbestemmelse av denne arten har derfor i tillegg til voksemåte vært å sammenligne mine funn med illustrasjoner og publiserte bilder av arten. For eksempel har Pantazidou (2006) gode SEM-fotografier, og Feussner et al. (2004) inkluderer bilder av O. quekettii sett i lysmikroskop i sin artikkel. Algen gir kalken den vokser i et grønnlig preg. Fikk jeg mistanke om at arten var til stede, fjernet jeg kalken med saltsyre. Uten unntak fant jeg da O. quekettii. I noen av prøvene jeg avkalket kunne man også se lange, rette filamenter. Disse var marginalt tykkere og mer regelmessige enn de typiske uregelmessige Ostreobiumfilamentene. Jeg fikk derfor mistanke om at det vokste en annen art sammen med O. quekettii ettersom de fleste kilder beskriver nettopp uregelmessigheten som et kjennetegn ved arten. Newton (1931), derimot, viser en illustrasjon av arten der begge observerte typer filamenter er tegnet. Jeg har derfor konkludert med at de observerte filamentene i mine prøver kun er O. quekettii. Klasse CHLOROPHYCEAE Orden CHAETOPHORALES Familie Chaetophoraceae Entocladia viridis Reinke Beskrivelse: Tallus består av krypende filamenter som vokser endofyttisk i celleveggen hos et bredt spekter av alger. Cellene er sylindriske til uregelmessige og har en parietal kloroplast, som regel med en pyrenoide (noen celler kan ha flere), se for eksempel pil figur 24. Belegg: Plansje 5, bilde c). Preparat 51. Forekomst: Arten ble kun funnet i en prøve, denne fra 15 meter. Der vokste den endofyttisk i Phycodrys rubens. Fertile strukturer: Ikke funnet. 250 µm Figur 24: Entocladia viridis. Pilen peker på et eksempel på pyrenoide. Pyrenoider er også tydelige i de andre cellene. 39

44 Phylum OCHROPHYTA Klasse PHAEOPHYCEAE Orden CUTLERIALES Familie Cutleriaceae Cutleria multifida (Turner) Greville, Aglaozonia-stadiet Beskrivelse: Aglaozonia-stadiet er artens diploide sporofyttstadium. Det vokser som en skive, og er løst festet til underlaget med rhizoider, se figur 25. Overflatecellene er rektangulære, og ligger i langsgående rekker. Typiske avrundete fliker langs kanten. Belegg: Plansje 5, bilde d). Preparat 52 og 53. Forekomst: Arten er funnet flere ganger i nesten samtlige prøver fra 10 meter og 15 meter, men ikke et eneste individ er funnet fra andre dyp. Arten er utelukkende funnet på hapterene. Fertile strukturer: Ikke funnet. Figur 25: Undersiden av Cutleria multifida med hefterøtter. 50 µm Orden DESMARESTIALES Familie Desmarestiaceae Desmarestia viridis (O.F. Müller) J.V. Lamouroux Beskrivelse: Tallus er trådformet og rikt motsatt grenet. En stor sentralcelle omgis av et tykt barklag (når planten er utvokst). I tverrsnitt er grenene sirkelrunde. Belegg: Plansje 5, bilde e). Preparat 54. Forekomst: Arten har ikke vært vanlig på noe dyp, men er funnet i flere prøver på 15 meter. I tillegg har den forekommet i en prøve på 5 meter og en på 20 meter. Fertile strukturer: Ikke funnet. Kommentarer: Kun små kimplanter av arten er funnet. 40
45 Orden ECTOCARPALES Familie Ectocarpaceae Ectocarpus sp. Beskrivelse: Tallus består av enradete, forgrenete cellerekker. Kloroplastene er båndformete og har flere pyrenoider. Flerradete plurilokulære sporangier som som regel sitter sidestilt på en kort stilk. Belegg: Preparat 55. Forekomst: Slekten er funnet flere ganger i samtlige prøver fra 20 meters dyp. Den er ikke funnet på 15 meters dyp, på de resterende dypene er den funnet sporadisk. Fertile strukturer: Plurilokulære sporangier er funnet på mange av individene. Kommentarer: Individer av denne slekten er ikke bestemt til art.. I Norge skiller man pr. i dag mellom to arter, E. siliculosus og E. fasciculatus (Rueness, Upublisert-a). Et viktig skilletegn mellom disse er forgreningen. Individene jeg har funnet har vært for små til å kunne si noe om forgrening. Videre kan man skille artene på at de plurilokulære sporangiene til E. siliculosus ofte ender i et pseudohår. Jeg har ikke funnet slike hår. Dette kan tilsi at mine funn dreier seg om E. fasciculatus. Slike hår er dog ikke alltid til stede. Dessuten har ikke alle individer jeg har funnet hatt sporangier i det hele tatt. I tillegg kommer at formvariasjonen er stor innenfor denne gruppen (Rueness, 1977). Jeg har derfor valgt å kun bestemme disse algene til slekt. Familie Acinetosporaceae Hincksia hincksiae (Harvey) P.C. Silva Beskrivelse: Som hos Ectocarpus er cellerekkene uniseriate og rikt grenete. Derimot har Hincksia skiveformete kloroplaster. De plurilokulære sporangiene er uten stilk og sitter på innsiden av grenene så tett at de berører hverandre. Belegg: Plansje 6, bilde a). Preparat 56. Forekomst: Er kun funnet i 3 prøver, en prøve på henholdsvis 5, 10 og 15 meter. Fertile strukturer: Rikt med plurilokulære sporangier funnet. 41

46 Familie Chordariaceae Protectocarpus speciosus (Børgesen) Kornmann Beskrivelse: Med det blotte øyet ses arten kun som små, brunfargede flekker mot underlaget. Ut fra en basal skive vokser opprette, uniseriate filamenter som kan bli opp til 1 mm. Kloroplasten er uregelmessig skiveformet. De plurilokulære sporangiene er flerradete, og kan sitte basalt på skiven, sidestilt eller endestilt på filamentene. Unilokulære sporangier sitter gjerne sidestilt og finnes ofte på samme individ som bærer plurilokulære sporangier (som på figur 26). Belegg: Plansje 6, bilde d) og e). Preparat 57 og 58. Forekomst: Funnet i noen få prøver på alle dyp unntatt 25 meter. Hovedsakelig funnet på hapterene, men også funnet på Palmaria palmata. Fertile strukturer: Både plurilokulære og unilokulære sporangier funnet, se figur 26. Kommentarer: Funnene kan også ligne Myrionema, men denne 25 µm slekten har som hovedregel uniseriate plurilokulære sporangier, sjelden biseriate (Fletcher, 1987). De plurilokulære sporangiene i mitt funn er ofte multiseriate. Figur 26: Protectocarpus speciosus med et unilokulært og et plurilokulært sporangium på samme filament. Orden LAMINARIALES Familie Laminariaceae Laminaria sp. Beskrivelse: Tallus er differensiert i hapter, stipes og lamina. Vekstsonen sitter i overgangen mellom stipes og lamina, og derfra vokser et nytt blad frem hvert år. Unilokulære sporangier sitter i sori som kan ses som mørke flekker på lamina. Belegg: Preparat 59 Forekomst: Juvenile individer som ikke har latt seg bestemme til art har vært relativt vanlig. De er funnet i minst to prøver på alle dyp. Hovedsakelig er de funnet på hapter. Fertile strukturer: Ikke funnet. 42
47 Orden SPHACELARIALES Familie Sphacelariaceae Slekten Sphacelaria Lyngbye Alle arter i denne slekten har en typisk vekstmåte. En sylindrisk, gjerne mørkere farget toppcelle gir opphav til segmenter. Hvert segment deles først en gang på tvers (til to sekundære segmenter) og deretter på langs (hvor mange ganger varierer mellom artene). Denne segmenteringen gjør at Sphacelaria-arter er lett gjenkjennelige. Kloroplastene er mange og skiveformete. Ved gjennomgang av denne slekten har jeg benyttet Prud homme van Reine (1982). Sphacelaria caespitula Lyngbye Beskrivelse: Arten danner 5-10 mm høye tuster av oppreiste skudd som er sparsomt forgrenet. Den vokser epifyttisk, helst på stipes av Laminaria hyperborea. Skuddene er µm brede. Sekundære segmenter er ofte delt med tverrvegger. Propagulae er ukjent. Pericyster forekommer. Belegg: Plansje 6, bilde b). Preparat 60. Forekomst: Funnet på alle dyp, dog kun med ett individ fra 5 meter. Arten var vanligst på 20 og 25 meters dyp. Fertile strukturer: Ingen funnet Sphacelaria plumosa Lyngbye Beskrivelse: Hovedakser uregelmessig forgrenet, sideskudd fjærgrenet. Hovedfilamenter får etter hvert rhizoidal bark. Disse er µm brede før bark dannes og opp til 700 µm brede etter barkdannelse. Sideskudd er µm brede. På hovedaksen kan man se langsgående vegger. Sekundære tverrdelinger er vanlig. Propagulae er ukjent. S. plumosa er en vanlig art som i tillegg til å vokse epifyttisk på Laminaria hyperborea vokser på skjell og stein. Belegg: Plansje 6, bilde c). Preparat
48 Forekomst: Denne arten er kun funnet i to prøver, begge fra på 25 meter. I disse prøvene var arten til gjengjeld vanlig. Fertile strukturer: Ingen funnet Sphacelaria spp. I flere prøver, og på samtlige dyp, har jeg Sphacelaria-arter som ikke er en av de overnevnte. Disse har vært umulige å bestemme til art. Sannsynligvis er en stor andel av disse S. cirrosa. Jeg kan heller ikke utelukke at noen av dem er S. rigidula. 44
 10 µm e) 100 µm")

 Erythrocladia")

 Acrochaetium minimum med")
 Antithamnionella")
 Aglaothamnion priceanum.")
49 PLANSJE 1 a) b) 25 µm c) 100 µm d) 10 µm e) 100 µm f) 100 µm a) Erythrocladia irregularis på Spermothamnion repens. b) Erythrocladia irregularis og Acrochaetium minimum på Polysiphonia fibrillosa. c) Acrochaetium minimum med monosporangium (pil). d) Antithamnionella floccosa. e) Aglaothamnion priceanum. f) Porphyropsis coccinea sett i lupe med mikroskopbilde av celler innfelt. 50 µm 45

 100 µm g) a)")

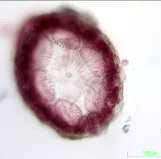







50 PLANSJE 2 a) b) 1000 µm 100 µm c) d) 100 µm e) 1000 µm 100 µm f) 100 µm g) a) Callithamnion tetragonum. b) Ceramium shuttleworthianum. c) Ceramium virgatum med snitt som viser 6 periaksialceller innfelt. d) Pterothamnion plumula. e) Spermothamnion repens med gonimoblaster (piler). f) Underside av Cryptopleura ramosa som viser pseudonerver og kroklignende hapter. 46
 d) e) 100 µm")
 Brongniartella")
 Hanlig gametofytt av")
 Pterosiphonia")
 Odonthalia dentata.")


 Rhodomela")
51 PLANSJE 3 a) 100 µm b) 100 µm c) d) e) 100 µm f) 50 µm g) 100 µm h) 100 µm a) Brongniartella byssoides. b) Hanlig gametofytt av Polysiphonia fibrillosa. c) Pterosiphonia parasitica. d) Odonthalia dentata. e) Colaconema membranaceum i hydroide. f) Colaconema membranaceum med tetrasporer. g) Rhodomela confervoides i lupe med snitt som viser 6 periaksialceller innfelt. h) Colaconema daviesii. 47
 100 µm c) 100")
 100 µm a)")



")




 Meiodiscus")
52 PLANSJE 4 a) b) 100 µm d) 100 µm c) 100 µm 100 µm e) f) g) h) 100 µm a) Corallina officinalis, bilde av avkalkede ledd innfelt. b) Melobesia membranacea med tetrasporer, habitus sett i lupe innfelt. c) Titanoderma pustulatum, konseptakel med bisporer, algen sett ovenfra i lupe innfelt. d) Lithophyllum crouaniorum med globulære konseptakler. e) Meiodiscus spetsbergensis. f) Plocamium cartilagineum. g) Lomentaria orcadensis. h) Coccotylus truncatus. 48
 e) a)")


 Entocladia viridis.")


53 PLANSJE 5 a) 100 µm b) c) 10 µm 50 µm d) e) a) Derbesia marina med sporangium, innfelt bilde av arten voksende på bryozooer sett i lupe. b) Ostreobium quekettii. c) Entocladia viridis. d) Cutleria multifida Aglaozonia-stadiet. Innfelt den skorpefomete algen som vokser over hapter. e) Desmarestia viridis. 100 µm 49
 100 µm a)")

 Sphacelaria plumosa.")


54 PLANSJE 6 a) b) c) 100 µm d) 100 µm 100 µm e) 100 µm a) Hincksia hincksiae med plurilokulære sporangier. b) Sphacelaria caespitula. c) Sphacelaria plumosa. d) Protectocarpus speciosus med plurilokulære sporangier både fra basalskiven og opprette grener. e) Bit av den basale skiven til Protectocarpus speciosus. 50 µm 50
LIVET I FJÆRA. Illustrasjoner: Stein Mortensen. Tegnforklaring: Rød triangel indikerer en introdusert art.
 ALGER LIVET I FJÆRA Illustrasjoner: Stein Mortensen Tegnforklaring: Rød triangel indikerer en introdusert art. GRISETANG Ascophyllum nodosum Stasjonsnummer Blæretang Du kan finne alderen ved å telle antall
ALGER LIVET I FJÆRA Illustrasjoner: Stein Mortensen Tegnforklaring: Rød triangel indikerer en introdusert art. GRISETANG Ascophyllum nodosum Stasjonsnummer Blæretang Du kan finne alderen ved å telle antall
Havbrukstjeneten AS 7260 Sistranda
 Havbrukstjeneten AS 7260 Sistranda Telefon: 72 44 93 77 Felefaks: 72 44 97 61 Internett: www.havbrukstjenesten.no E-post: arild@havbrukstjenesten.no Rapport nr: StrFjo0913 Gradering: Åpen Strandsone Rapport
Havbrukstjeneten AS 7260 Sistranda Telefon: 72 44 93 77 Felefaks: 72 44 97 61 Internett: www.havbrukstjenesten.no E-post: arild@havbrukstjenesten.no Rapport nr: StrFjo0913 Gradering: Åpen Strandsone Rapport
BESKRIVELSE AV ALGER
 BESKRIVELSE AV ALGER Brunalger Sauetang vokser ved flomålet øverst i fjæresonen. Den kommer under vann bare noen få timer hvert døgn og er derfor svært motstandsdyktig mot uttørking (figur til høyre).
BESKRIVELSE AV ALGER Brunalger Sauetang vokser ved flomålet øverst i fjæresonen. Den kommer under vann bare noen få timer hvert døgn og er derfor svært motstandsdyktig mot uttørking (figur til høyre).
Livet i fjæresonen. 1 Innledning
 Livet i fjæresonen 1 Innledning I denne rapporten vil jeg forsøke å belyse sentrale aspekter ved å dra på en ekskursjonen til fjæra for studere fjæresonen og de forskjellige tangartene man finner der.
Livet i fjæresonen 1 Innledning I denne rapporten vil jeg forsøke å belyse sentrale aspekter ved å dra på en ekskursjonen til fjæra for studere fjæresonen og de forskjellige tangartene man finner der.
Kan vi stole på resultater fra «liten N»?
 Kan vi stole på resultater fra «liten N»? Olav M. Kvalheim Universitetet i Bergen Plan for dette foredraget Hypotesetesting og p-verdier for å undersøke en variabel p-verdier når det er mange variabler
Kan vi stole på resultater fra «liten N»? Olav M. Kvalheim Universitetet i Bergen Plan for dette foredraget Hypotesetesting og p-verdier for å undersøke en variabel p-verdier når det er mange variabler
FAKTA. Tareskog nedbeitet av kråkeboller utenfor Midt-Norge: Beiting av grønne kråkeboller i tareskog. har tareskogen fått bestå urørt.
 FAKTA-ark Stiftelsen for naturforskning og kulturminneforskning er et nasjonalt og internasjonalt kompetansesenter innen miljøvernforskning. Stiftelsen har ca. 21 ansatte (1994) og omfatter NINA - Norsk
FAKTA-ark Stiftelsen for naturforskning og kulturminneforskning er et nasjonalt og internasjonalt kompetansesenter innen miljøvernforskning. Stiftelsen har ca. 21 ansatte (1994) og omfatter NINA - Norsk
Næringsstrøm i tareskogen
 Næringsstrøm i tareskogen av Kjell Magnus Norderhaug kjell.norderhaug@niva.no Importance of macrofauna in transferring kelp forest primary production to higher levels in the food web Doktorprosjekt gjennomført
Næringsstrøm i tareskogen av Kjell Magnus Norderhaug kjell.norderhaug@niva.no Importance of macrofauna in transferring kelp forest primary production to higher levels in the food web Doktorprosjekt gjennomført
Prolog. Stein Fredriksen. Biologisk Institutt. Universitetet i Oslo
 Prolog. Stein Fredriksen Biologisk Institutt Universitetet i Oslo Er forskningsmålene nådd innen delprogram A? Våre marine økosystemer Fokus på bentos Hovedmål: Å framskaffe ny viten om økosystemers struktur,
Prolog. Stein Fredriksen Biologisk Institutt Universitetet i Oslo Er forskningsmålene nådd innen delprogram A? Våre marine økosystemer Fokus på bentos Hovedmål: Å framskaffe ny viten om økosystemers struktur,
Littoral and shallow sublittoral algal vegetation from 8 sites around Svalbard
 Littoral and shallow sublittoral algal vegetation from 8 sites around Svalbard Siri Moy Stein Fredriksen, UiO Tove Gabrielsen, UNIS Wenche Eikrem, UiO Makroalger er en viktig komponent i det marine samfunn
Littoral and shallow sublittoral algal vegetation from 8 sites around Svalbard Siri Moy Stein Fredriksen, UiO Tove Gabrielsen, UNIS Wenche Eikrem, UiO Makroalger er en viktig komponent i det marine samfunn
ME Metode og statistikk Candidate 2511
 ME-400, forside Emnekode: ME-400 Emnenavn: Metode og statistikk Dato: 31. mai Varighet: 5 timer Tillatte hjelpemidler: Kalkulator (enkel type) Merknader: Besvar 3 av 4 oppgaver (Oppgavene teller likt)
ME-400, forside Emnekode: ME-400 Emnenavn: Metode og statistikk Dato: 31. mai Varighet: 5 timer Tillatte hjelpemidler: Kalkulator (enkel type) Merknader: Besvar 3 av 4 oppgaver (Oppgavene teller likt)
Kort overblikk over kurset sålangt
 Kort overblikk over kurset sålangt Kapittel 1: Deskriptiv statististikk for en variabel Kapittel 2: Deskriptiv statistikk for samvariasjon mellom to variable (regresjon) Kapittel 3: Metoder for å innhente
Kort overblikk over kurset sålangt Kapittel 1: Deskriptiv statististikk for en variabel Kapittel 2: Deskriptiv statistikk for samvariasjon mellom to variable (regresjon) Kapittel 3: Metoder for å innhente
OPPGAVESETTET BESTÅR AV 3 OPPGAVER PÅ 6 SIDER MERKNADER: Alle deloppgaver vektlegges likt.
 EKSAMEN I: MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 08. mai 2008 TILLATTE HJELPEMIDLER: Kalkulator: HP30S, Casio FX82 eller TI-30 Tabeller og formler i statistikk (Tapir forlag) OPPGAVESETTET
EKSAMEN I: MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 08. mai 2008 TILLATTE HJELPEMIDLER: Kalkulator: HP30S, Casio FX82 eller TI-30 Tabeller og formler i statistikk (Tapir forlag) OPPGAVESETTET
NOTAT Elvemuslingundersøkelser i Breivasselv, Grong kommune
 NOTAT Notat nr.: 1 06.11.2012 Dato Fylkesmannen i Nord-Trøndelag v/ Anton Rikstad Kopi til: Fra: Lars Erik Andersen Sweco Norge AS Bakgrunn: Sommeren 2011 ble det påvist et individ av elvemusling i Breivasselv,
NOTAT Notat nr.: 1 06.11.2012 Dato Fylkesmannen i Nord-Trøndelag v/ Anton Rikstad Kopi til: Fra: Lars Erik Andersen Sweco Norge AS Bakgrunn: Sommeren 2011 ble det påvist et individ av elvemusling i Breivasselv,
ST0202 Statistikk for samfunnsvitere
 ST0202 Statistikk for samfunnsvitere Bo Lindqvist Institutt for matematiske fag 2 Kap. 10: Inferens om to populasjoner Situasjon: Det er to populasjoner som vi ønsker å sammenligne. Vi trekker da et utvalg
ST0202 Statistikk for samfunnsvitere Bo Lindqvist Institutt for matematiske fag 2 Kap. 10: Inferens om to populasjoner Situasjon: Det er to populasjoner som vi ønsker å sammenligne. Vi trekker da et utvalg
Rødalger i ferskvann
 Rødalger i ferskvann Kunnskapsstatus med oversikt over arter i Norge (og Norden) Eli-Anne Lindstrøm, NIVA Jan Rueness, Universitetet i Oslo Maia Røst Kile, NIVA Mål Status for eksisterende kunnskap om
Rødalger i ferskvann Kunnskapsstatus med oversikt over arter i Norge (og Norden) Eli-Anne Lindstrøm, NIVA Jan Rueness, Universitetet i Oslo Maia Røst Kile, NIVA Mål Status for eksisterende kunnskap om
Oppgave 1. Det oppgis at dersom y ij er observasjon nummer j fra laboratorium i så er SSA = (y ij ȳ i ) 2 = 3.6080.
 2 = 3.6080.") EKSAMEN I: MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 28. FEBRUAR 2005 TILLATTE HJELPEMIDLER: KALKULATOR, TABELLER OG FORMLER I STATISTIKK (TAPIR FORLAG) OPPGAVESETTET BESTÅR AV 4 OPPGAVER PÅ
EKSAMEN I: MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 28. FEBRUAR 2005 TILLATTE HJELPEMIDLER: KALKULATOR, TABELLER OG FORMLER I STATISTIKK (TAPIR FORLAG) OPPGAVESETTET BESTÅR AV 4 OPPGAVER PÅ
Prøvehøsting av tare i Nord-Trøndelag. Henning Steen, Frithjof Moy, Torjan Bodvin Havforskningsinstituttet
 Prøvehøsting av tare i Nord-Trøndelag Henning Steen, Frithjof Moy, Torjan Bodvin Havforskningsinstituttet Stortare (Laminaria hyperborea) -Stortare er en av syv tarearter langs norskekysten, og dominerer
Prøvehøsting av tare i Nord-Trøndelag Henning Steen, Frithjof Moy, Torjan Bodvin Havforskningsinstituttet Stortare (Laminaria hyperborea) -Stortare er en av syv tarearter langs norskekysten, og dominerer
[2D] Målet for opplæringa er at elevane skal kunne gjere greie for korleis ytre faktorar verkar inn på fotosyntesen.
![[2D] Målet for opplæringa er at elevane skal kunne gjere greie for korleis ytre faktorar verkar inn på fotosyntesen.](/thumbs/50/26953017.jpg "[2D] Målet for opplæringa er at elevane skal kunne gjere greie for korleis ytre faktorar verkar inn på fotosyntesen.") Bi2 «Energiomsetning» [2D] Målet for opplæringa er at elevane skal kunne gjere greie for korleis ytre faktorar verkar inn på fotosyntesen. Oppgave 1a, 1b, 1c V1984 Kurven viser hvordan C0 2 -innholdet
Bi2 «Energiomsetning» [2D] Målet for opplæringa er at elevane skal kunne gjere greie for korleis ytre faktorar verkar inn på fotosyntesen. Oppgave 1a, 1b, 1c V1984 Kurven viser hvordan C0 2 -innholdet
HØGSKOLEN I STAVANGER
 EKSAMEN I: MOT0 STATISTISKE METODER VARIGHET: TIMER DATO:. NOVEMBER 00 TILLATTE HJELPEMIDLER: KALKULATOR, TABELLER OG FORMLER I STATISTIKK (TAPIR FORLAG) OPPGAVESETTET BESTÅR AV OPPGAVER PÅ 7 SIDER HØGSKOLEN
EKSAMEN I: MOT0 STATISTISKE METODER VARIGHET: TIMER DATO:. NOVEMBER 00 TILLATTE HJELPEMIDLER: KALKULATOR, TABELLER OG FORMLER I STATISTIKK (TAPIR FORLAG) OPPGAVESETTET BESTÅR AV OPPGAVER PÅ 7 SIDER HØGSKOLEN
Kvinne Antall Tabell 1a. Antall migreneanfall i året før kvinnene fikk medisin.
 Eksamen STAT100, Høst 2011 (lettere revidert). Tillatte hjelpemidler: C3: alle typer kalkulator, alle andre hjelpemidler Ved alle hypotesetester skal både nullhypotese og alternativ hypotese skrives ned.
Eksamen STAT100, Høst 2011 (lettere revidert). Tillatte hjelpemidler: C3: alle typer kalkulator, alle andre hjelpemidler Ved alle hypotesetester skal både nullhypotese og alternativ hypotese skrives ned.
Eksamensoppgave i TMA4255 Anvendt statistikk
 Institutt for matematiske fag Eksamensoppgave i TMA4255 Anvendt statistikk Faglig kontakt under eksamen: Anna Marie Holand Tlf: 951 38 038 Eksamensdato: 30. mai 2014 Eksamenstid (fra til): 09:00-13:00
Institutt for matematiske fag Eksamensoppgave i TMA4255 Anvendt statistikk Faglig kontakt under eksamen: Anna Marie Holand Tlf: 951 38 038 Eksamensdato: 30. mai 2014 Eksamenstid (fra til): 09:00-13:00
Snøtetthet. Institutt for matematiske fag, NTNU 15. august Notat for TMA4240/TMA4245 Statistikk
 Snøtetthet Notat for TMA424/TMA4245 Statistikk Institutt for matematiske fag, NTNU 5. august 22 I forbindelse med varsling av om, klimaforskning og særlig kraftproduksjon er det viktig å kunne anslå hvor
Snøtetthet Notat for TMA424/TMA4245 Statistikk Institutt for matematiske fag, NTNU 5. august 22 I forbindelse med varsling av om, klimaforskning og særlig kraftproduksjon er det viktig å kunne anslå hvor
ST0103 Brukerkurs i statistikk Forelesning 26, 18. november 2016 Kapittel 8: Sammenligning av grupper
 ST0103 Brukerkurs i statistikk Forelesning 26, 18. november 2016 Kapittel 8: Sammenligning av grupper Bo Lindqvist Institutt for matematiske fag 2 Kapittel 8: Sammenligning av grupper Situasjon: Vi ønsker
ST0103 Brukerkurs i statistikk Forelesning 26, 18. november 2016 Kapittel 8: Sammenligning av grupper Bo Lindqvist Institutt for matematiske fag 2 Kapittel 8: Sammenligning av grupper Situasjon: Vi ønsker
Tilstandsregistrering Ålegrass, Indre Viksfjord,
 Tilstandsregistrering Ålegrass, Indre Viksfjord, 29.09.2017 TILSTANDSREGISTRERING ÅLEGRASS, INDRE VIKSFJORD, 29.09.2017 SIDE 1 Innledning Ålegrass inngår som et av de biologiske kvalitetselementene som
Tilstandsregistrering Ålegrass, Indre Viksfjord, 29.09.2017 TILSTANDSREGISTRERING ÅLEGRASS, INDRE VIKSFJORD, 29.09.2017 SIDE 1 Innledning Ålegrass inngår som et av de biologiske kvalitetselementene som
Vurdering av eutrofieringssituasjonen i kystområder, med særlig fokus på Hardangerfjorden og Boknafjorden. Stein Fredriksen Universitetet i Oslo
 Vurdering av eutrofieringssituasjonen i kystområder, med særlig fokus på Hardangerfjorden og Boknafjorden Stein Fredriksen Universitetet i Oslo Ekspertgruppen Kjersti Sjøtun (Universitetet i Bergen) Hartvig
Vurdering av eutrofieringssituasjonen i kystområder, med særlig fokus på Hardangerfjorden og Boknafjorden Stein Fredriksen Universitetet i Oslo Ekspertgruppen Kjersti Sjøtun (Universitetet i Bergen) Hartvig
Profil Lavpris Supermarked Hypermarked Totalt. Coop Prix 4 4. Coop Extra 13 5. Coop Mega 7 7. Coop Obs 5 13. Rimi 24 24. Ica Supermarked 7 7
 Vedlegg 1 - Regresjonsanalyser 1 Innledning og formål (1) Konkurransetilsynet har i forbindelse med Vedtak 2015-24, (heretter "Vedtaket") utført kvantitative analyser på data fra kundeundersøkelsen. I
Vedlegg 1 - Regresjonsanalyser 1 Innledning og formål (1) Konkurransetilsynet har i forbindelse med Vedtak 2015-24, (heretter "Vedtaket") utført kvantitative analyser på data fra kundeundersøkelsen. I
Oppgave 1. X 1 B(n 1, p 1 ) X 2. Vi er interessert i forskjellen i andeler p 1 p 2, som vi estimerer med. p 1 p 2 = X 1. n 1 n 2.
 X 2. Vi er interessert i forskjellen i andeler p 1 p 2, som vi estimerer med. p 1 p 2 = X 1. n 1 n 2.") Løsningsforslag til eksamen i MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 17 november 2008 TILLATTE HJELPEMIDLER: Kalkulator: HP30S, Casio FX82 eller TI-30 Tabeller og formler i statistikk Tapir
Løsningsforslag til eksamen i MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 17 november 2008 TILLATTE HJELPEMIDLER: Kalkulator: HP30S, Casio FX82 eller TI-30 Tabeller og formler i statistikk Tapir
I enkel lineær regresjon beskrev linja. μ y = β 0 + β 1 x
 Multiple regresjon Her utvider vi perspektivet for enkel lineær regresjon til også å omfatte flere forklaringsvariable.det er fortsatt en responsvariabel. Måten dette gjøre på er nokså naturlig. Prediktoren
Multiple regresjon Her utvider vi perspektivet for enkel lineær regresjon til også å omfatte flere forklaringsvariable.det er fortsatt en responsvariabel. Måten dette gjøre på er nokså naturlig. Prediktoren
Utviklingen av tarekråkebollesituasjonen
 Utviklingen av tarekråkebollesituasjonen i Nordland Fylkesmannsamlingen i Bodø 12. juni 2012 Eli Rinde, Hartvig Christie (NIVA) 1 Oversikt Hvor finner en stortare og sukkertare? Hvor og hvorfor har tareskog
Utviklingen av tarekråkebollesituasjonen i Nordland Fylkesmannsamlingen i Bodø 12. juni 2012 Eli Rinde, Hartvig Christie (NIVA) 1 Oversikt Hvor finner en stortare og sukkertare? Hvor og hvorfor har tareskog
Bioindikatorer i kystvann. Indikatorer og påvirkningstyper
 Bioindikatorer i kystvann Indikatorer og påvirkningstyper 29. mars 2012 1 Innhold 1. Bakgrunn bioindikatorer 2. Økoregioner og vanntyper 3. Hvilke bioindikatorer har vi 4. Når/Hvor skal bioindikatorene
Bioindikatorer i kystvann Indikatorer og påvirkningstyper 29. mars 2012 1 Innhold 1. Bakgrunn bioindikatorer 2. Økoregioner og vanntyper 3. Hvilke bioindikatorer har vi 4. Når/Hvor skal bioindikatorene
Løsningsforslag ECON 2130 Obligatorisk semesteroppgave 2017 vår
 Løsningsforslag ECON 130 Obligatorisk semesteroppgave 017 vår Andreas Myhre Oppgave 1 1. (i) Siden X og Z er uavhengige, vil den simultane fordelingen mellom X og Z kunne skrives som: f(x, z) = P(X = x
Løsningsforslag ECON 130 Obligatorisk semesteroppgave 017 vår Andreas Myhre Oppgave 1 1. (i) Siden X og Z er uavhengige, vil den simultane fordelingen mellom X og Z kunne skrives som: f(x, z) = P(X = x
TMA4245 Statistikk Eksamen desember 2016
 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag TMA4245 Statistikk Eksamen desember 2016 Oppgave 1 En bedrift produserer elektriske komponenter. Komponentene kan ha to typer
Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag TMA4245 Statistikk Eksamen desember 2016 Oppgave 1 En bedrift produserer elektriske komponenter. Komponentene kan ha to typer
1.1 Jakten på en sammenheng
 1.1 Jakten på en sammenheng Viktige definisjoner fra 8. klasse: ATOMER MOLEKYL KJEMISK FORBINDELSE ENERGI TEMPERATUR MASSE VOLUM MASSETETTHET KREFTER GRAVITASJONSKRAFT (Tyngdekraft) ELEKTROMAGNETISK STRÅLING
1.1 Jakten på en sammenheng Viktige definisjoner fra 8. klasse: ATOMER MOLEKYL KJEMISK FORBINDELSE ENERGI TEMPERATUR MASSE VOLUM MASSETETTHET KREFTER GRAVITASJONSKRAFT (Tyngdekraft) ELEKTROMAGNETISK STRÅLING
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord. Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 8.
 Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 8. mai 2014 26. juni 2014 1 Det kommunale samarbeidsorganet «Fagrådet for
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 8. mai 2014 26. juni 2014 1 Det kommunale samarbeidsorganet «Fagrådet for
NOTAT Tiltak for elvemusling i Hitra kommune Bruelva
 NOTAT Tiltak for elvemusling i Hitra kommune Bruelva Notat nr.: Dato 1 25.07.2011 Til: Navn Firma Fork. Anmerkning Hitra kommune v/arne Aarnes Fylkesmannen i Sør- Trøndelag v/ Kari Tønset Guttvik Norges
NOTAT Tiltak for elvemusling i Hitra kommune Bruelva Notat nr.: Dato 1 25.07.2011 Til: Navn Firma Fork. Anmerkning Hitra kommune v/arne Aarnes Fylkesmannen i Sør- Trøndelag v/ Kari Tønset Guttvik Norges
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 18.
 Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 18. august 2014 16. oktober 2014 1 Det kommunale samarbeidsorganet «Fagrådet
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 18. august 2014 16. oktober 2014 1 Det kommunale samarbeidsorganet «Fagrådet
Forfattere: Jenny Manne og Vilrun Otre Røssummoen, Bergen katedralskole
 SPISS Tidsskrift for elever med teknologi og forskningslære i videregående skole på PC og mobil Forfattere: Jenny Manne og Vilrun Otre Røssummoen, Bergen katedralskole Abstrakt I vårt forsøk har vi undersøkt
SPISS Tidsskrift for elever med teknologi og forskningslære i videregående skole på PC og mobil Forfattere: Jenny Manne og Vilrun Otre Røssummoen, Bergen katedralskole Abstrakt I vårt forsøk har vi undersøkt
UNIVERSITETET I OSLO
 UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen i: STK1120 Statistiske metoder og dataanalyse 2. Eksamensdag: Mandag 30. mai 2005. Tid for eksamen: 14.30 17.30. Oppgavesettet er
UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen i: STK1120 Statistiske metoder og dataanalyse 2. Eksamensdag: Mandag 30. mai 2005. Tid for eksamen: 14.30 17.30. Oppgavesettet er
QED 1 7. Matematikk for grunnskolelærerutdanningen. Bind 2. Fasit kapittel 4 Statistikk og kvantitativ metode
 QED 1 7 Matematikk for grunnskolelærerutdanningen Bind 2 Fasit kapittel 4 Statistikk og kvantitativ metode Kapittel 4 Oppgave 1 La være antall øyne på terningen. a) Vi får følgende sannsynlighetsfordeling
QED 1 7 Matematikk for grunnskolelærerutdanningen Bind 2 Fasit kapittel 4 Statistikk og kvantitativ metode Kapittel 4 Oppgave 1 La være antall øyne på terningen. a) Vi får følgende sannsynlighetsfordeling
TMA4240 Statistikk Høst 2016
 TMA4240 Statistikk Høst 2016 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Anbefalt øving 11 Oppgavene i denne øvingen dreier seg om hypotesetesting og sentrale begreper
TMA4240 Statistikk Høst 2016 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Anbefalt øving 11 Oppgavene i denne øvingen dreier seg om hypotesetesting og sentrale begreper
NOTAT. SMS Sandbukta Moss Såstad. Temanotat Kartlegging av strømningsforhold. Sammendrag
 NOTAT Oppdrag 960168 Sandbukta Moss Såstad, Saks. Nr 201600206 Kunde Bane NOR Notat nr. Forurenset grunn/002-2017 Dato 17-03-2017 Til Fra Kopi Ingunn Helen Bjørnstad/ Bane NOR Rambøll Sweco ANS/ Michael
NOTAT Oppdrag 960168 Sandbukta Moss Såstad, Saks. Nr 201600206 Kunde Bane NOR Notat nr. Forurenset grunn/002-2017 Dato 17-03-2017 Til Fra Kopi Ingunn Helen Bjørnstad/ Bane NOR Rambøll Sweco ANS/ Michael
ST0103 Brukerkurs i statistikk Høst 2014
 Norges teknisk naturvitenskapelige universitet Institutt for matematiske fag ST0103 Brukerkurs i statistikk Høst 2014 Løsningsforslag Øving 1 2.1 Frekvenstabell For å lage en frekvenstabell må vi telle
Norges teknisk naturvitenskapelige universitet Institutt for matematiske fag ST0103 Brukerkurs i statistikk Høst 2014 Løsningsforslag Øving 1 2.1 Frekvenstabell For å lage en frekvenstabell må vi telle
Løsningsforslag. n X. n X 1 i=1 (X i X) 2 og SY 2 = 1 ny S 2 X + S2 Y
 2 og SY 2 = 1 ny S 2 X + S2 Y") Statistiske metoder 1 høsten 004. Løsningsforslag Oppgave 1: a) Begge normalplottene gir punkter som ligger omtrent på ei rett linje så antagelsen om normalfordeling ser ut til å holde. Konfidensintervall
Statistiske metoder 1 høsten 004. Løsningsforslag Oppgave 1: a) Begge normalplottene gir punkter som ligger omtrent på ei rett linje så antagelsen om normalfordeling ser ut til å holde. Konfidensintervall
MAREANO-biomangfold. Lene Buhl Mortensen/J.H. Fosså. Foreløpige resultater fra 2006
 MAREANO-biomangfold Lene Buhl Mortensen/J.H. Fosså Foreløpige resultater fra 2006 Hvor, hva og hvordan materialet er samlet inn Resultater fra videoundersøkelsene Effekter av fiskeri på økosystemet Eksempler
MAREANO-biomangfold Lene Buhl Mortensen/J.H. Fosså Foreløpige resultater fra 2006 Hvor, hva og hvordan materialet er samlet inn Resultater fra videoundersøkelsene Effekter av fiskeri på økosystemet Eksempler
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord. Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 8.
 Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 8. desember 2014 14. januar 2015 1 Det kommunale samarbeidsorganet «Fagrådet
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 8. desember 2014 14. januar 2015 1 Det kommunale samarbeidsorganet «Fagrådet
Kontroller at oppgavesettet er komplett før du begynner å besvare spørsmålene. Ved sensuren teller alle delspørsmål likt.
 Eksamen i: MET040 Statistikk for økonomer Eksamensdag: 4 november 2008 Tid for eksamen: 09.00-13.00 Oppgavesettet er på 4 sider. Tillatte hjelpemidler: Alle trykte eller egenskrevne hjelpemidler og kalkulator.
Eksamen i: MET040 Statistikk for økonomer Eksamensdag: 4 november 2008 Tid for eksamen: 09.00-13.00 Oppgavesettet er på 4 sider. Tillatte hjelpemidler: Alle trykte eller egenskrevne hjelpemidler og kalkulator.
Bærekraftig utvikling - forskerspiren. Maria Sviland, Skolelaboratoriet NTNU
 Bærekraftig utvikling - forskerspiren Maria Sviland, Skolelaboratoriet NTNU 1 2 Forskerspiren Forskerspiren Kompetansemål etter Vg1 studieforberedende utdanningsprogram ( - Naturfag i vidregående opplæring)
Bærekraftig utvikling - forskerspiren Maria Sviland, Skolelaboratoriet NTNU 1 2 Forskerspiren Forskerspiren Kompetansemål etter Vg1 studieforberedende utdanningsprogram ( - Naturfag i vidregående opplæring)
EKSAMEN I FAG TMA4260 INDUSTRIELL STATISTIKK
 Norges teknisk naturvitenskapelige universitet Institutt for matematiske fag Side 1 av 12 Faglig kontakt under eksamen: Bo Lindqvist Tlf. 975 89 418 EKSAMEN I FAG TMA4260 INDUSTRIELL STATISTIKK Onsdag
Norges teknisk naturvitenskapelige universitet Institutt for matematiske fag Side 1 av 12 Faglig kontakt under eksamen: Bo Lindqvist Tlf. 975 89 418 EKSAMEN I FAG TMA4260 INDUSTRIELL STATISTIKK Onsdag
Påvirker sur nedbør planter sin spiringsprosent og vekst?
 SPISS Naturfaglige artikler av elever i videregående opplæring Påvirker sur nedbør planter sin spiringsprosent og vekst? Forfattere: Elias Jegervatn Severinsen, Bjørn Werner Laug, Tobias Furuvald Sunde,
SPISS Naturfaglige artikler av elever i videregående opplæring Påvirker sur nedbør planter sin spiringsprosent og vekst? Forfattere: Elias Jegervatn Severinsen, Bjørn Werner Laug, Tobias Furuvald Sunde,
Når Merge sort og Insertion sort samarbeider
 Når Merge sort og Insertion sort samarbeider Lars Sydnes 8. november 2014 1 Innledning Her skal vi undersøke to algoritmer som brukes til å sortere lister, Merge sort og Insertion sort. Det at Merge sort
Når Merge sort og Insertion sort samarbeider Lars Sydnes 8. november 2014 1 Innledning Her skal vi undersøke to algoritmer som brukes til å sortere lister, Merge sort og Insertion sort. Det at Merge sort
6.2 Signifikanstester
 6.2 Signifikanstester Konfidensintervaller er nyttige når vi ønsker å estimere en populasjonsparameter Signifikanstester er nyttige dersom vi ønsker å teste en hypotese om en parameter i en populasjon
6.2 Signifikanstester Konfidensintervaller er nyttige når vi ønsker å estimere en populasjonsparameter Signifikanstester er nyttige dersom vi ønsker å teste en hypotese om en parameter i en populasjon
Fremmede arter: Stillehavsøsters. Kartlegging, økologisk effekt og tiltak
 Fremmede arter: Stillehavsøsters Kartlegging, økologisk effekt og tiltak Stillehavsøsters Introdusert art (første observasjoner i Norge 2003) I dag; tilstede langs hele kysten av Skagerrak Kjønnsmoden
Fremmede arter: Stillehavsøsters Kartlegging, økologisk effekt og tiltak Stillehavsøsters Introdusert art (første observasjoner i Norge 2003) I dag; tilstede langs hele kysten av Skagerrak Kjønnsmoden
NOTAT. SMS Sandbukta Moss Såstad. Temanotat Økologisk tilstandsklassifisering av ålegras i Mossesundet og Verlebukta. Sammendrag
 NOTAT Oppdrag 960168 Sandbukta Moss Såstad, Saks. Nr 2018002800 Kunde Bane NOR Notat nr. Not_002_20180323_Bane NOR_2018002800_Temanotat - Ålegras Dato 23.03.2018 Til Fra Kopi Ingunn Helen Bjørnstad/ Bane
NOTAT Oppdrag 960168 Sandbukta Moss Såstad, Saks. Nr 2018002800 Kunde Bane NOR Notat nr. Not_002_20180323_Bane NOR_2018002800_Temanotat - Ålegras Dato 23.03.2018 Til Fra Kopi Ingunn Helen Bjørnstad/ Bane
Statistikk for språk- og musikkvitere 1
 Statistikk for språk- og musikkvitere 1 Mitt navn: Åsne Haaland, Vitenskapelig databehandling USIT Ikke nøl, avbryt med spørsmål! Hva oppnår en med statistikk? Få oversikt over data: typisk verdi, spredning,
Statistikk for språk- og musikkvitere 1 Mitt navn: Åsne Haaland, Vitenskapelig databehandling USIT Ikke nøl, avbryt med spørsmål! Hva oppnår en med statistikk? Få oversikt over data: typisk verdi, spredning,
Statistikk 1. Nico Keilman. ECON 2130 Vår 2014
 Statistikk 1 Nico Keilman ECON 2130 Vår 2014 Pensum Kap 1-7.3.6 fra Løvås «Statistikk for universiteter og høgskoler» 3. utgave 2013 (eventuelt 2. utgave) Se overspringelsesliste på emnesiden Supplerende
Statistikk 1 Nico Keilman ECON 2130 Vår 2014 Pensum Kap 1-7.3.6 fra Løvås «Statistikk for universiteter og høgskoler» 3. utgave 2013 (eventuelt 2. utgave) Se overspringelsesliste på emnesiden Supplerende
Overvåking av elvemusling i Strømselva, Averøy kommune Forundersøkelse
 Overvåking av elvemusling i Strømselva, Averøy kommune Miljøfaglig Utredning, rapport 2006:48 Miljøfaglig Utredning 2 Miljøfaglig Utredning AS Rapport 2006:48 Utførende institusjon: Miljøfaglig Utredning
Overvåking av elvemusling i Strømselva, Averøy kommune Miljøfaglig Utredning, rapport 2006:48 Miljøfaglig Utredning 2 Miljøfaglig Utredning AS Rapport 2006:48 Utførende institusjon: Miljøfaglig Utredning
KYSTOVERVÅKINGSPROGRAMMET
 KYSTOVERVÅKINGSPROGRAMMET HARDBUNNSSAMFUNN DATARAPPORT 2008 2732 2010 Statlig program for forurensningsovervåking Langtidsovervåking av miljøkvaliteten i kystområdene av Norge. Kystovervåkingsprogrammet
KYSTOVERVÅKINGSPROGRAMMET HARDBUNNSSAMFUNN DATARAPPORT 2008 2732 2010 Statlig program for forurensningsovervåking Langtidsovervåking av miljøkvaliteten i kystområdene av Norge. Kystovervåkingsprogrammet
+ S2 Y ) 2. = 6.737 6 (avrundet nedover til nærmeste heltall) n Y 1
 2. = 6.737 6 (avrundet nedover til nærmeste heltall) n Y 1") Løsningsforslag for: MOT10 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 6. november 007 TILLATTE HJELPEMIDLER: Kalkulator: HP0S, Casio FX8 eller TI-0 Tabeller og formler i statistikk (Tapir forlag) MERKNADER:
Løsningsforslag for: MOT10 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 6. november 007 TILLATTE HJELPEMIDLER: Kalkulator: HP0S, Casio FX8 eller TI-0 Tabeller og formler i statistikk (Tapir forlag) MERKNADER:
Godkjent prosjektansvarlig:
 Olje & Energi Seksjon for Materialteknologi Porsgrunn MATERIALTEKNISK RAPPORT Gradering: Internt Tittel: Westerns forlis. Sakkyndig uttalelse vedrørende hull i aluminium bakkdekk. Forfatter(e): Håkon Leth-Olsen
Olje & Energi Seksjon for Materialteknologi Porsgrunn MATERIALTEKNISK RAPPORT Gradering: Internt Tittel: Westerns forlis. Sakkyndig uttalelse vedrørende hull i aluminium bakkdekk. Forfatter(e): Håkon Leth-Olsen
Oppgave 1. T = 9 Hypotesetest for å teste om kolesterolnivået har endret seg etter dietten: T observert = 2.16 0
 Løsningsforslag til eksamen i MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 08. mai 2008 TILLATTE HJELPEMIDLER: Kalkulator: HP30S, Casio FX82 eller TI-30 Tabeller og formler i statistikk (Tapir
Løsningsforslag til eksamen i MOT310 STATISTISKE METODER 1 VARIGHET: 4 TIMER DATO: 08. mai 2008 TILLATTE HJELPEMIDLER: Kalkulator: HP30S, Casio FX82 eller TI-30 Tabeller og formler i statistikk (Tapir
Kapittel 3: Studieopplegg
 Oversikt over pensum Kapittel 1: Empirisk fordeling for en variabel o Begrepet fordeling o Mål for senter (gj.snitt, median) + persentiler/kvartiler o Mål for spredning (Standardavvik s, IQR) o Outliere
Oversikt over pensum Kapittel 1: Empirisk fordeling for en variabel o Begrepet fordeling o Mål for senter (gj.snitt, median) + persentiler/kvartiler o Mål for spredning (Standardavvik s, IQR) o Outliere
i x i
 TMA4245 Statistikk Vår 2016 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Anbefalte oppgaver 11, blokk II Oppgavene i denne øvingen dreier seg om hypotesetesting og sentrale
TMA4245 Statistikk Vår 2016 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Anbefalte oppgaver 11, blokk II Oppgavene i denne øvingen dreier seg om hypotesetesting og sentrale
Sentralmål og spredningsmål
 Sentralmål og spredningsmål av Peer Andersen Peer Andersen 2014 Sentralmål og spredningsmål i statistikk I dette notatet skal vi se på de viktigste momentene om sentralmål og spredningsmål slik de blir
Sentralmål og spredningsmål av Peer Andersen Peer Andersen 2014 Sentralmål og spredningsmål i statistikk I dette notatet skal vi se på de viktigste momentene om sentralmål og spredningsmål slik de blir
Forskningsmetoder i menneske-maskin interaksjon
 Forskningsmetoder i menneske-maskin interaksjon Kapittel 2- Eksperimentell forskning Oversikt Typer atferdsforskning Forskningshypoteser Grunnleggende om eksperimentell forskning Signifikanstesting Begrensninger
Forskningsmetoder i menneske-maskin interaksjon Kapittel 2- Eksperimentell forskning Oversikt Typer atferdsforskning Forskningshypoteser Grunnleggende om eksperimentell forskning Signifikanstesting Begrensninger
Eksamensoppgave i TMA4240 Statistikk
 Institutt for matematiske fag Eksamensoppgave i TMA4240 Statistikk Faglig kontakt under eksamen: Mette Langaas a, Ingelin Steinsland b, Geir-Arne Fuglstad c Tlf: a 988 47 649, b 926 63 096, c 452 70 806
Institutt for matematiske fag Eksamensoppgave i TMA4240 Statistikk Faglig kontakt under eksamen: Mette Langaas a, Ingelin Steinsland b, Geir-Arne Fuglstad c Tlf: a 988 47 649, b 926 63 096, c 452 70 806
Marinbiologi. Nyttig litteratur - kilder til kunnskap om livet under vann
 Marinbiologi Nyttig litteratur - kilder til kunnskap om livet under vann Tips til nyttig litteratur Vi i utvalg for marinbiologi ønsker å gi tips til litteratur som kan være nyttig i forbindelse med den
Marinbiologi Nyttig litteratur - kilder til kunnskap om livet under vann Tips til nyttig litteratur Vi i utvalg for marinbiologi ønsker å gi tips til litteratur som kan være nyttig i forbindelse med den
NOTAT Tiltak for elvemusling på Hitra Langvasselva
 NOTAT Tiltak for elvemusling på Hitra Langvasselva Notat nr.: Dato 1 26.07.2011 Til: Navn Firma Fork. Anmerkning Hitra kommune v/arne Aarnes Fylkesmannen i Sør- Trøndelag v/ Kari Tønset Guttvik Norges
NOTAT Tiltak for elvemusling på Hitra Langvasselva Notat nr.: Dato 1 26.07.2011 Til: Navn Firma Fork. Anmerkning Hitra kommune v/arne Aarnes Fylkesmannen i Sør- Trøndelag v/ Kari Tønset Guttvik Norges
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord. Miljøovervåking av Indre Oslofjord
 Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Resultater fra tokt 14-5-2013 1. juli 2013 1 Det kommunale samarbeidsorganet Fagrådet for indre Oslofjord
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Resultater fra tokt 14-5-2013 1. juli 2013 1 Det kommunale samarbeidsorganet Fagrådet for indre Oslofjord
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord. Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 15.
 Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 15. oktober 2014 13. november 2014 1 Det kommunale samarbeidsorganet «Fagrådet
Fagrådet for vann- og avløpsteknisk samarbeid i indre Oslofjord Miljøovervåking av Indre Oslofjord Rapport for tokt gjennomført 15. oktober 2014 13. november 2014 1 Det kommunale samarbeidsorganet «Fagrådet
Andre sett med obligatoriske oppgaver i STK1110 høsten 2010
 Andre sett med obligatoriske oppgaver i STK1110 høsten 2010 Dette er det andre settet med obligatoriske oppgaver i STK1110 høsten 2010. Oppgavesettet består av fire oppgaver. Det er valgfritt om du vil
Andre sett med obligatoriske oppgaver i STK1110 høsten 2010 Dette er det andre settet med obligatoriske oppgaver i STK1110 høsten 2010. Oppgavesettet består av fire oppgaver. Det er valgfritt om du vil
ØVINGER 2017 Løsninger til oppgaver. Øving 1
 ØVINGER 017 Løsninger til oppgaver Øving 1.1. Frekvenstabell For å lage en frekvenstabell må vi telle antall observasjoner av hvert antall henvendelser. Siden antall henvendelser på en gitt dag alltid
ØVINGER 017 Løsninger til oppgaver Øving 1.1. Frekvenstabell For å lage en frekvenstabell må vi telle antall observasjoner av hvert antall henvendelser. Siden antall henvendelser på en gitt dag alltid
Kan vi forutse en pendels bevegelse, før vi har satt den i sving?
 Gjør dette hjemme 6 #8 Kan vi forutse en pendels bevegelse, før vi har satt den i sving? Skrevet av: Kristian Sørnes Dette eksperimentet ser på hvordan man finner en matematisk formel fra et eksperiment,
Gjør dette hjemme 6 #8 Kan vi forutse en pendels bevegelse, før vi har satt den i sving? Skrevet av: Kristian Sørnes Dette eksperimentet ser på hvordan man finner en matematisk formel fra et eksperiment,
Kunstige rev i Nordfjorden ved Risør Rapport fra undersøkelser rundt to enheter av Runde reef Fra Hartvig Christie, Forsker NIVA
 1 Kunstige rev i Nordfjorden ved Risør Rapport fra undersøkelser rundt to enheter av Runde reef Fra Hartvig Christie, Forsker NIVA To kunstige rev-konstruksjoner fra Reef Systems as ble plassert ut på
1 Kunstige rev i Nordfjorden ved Risør Rapport fra undersøkelser rundt to enheter av Runde reef Fra Hartvig Christie, Forsker NIVA To kunstige rev-konstruksjoner fra Reef Systems as ble plassert ut på
Flervalgsoppgaver: Gassutveksling i planter
 Flervalgsoppgaver gassutveksling planter Hver oppgave har ett riktig svaralternativ. Gassutveksling planter 1 Hva er riktig om spalteåpninger? A) De er åpne om natta i de fleste planter. B) De er åpne
Flervalgsoppgaver gassutveksling planter Hver oppgave har ett riktig svaralternativ. Gassutveksling planter 1 Hva er riktig om spalteåpninger? A) De er åpne om natta i de fleste planter. B) De er åpne
BKE - Makroalger. Are Pedersen. 2. april
 BKE - Makroalger 2. april 2014 1 Krav til data makroalger Innsamling i sommerperioden mai/juni - august/september (perioden tilpasses for de ulike økoregioner) Innsamling bør gjøres årlig, - min. hvert
BKE - Makroalger 2. april 2014 1 Krav til data makroalger Innsamling i sommerperioden mai/juni - august/september (perioden tilpasses for de ulike økoregioner) Innsamling bør gjøres årlig, - min. hvert
Eksamensoppgave i TMA4255 Anvendt statistikk
 Institutt for matematiske fag Eksamensoppgave i TMA4255 Anvendt statistikk Faglig kontakt under eksamen: Anna Marie Holand Tlf: 951 38 038 Eksamensdato: 16. mai 2015 Eksamenstid (fra til): 09:00-13:00
Institutt for matematiske fag Eksamensoppgave i TMA4255 Anvendt statistikk Faglig kontakt under eksamen: Anna Marie Holand Tlf: 951 38 038 Eksamensdato: 16. mai 2015 Eksamenstid (fra til): 09:00-13:00
Rådgivende Biologer AS
 Rådgivende Biologer AS NOTAT Tilstandsovervåkning av ålegraseng i Eltarvågen ved lokaliteten Rennaren i Rennesøy kommune 2018 Av: Joar Tverberg Til: Grieg Seafood Rogaland AS ved Liv Marit Årseth Dato:
Rådgivende Biologer AS NOTAT Tilstandsovervåkning av ålegraseng i Eltarvågen ved lokaliteten Rennaren i Rennesøy kommune 2018 Av: Joar Tverberg Til: Grieg Seafood Rogaland AS ved Liv Marit Årseth Dato:
Bekreftelse på utført resipientundersøkelse ved Kvithylla, samt foreløpige resultater
 Kontoradresse: Strandaveien, Lauvsnes Postadresse: Lauvsneshaugen 7, 7770 Flatanger Telefon: 74 28 84 30 Mobil: 909 43 493 E-post: post@aqua-kompetanse.no www.aqua-kompetanse.no Bankgiro: 4400.07.25541
Kontoradresse: Strandaveien, Lauvsnes Postadresse: Lauvsneshaugen 7, 7770 Flatanger Telefon: 74 28 84 30 Mobil: 909 43 493 E-post: post@aqua-kompetanse.no www.aqua-kompetanse.no Bankgiro: 4400.07.25541
Bokmål. Eksamen i: Stat100 Statistikk Tid: 18. mai Emneansvarlig: Trygve Almøy:
 Bokmål Institutt: IKBM Eksamen i: Stat100 Statistikk Tid: 18. mai 2010 09.00-12.30 Emneansvarlig: Trygve Almøy: 64 96 58 20 Tillatte hjelpemidler: C3: alle typer kalkulatorer, alle andre hjelpemiddel Oppgaveteksten
Bokmål Institutt: IKBM Eksamen i: Stat100 Statistikk Tid: 18. mai 2010 09.00-12.30 Emneansvarlig: Trygve Almøy: 64 96 58 20 Tillatte hjelpemidler: C3: alle typer kalkulatorer, alle andre hjelpemiddel Oppgaveteksten
Hypotesetesting. Hvorfor og hvordan? Gardermoen 21. april 2016 Ørnulf Borgan. H. Aschehoug & Co Sehesteds gate 3, 0102 Oslo Tlf:
 Hypotesetesting Hvorfor og hvordan? Gardermoen 21. april 2016 Ørnulf Borgan H. Aschehoug & Co Sehesteds gate 3, 0102 Oslo Tlf: 22 400 400 www.aschehoug.no 1 Oversikt Sannsynlighetsregning og statistikk
Hypotesetesting Hvorfor og hvordan? Gardermoen 21. april 2016 Ørnulf Borgan H. Aschehoug & Co Sehesteds gate 3, 0102 Oslo Tlf: 22 400 400 www.aschehoug.no 1 Oversikt Sannsynlighetsregning og statistikk
Eksamen PSY1011/PSYPRO4111: Sensorveiledning
 Eksamen PSY1011/PSYPRO4111 1. Hva vil det si at et instrument for å måle angst er valid? Hvordan kan man undersøke validiteten til instrumentet? 2. Hva vil det si at et resultat er statistisk signifikant?
Eksamen PSY1011/PSYPRO4111 1. Hva vil det si at et instrument for å måle angst er valid? Hvordan kan man undersøke validiteten til instrumentet? 2. Hva vil det si at et resultat er statistisk signifikant?
UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet
 UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Underveiseksamen i: STK1000 Innføring i anvendt statistikk. Eksamensdag: Fredag 13.10.2006. Tid for eksamen: Kl. 09.00 11.00. Tillatte hjelpemidler:
UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Underveiseksamen i: STK1000 Innføring i anvendt statistikk. Eksamensdag: Fredag 13.10.2006. Tid for eksamen: Kl. 09.00 11.00. Tillatte hjelpemidler:
OVERFLATE FRA A TIL Å
 OVERFLATE FRA A TIL Å VEILEDER FOR FORELDRE MED BARN I 5. 7. KLASSE EMNER Side 1 Innledning til overflate... 2 2 Grunnleggende om overflate.. 2 3 Overflate til:.. 3 3 3a Kube. 3 3b Rett Prisme... 5 3c
OVERFLATE FRA A TIL Å VEILEDER FOR FORELDRE MED BARN I 5. 7. KLASSE EMNER Side 1 Innledning til overflate... 2 2 Grunnleggende om overflate.. 2 3 Overflate til:.. 3 3 3a Kube. 3 3b Rett Prisme... 5 3c
TMA4240 Statistikk H2010 (20)
") TMA4240 Statistikk H2010 (20) 10.5: Ett normalfordelt utvalg, kjent varians (repetisjon) 10.4: P-verdi 10.6: Konfidensintervall vs. hypotesetest 10.7: Ett normalfordelt utvalg, ukjent varians Mette Langaas
TMA4240 Statistikk H2010 (20) 10.5: Ett normalfordelt utvalg, kjent varians (repetisjon) 10.4: P-verdi 10.6: Konfidensintervall vs. hypotesetest 10.7: Ett normalfordelt utvalg, ukjent varians Mette Langaas
PSY2012 Forskningsmetodologi III: Statistisk analyse, design og måling Eksamen vår 2014
 Psykologisk institutt PSY2012 Forskningsmetodologi III: Statistisk analyse, design og måling Eksamen vår 2014 Skriftlig skoleeksamen fredag 2. mai, 09:00 (4 timer). Kalkulator uten grafisk display og tekstlagringsfunksjon
Psykologisk institutt PSY2012 Forskningsmetodologi III: Statistisk analyse, design og måling Eksamen vår 2014 Skriftlig skoleeksamen fredag 2. mai, 09:00 (4 timer). Kalkulator uten grafisk display og tekstlagringsfunksjon
Tillatte hjelpemidler: C3: alle typer kalkulator, alle andre hjelpemidler
 EKSAMENSOPPGAVER Institutt: Eksamen i: Tid: IKBM STAT100 Torsdag 13.des 2012 STATISTIKK 09.00-12.30 (3.5 timer) Emneansvarlig: Solve Sæbø ( 90065281) Tillatte hjelpemidler: C3: alle typer kalkulator, alle
EKSAMENSOPPGAVER Institutt: Eksamen i: Tid: IKBM STAT100 Torsdag 13.des 2012 STATISTIKK 09.00-12.30 (3.5 timer) Emneansvarlig: Solve Sæbø ( 90065281) Tillatte hjelpemidler: C3: alle typer kalkulator, alle
Dobbel og enkel Guyot.
 Dobbel og enkel Guyot. Guyotsystemet, særlig enkel Guyot, er mye brukt i Mellom- Europa, og det er også godt egnet for dyrking på åpen mark i Norge. For å få fullmodne druer er det viktig at en velger
Dobbel og enkel Guyot. Guyotsystemet, særlig enkel Guyot, er mye brukt i Mellom- Europa, og det er også godt egnet for dyrking på åpen mark i Norge. For å få fullmodne druer er det viktig at en velger
ST0202 Statistikk for samfunnsvitere
 ST0202 Statistikk for samfunnsvitere Bo Lindqvist Institutt for matematiske fag 2 Statistisk inferens (kap. 8) Statistisk inferens er å tolke/analysere resultater fra utvalget for å finne ut mest mulig
ST0202 Statistikk for samfunnsvitere Bo Lindqvist Institutt for matematiske fag 2 Statistisk inferens (kap. 8) Statistisk inferens er å tolke/analysere resultater fra utvalget for å finne ut mest mulig
EKSAMEN ST0202 STATISTIKK FOR SAMFUNNSVITERE
 Norges teknisk naturvitenskapelige universitet Institutt for matematiske fag Side 1 av 7 Bokmål Faglig kontakt under eksamen: Bo Lindqvist, tlf. 97589418 EKSAMEN ST00 STATISTIKK FOR SAMFUNNSVITERE Torsdag
Norges teknisk naturvitenskapelige universitet Institutt for matematiske fag Side 1 av 7 Bokmål Faglig kontakt under eksamen: Bo Lindqvist, tlf. 97589418 EKSAMEN ST00 STATISTIKK FOR SAMFUNNSVITERE Torsdag
Inferens i regresjon
 Strategi som er fulgt hittil: Inferens i regresjon Deskriptiv analyse og dataanalyse først. Analyse av en variabel før studie av samvariasjon. Emne for dette kapittel er inferens når det er en respons
Strategi som er fulgt hittil: Inferens i regresjon Deskriptiv analyse og dataanalyse først. Analyse av en variabel før studie av samvariasjon. Emne for dette kapittel er inferens når det er en respons
Denne uken: kap : Introduksjon til statistisk inferens. - Konfidensintervall - Hypotesetesting - P-verdier - Statistisk signifikans
 Denne uken: kap. 6.1-6.2-6.3: Introduksjon til statistisk inferens - Konfidensintervall - Hypotesetesting - P-verdier - Statistisk signifikans VG 25/9 2011 Statistisk inferens Mål: Trekke konklusjoner
Denne uken: kap. 6.1-6.2-6.3: Introduksjon til statistisk inferens - Konfidensintervall - Hypotesetesting - P-verdier - Statistisk signifikans VG 25/9 2011 Statistisk inferens Mål: Trekke konklusjoner
TMA4245 Statistikk Eksamen august 2014
 TMA4245 Statistikk Eksamen august 2014 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Oppgave 1 En bedrift produserer en type medisin i pulverform Medisinen selges på flasker
TMA4245 Statistikk Eksamen august 2014 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Oppgave 1 En bedrift produserer en type medisin i pulverform Medisinen selges på flasker
ST0202 Statistikk for samfunnsvitere
 ST0202 Statistikk for samfunnsvitere Bo Lindqvist Institutt for matematiske fag 2 Kap. 10: Inferens om to populasjoner Situasjon: Vi ønsker å sammenligne to populasjoner med populasjonsgjennomsnitt henholdsvis
ST0202 Statistikk for samfunnsvitere Bo Lindqvist Institutt for matematiske fag 2 Kap. 10: Inferens om to populasjoner Situasjon: Vi ønsker å sammenligne to populasjoner med populasjonsgjennomsnitt henholdsvis
Løsningsforsalg til andre sett med obligatoriske oppgaver i STK1110 høsten 2015
 Løsningsforsalg til andre sett med obligatoriske oppgaver i STK1110 høsten 2015 R-kode for alle oppgaver er gitt bakerst. Oppgave 1 (a) Boksplottet antyder at verdiene er høyere for kvinner enn for menn.
Løsningsforsalg til andre sett med obligatoriske oppgaver i STK1110 høsten 2015 R-kode for alle oppgaver er gitt bakerst. Oppgave 1 (a) Boksplottet antyder at verdiene er høyere for kvinner enn for menn.
Undersøkelser av ferskvannsrødalger i Norge
 B LYTTIA NORGES BOTANISKE ANNALER Undersøkelser av ferskvannsrødalger i Norge Eli-Anne Lindstrøm og Jan Rueness Lindstrøm, E.-A. & Rueness, J. 2009. Undersøkelser av ferskvannsrødalger i Norge. Blyttia
B LYTTIA NORGES BOTANISKE ANNALER Undersøkelser av ferskvannsrødalger i Norge Eli-Anne Lindstrøm og Jan Rueness Lindstrøm, E.-A. & Rueness, J. 2009. Undersøkelser av ferskvannsrødalger i Norge. Blyttia
Krysstabellanalyse (forts.) SOS1120 Kvantitativ metode. 4. Statistisk generalisering. Forelesningsnotater 9. forelesning høsten 2005.
 SOS1120 Kvantitativ metode. 4. Statistisk generalisering. Forelesningsnotater 9. forelesning høsten 2005.") SOS112 Kvantitativ metode Krysstabellanalyse (forts.) Forelesningsnotater 9. forelesning høsten 25 4. Statistisk generalisering Per Arne Tufte Eksempel: Hypoteser Eksempel: observerte frekvenser (O) Hvordan
SOS112 Kvantitativ metode Krysstabellanalyse (forts.) Forelesningsnotater 9. forelesning høsten 25 4. Statistisk generalisering Per Arne Tufte Eksempel: Hypoteser Eksempel: observerte frekvenser (O) Hvordan
6. kurskveld Ila, 7. juni - 06 Statistikk og sannsynlighet
 . kurskveld Ila, 7. juni - 0 Statistikk og sannsynlighet Sannsynlighet og kombinatorikk Sannsynlighet er noe vi omgir oss med nesten daglig. Vi spiller Lotto og andre spill, og håper vi har flaks og vinner.
. kurskveld Ila, 7. juni - 0 Statistikk og sannsynlighet Sannsynlighet og kombinatorikk Sannsynlighet er noe vi omgir oss med nesten daglig. Vi spiller Lotto og andre spill, og håper vi har flaks og vinner.
TMA4240 Statistikk 2014
 TMA4240 Statistikk 2014 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Øving nummer 12, blokk II Oppgave 1 På ein av vegane inn til Trondheim er UP interessert i å måle effekten
TMA4240 Statistikk 2014 Norges teknisk-naturvitenskapelige universitet Institutt for matematiske fag Øving nummer 12, blokk II Oppgave 1 På ein av vegane inn til Trondheim er UP interessert i å måle effekten
1 Sec 3-2: Hvordan beskrive senteret i dataene. 2 Sec 3-3: Hvordan beskrive spredningen i dataene
 1 Sec 3-2: Hvordan beskrive senteret i dataene 2 Sec 3-3: Hvordan beskrive spredningen i dataene Todeling av statistikk Deskriptiv statistikk Oppsummering og beskrivelse av den stikkprøven du har. Statistisk
1 Sec 3-2: Hvordan beskrive senteret i dataene 2 Sec 3-3: Hvordan beskrive spredningen i dataene Todeling av statistikk Deskriptiv statistikk Oppsummering og beskrivelse av den stikkprøven du har. Statistisk
FORSLAG TIL AKTIVITETER
 FORSLAG TIL AKTIVITETER Når vi samler inn materiale, dvs. planter og dyr, fra ferskvann må vi oppbevare dem i det vannet vi henter dem fra, for eksempel i bøtter eller plastbakker. Skal etterarbeidet gjøres
FORSLAG TIL AKTIVITETER Når vi samler inn materiale, dvs. planter og dyr, fra ferskvann må vi oppbevare dem i det vannet vi henter dem fra, for eksempel i bøtter eller plastbakker. Skal etterarbeidet gjøres