(12) Oversettelse av europeisk patentskrift
|
|
|
- Kaj Mons Ellingsen
- 7 år siden
- Visninger:
Transkript
1 (12) Oversettelse av europeisk patentskrift (11) NO/EP B1 (19) NO NORGE (1) Int Cl. C07K 16/38 (06.01) A61K 39/39 (06.01) A61P 7/04 (06.01) Patentstyret (21) Oversettelse publisert (80) Dato for Den Europeiske Patentmyndighets publisering av det meddelte patentet (86) Europeisk søknadsnr (86) Europeisk innleveringsdag (87) Den europeiske søknadens Publiseringsdato () Prioritet , EP, (84) Utpekte stater AT BE BG CH CY CZ DE DK EE ES FI FR GB GR HR HU IE IS IT LI LT LU LV MC MK MT NL NO PL PT RO SE SI SK SM TR (73) Innehaver Novo Nordisk A/S, Novo Allé, 2880 Bagsværd, DK-Danmark (72) Oppfinner HILDEN, Ida, Grøndals Parkvej 8, DK-27 Vanløse, DK-Danmark KROGH, Berit Olsen, Vamdrupvej 7, DK-26 Rødovre, DK-Danmark CLAUSEN, Jes Thorn, Davrekildevej, DK-4270 Hoeng, DK-Danmark OLSEN, Ole Hvilsted, Bækkeskovvej 381., DK-2700 Brønshøj, DK-Danmark BREINHOLT, Jens, Aftenbakken 18, DK-2870 Dyssegård, DK-Danmark LAURITZEN, Brian, Kagsåbrinken 23, DK-27 Herlev, DK-Danmark SØRENSEN, Brit Binow, Marianelundsvej 1, DK-3460 Birkerød, DK-Danmark (74) Fullmektig Onsagers AS, Postboks 1813 Vika, 0123 OSLO, Norge (4) Benevnelse Antistoffer mot vevsfaktorsignalveisinhibitor (6) Anførte publikasjoner WO-A-92/0784 R&D Systems: "R&D Systems New Products - June 07" Internet 1 June 07 ( ), page 8, XP Retrieved from the Internet: URL: /934.pdf> [retrieved on -02-] ENGELMANN BERND: "Novel initiation mechanism of blood coagulation by intravascular tissue factor" BLOOD, AMERICAN SOCIETY OF HEMATOLOGY, US, [Online] vol. 4, no. 11, PART 2, 1 November 04 ( ), page 78B, XP ISSN: Retrieved from the Internet: URL: /cgi/content/abstract/4/11/3967> [retrieved on ] ERHARDTSEN E ET AL: "Blocking of tissue factor pathway inhibitor (TFPI) shortens the bleeding time in rabbits with antibody induced haemophilia A." BLOOD COAGULATION & FIBRINOLYSIS : AN INTERNATIONAL JOURNAL IN HAEMOSTASIS AND THROMBOSIS JUL 199, vol. 6, no., July 199 (199-07), pages , XP ISSN: BRINKMANN T ET AL: "Synthesis of tissue factor pathway inhibitor in human synovial cells and chondrocytes makes joints the predilected site of bleeding in haemophiliacs." EUROPEAN JOURNAL OF CLINICAL CHEMISTRY AND CLINICAL BIOCHEMISTRY : JOURNAL OF THE FORUM OF EUROPEAN CLINICAL CHEMISTRY SOCIETIES APR 1994, vol. 32, no. 4, April 1994 ( ), pages , XP ISSN: WELSCH D J ET AL: "Effect of lipoprotein-associated coagulation inhibitor (LACI) on thromboplastin-induced coagulation of normal and hemophiliac plasmas" THROMBOSIS
2 RESEARCH, TARRYTOWN, NY, US, vol. 64, no. 2, 1 October 1991 ( ), pages , XP ISSN: [retrieved on ] BURGERING M J M ET AL: "The second Kunitz domain of human tissue factor pathway inhibitor: cloning, structure determination and interaction with factor Xa", JOURNAL OF MOLECULAR BIOLOGY, ACADEMIC PRESS, UNITED KINGDOM, vol. 269, no. 3, 13 June 1997 ( ), pages , XP004439, ISSN: , DOI:.06/JMBI
3 1 Oppfinnelsen område Foreliggende oppfinnelse angår antistoffer som spesifikt binder til vevsfaktorsignalveisinhibitor (TFPI, «tissue factor pathway inhibitor»). 1 Bakgrunn for oppfinnelsen Hos subjekter med koagulopati, som for eksempel hos mennesker med hemofili A og B, er forskjellige trinn i koagulasjonskaskaden dysfunksjonelle på grunn av, for eksempel, fravær, eller utilstrekkelig tilstedeværelse, av en koagulasjonsfaktor. Slik dysfunksjon av én del av koagulasjonskaskaden fører til insuffisient blodkoagulasjon og potensielt livstruende blødning, eller skade på indre organer, som for eksempel leddene. Subjekter slik som mennesker med hemofili A og B kan motta erstatningsterapi av koagulasjonsfaktor slik som eksogen FVIIIa eller FIXa, henholdsvis. Imidlertid er slike pasienter med risiko for å utvikle "inhibitorer" (antistoffer) mot slike eksogene faktorer, hvilket gjør tidligere effektiv behandling ineffektiv. Videre kan eksogene koagulasjons-faktorer kun administreres intravenøst, hvilket er meget besværlig og ubehagelig for pasientene. For eksempel kan det være nødvendig med kirurgisk innlegging av intravenøse katetere i en brystvene hos spedbarn og småbarn, for å sikre venetilgang. Dette medfører stor risiko for å utvikle bakterielle infeksjoner. Subjekter med koagulopati kan kun motta behandling etter at en blødning har startet, heller enn som et forebyggende tiltak, hvilket ofte har innvirkning på deres generelle livskvalitet. Det er følgelig fortsatt mange uoppfylte medisinske behov angående hemofili, spesielt, og hos subjekter med koagulopatier, generelt. 2 3 Når en årevegg blir skadet, blir vevsfaktor (TF) eksponert for innholdet i sirkulerende blod og TF danner et kompleks med Faktor VII/aktivert Faktor VII (FVII/FVIIa) på overflaten av TF-bærende celler. Dette fører til aktivering av Faktor X (FX) til FXa som sammen med FVa danner en begrenset mengde av trombin (FIIa). Små mengder av trombin aktiverer blodplater, hvilket fører til overflateeksponering av fosfolipider som støtter binding av tenasekomplekset bestående av FVIIIa/FIXa. Tenasekomplekset produserer store mengder FXa, som deretter fremmer et fullstendig utbrudd av trombin. Et fullstendig trombin-utbrudd er nødvendig for dannelse av en mekanisk sterk fibrinstruktur og stabilisering av den hemostatiske pluggen. FVIII eller FIX mangler eller er tilstede i svært lave nivåer hos hemofilipasienter, og på grunn av mangel på tenaseaktivitet, er evnen til å danne FXa lav og ikke tilstrekkelig til å støtte propageringsfasen av koagulasjonen. I motsetning til dette er den TF-medierte initieringsfasen ikke avhengig av dannelse
4 2 1 2 av tenasekompleks. Imidlertid vil TF-reaksjonsveien kort tid etter en initiell FXagenerering, bli blokkert av plasma inhibitorer. Vevsfaktorsignalveisinhibitor (TFPI) nedregulerer pågående koagulasjon ved å nøytralisere den katalytiske aktiviteten av FXa og ved å inhibere TF-FVIIakomplekset i nærvær av FXa. TFPI inhiberer enten TF/FVIIa/FXa-komplekset på celleoverflaten eller inhiberer frigjort FXa fulgt av FVIIa/TF-inhibering. Int. Jour. Haem. Fibrin. (199), 6,, beskriver et polyklonalt geit TFPI antistoff. Etter publisering av denne artikkelen anvendte Warn Cramer og Rapaport den samme metoden som den beskrevet for å frembringe et geit pab. (Arterioscler. Thromb. (1993), 13, 11-17). Dette pab ble testet og funnet å oppvise den uønskede egenskapen med å føre til en reduksjon i blodplatetallet (se Fig. 4). Blood; Amer. Soc. Haem. (04), 4, 11, 708 beskriver et TFPI antistoff målrettet mot K2-domenet av TFPI (se linje i sammendraget). Iingen ytterligere informasjon er gitt om antistoffet. Throm. Res. (1991), 64, 2, er en artikkel som angår den effekten som LACI (f.eks. TFPI) har på den tromboplastin-induserte koagulasjonen av normalt og hemofilt plasma. Artikkelen beskriver et kanin polyklonalt TFPI antistoff (pab) og et mus monoklonalt TFPI antistoff (mab) (se avsnittet "Materials", side 21), anvendt for å vurdere LACI-funksjon. Artikkelen beskriver ikke noe angående strukturen av antistoffene som ble benyttet for å vurdere LACI-funksjon., den undersøker heller ikke de biologiske egenskapene av pab og mab som ble anvendt i forfatterens forsøk. mab 2974 er et kommersielt tilgjengelig monoklonalt TFPI antistoff, fremstilt av R&D Systems og tilgjengelig kun for forskningsformål. 3 Oppsummering av oppfinnelsen Oppfinnerne har identifisert monoklonale antistoffer som spesifikt binder til vevsfaktorsignalveisinhibitor ("TFPI", noen ganger referert til som "TFPI1") og derved modulerer dens aktivitet. Foreliggende oppfinnelse angår disse antistoffene og andre relaterte antistoffer som er avledet fra disse antistoffene eller har lignende bindingsegenskaper som disse antistoffene. Følgelig angår foreliggende oppfinnelse antistoffer som binder spesifikt til vevsfaktorsignalveisinhibitor (TFPI) og som reduserer koagulasjonstid i, for eksempel, (a) humant plasma med FVIII-mangel og/eller (b) humant helblod. Foreliggende oppfinnelse angår et monoklonalt antistoff som er i stand til å binde spesifikt til K2-domenet av TFPI, hvori nevnte antistoff er i stand til å binde
5 3 1 spesifikt til en epitop omfattende rest R17 av SEQ ID NO: 2 og KD av nevnte antistoff er mindre enn 0,8 nm, som bestemt ved anvendelse av overflate plasmonresonans. Ett antistoff omfatter lett kjede variabel region av SEQ ID NO: 4 og tung kjede variabel region av SEQ ID NO: 8. Et annet antistoff omfatter lett kjede variabel region av SEQ ID NO: 1 og tung kjede variabel region av SEQ ID NO: 18. Oppfinnelsen tilveiebringer også polynukleotider som koder for et antistoff ifølge oppfinnelsen, slik som polynukleotider som koder for en lett kjede av et antistoff og/eller en tung kjede av et antistoff ifølge oppfinnelsen. Oppfinnelsen tilveiebringer også farmasøytiske preparater omfattende et antistoff eller polynukleotid ifølge oppfinnelsen og en farmasøytisk akseptabel bærer eller fortynningsmiddel. Antistoffene, polynukleotidene og preparatene ifølge oppfinnelsen tilveiebringes også for anvendelse ved (a) behandling eller forebygging av en koagulopati (blødningsforstyrrelse) eller (b) stimulering av blodkoagulasjon. Følgelig tilveiebringer oppfinnelsen en fremgangsmåte for (a) behandling eller forebygging av en koagulopati (blødningsforstyrrelse) eller (b) stimulering av blodkoagulasjon, hvor fremgangsmåten omfatter å administrere til en pasient med behov derfor en terapeutisk eller profylaktisk effektiv mengde av et antistoff, polynukleotid eller preparat ifølge oppfinnelsen. Videre tilveiebringer oppfinnelsen doseringsregimer av nevnte monoklonale antistoff ifølge oppfinnelsen. 2 3 Kort beskrivelse av Figurene Figur 1 viser sekvensene av VH (A) og VL (B)-domenene fra mus anti- TFPI4F36A1B2 (heri også referrert til som MuTFPI4F36 eller 4F36), sammenstilt med sekvensene for human kjønnscelle og initiell CDR-graftet versjon av humanisert TFPI4F36. Kabat nummereringsskjema er angitt over sekvensene. Figur 2 viser nukleotidsekvensene og de translaterte polypeptidsekvensene for VH og VL-sekvensene for det murine antistoffet TFPI4F36A1B2 (MuTFPI4F36). Figur 3 viser aminosyresekvensene for lett (A) og tung (B) kjede av Fabfragmenter av det murine 4F36-antistoffet, MuTFPI4F36. Nummerering over sekvensene er vist i henhold til Kabat. Posisjoner som svarer til CDR-løkker er fremhevet i understreket fet skrift i Kabat-nummereringen. Aminosyrerester som utgjør paratopen er fremhevet i understreket fet skrift. Paratopen er bestemt fra røntgenstrukturen av komplekset mellom MuTFPI4F36-Fab og TFPI K2-domenet og er definert som rester i Fab som har et tungt atom innenfor en avstand på mindre enn 4 Å fra et tungt atom i K2. Figur 4 viser sekvensen av TFPI
6 (signalpeptidsekvens utelatt). Kunitz-domenene er vist i fet skrift: TFPI Kunitzdomene 1 = aminosyrene 26 til 76; TFPI Kunitz-domene 2 = aminosyrene ; TFPI Kunitz-domene 3 = aminosyrene Den C-terminale delen av TFPI er vist i kursiv ved aminosyrene 240 til 276. Figur viser den relative tilgjengelighet av restene i TFPI. Restene som har en tilgjengelighet høyere enn 40 % er aminosyrene 94-9, 98, 0-1, , , 131, 134, og Figur 6 viser en SEC HPLC-analyse av et kompleks mellom TFPI Kunitzdomene 2 (K2) og MuTFPI4F36 Fab-fragmentet (Fab). SEC-HPLC-kromatogrammer detektert ved UV 280 nm av fritt K2 (hel linje, r t 13,1 min, topp vist ved 13,134), fri Fab (streket linje, r t 11,7 min, topp vist ved 11,676) og kompleks (prikket linje, r t 11, min, topp vist ved 11,496). Prøven av komplekset inneholdt ~ % overskudd av K2. Figur 7 viser den totale strukturen av MuTFPI4F36 Fab:K2-komplekset. Lette kjeder er vist i lys grå og tunge kjeder er vist i mørk grå. CDR-løkkene som definert i henhold til Kabat-skjema er merket som L1 til L3 og H1 til H3. Figur 8 viser strukturen av K2-domenet av TFPI når det er i et kompleks med MuTFPI4F36 Fab (Fab-molekyl ikke vist). De N- og C-terminale endene og sekundærstruktur-elementer er merket. Figur 9 viser en ryggrad ("back-bone") superponering av K2-strukturer. Viser forskjeller i struktur mellom K2 løsning, K2 er i kompleks med MuTFPI4F36 Fab og K2 i kompleks med trypsin fra svin. Figur viser MuTFPI4F36-bindingsepitopen på K2. (A) Tegnet fremstilling av K2-domenet fra TFPI med sidekjeder av rester inkludert i bindingsepitopen representert ved kulepinnemodell. (B) er som A, men med overflate tilføyd. (C) Bindingsepitop kartlagt til primærsekvens. Store bokstaver, bokstaver i fet skrift, kursiv og understrekede bokstaver svarer til rester i K2-bindingsepitopen som kommer i kontakt med MuTFPI4F36 Fab tung kjede kun (posisjonene, 11, 13, 28, 31, 33 og 3), lett kjede kun (posisjonene 21, 23 og 0), og med både tung og lett kjede (17, 19, 34 og 36), henholdsvis. Sekundærstruktur-elementer (h = heliks, s = sheet) er angitt (helikser ved posisjonene -8 og 0-6 og sheets ved posisjonene -26 og 31-37). Rester fremhevet i grått (posisjonene 1-2 og 9-66) er til stede i det uttrykte proteinet, men er ikke observert i krystallstrukturen på grunn av at de N- og C- terminale endene er fleksible. Figur 11 viser en sammenligning av ryggrad linjene for K2: MuTFPI4F36 Fab og 2: HzTFPI4F36 Fab-komplekser, som viser de identiske bindings formene for de murine MuTFPI4F36 og humaniserte HzTFPI4F36 Fab-fragmentene. K2:MuTFPI4F36 Fab er vist i grått og K2:HzTFPI4F36 Fab i svart. Strukturer er superponert for å
7 1 2 3 optimalisere matchen mellom den variable regionen av Fab-fragmentene. Figur 12 viser effekten av anti-tfpi monoklonale antistoffer (mabs) på TF/FVIIa-indusert aktivering av FX på overflaten av HUVECs stimulert med TNFα/IL1ß. Aktivering av FX ble målt i nærvær av 0- nm mab (mabtfpi 21 eller mab 2974), 0 pm FVIIa (NovoSeven ) og 0 nm FX i buffer med 2 mm HEPES, 137 mm NaCl, 3, mm KCl, mm CaCl 2, 1 mg/ml BSA (0,1 %) ph 7,4 som ble dekket med et monolag av HUVECs. Generert FXa-aktivitet ble bestemt i en amidolytisk assay med S-276 målt ved økningen i absorbans ved 40 nm. Figur 13 viser effekten av anti-tfpi mabs på TFPI-inhibering av TF/FVIIaindusert aktivering av FX på overflaten av MDA-MB 231-celler. Aktivering av FX ble målt i nærvær av 0- nm mab (Hz mabtfpi 21 eller mab 2974), 2, nm fl-tfpi, 0 pm FVIIa og 0 nm FX i buffer med 2 mm HEPES, 137 mm NaCl, 3, mm KCl, mm CaCl, 1 mg/ml BSA (0,1 %) ph 7,4 som ble dekket med monolag av MDA-MB 231-celler. Generert FXa-aktivitet ble bestemt i en amidolytisk assay med S-276 målt ved økningen i absorbans ved 40 nm. Figur 14 viser effekten av enkelt aminosyre alanin-substitusjoner av valgte rester innenfor TFPI Kunitz 2-domenet på binding til mabtfpi 21 ("mab4f36") og mab2974 (n = 2). De valgte restene er del av den mabtfpi 21 bindende epitopen. Nummereringen av aminosyrerester er angitt i fig. C. Figur 1 viser blødningstiden i huden og blodtap målt i transient hemofile kaniner etter behandling med kontroll IgG (Hemofili) eller med det murine anti-tfpiantistoffet, TFPI-4F36A1B2 ("4F36", MuTFPI4F36). Figur 16 viser blødningstiden i huden (enkelt observasjoner; middelverdi ± SEM) og blodtap (middelverdi+sem) i en behandling "på forespørsel" ("on demand") av kaniner med antistoff-indusert hemofili, behandlet med HzTFPI4F36 ("anti-tfpi", mabtfpi 21) (2 mg/kg) eller NovoSeven (9 mg/kg) minutter etter induksjon av blødning. Blødningen ble observert i 1 time (3600 sek). Figur 17 viser blødningstiden i huden (enkelt-observasjoner; middelverdi ± SEM) og blodtap (middelverdi+sem) hos kaniner med antistoff-indusert hemofili, når de er behandlet på forhånd med HzTFPI4F36 ("anti-tfpi", mabtfpi 21) (doser: 0,, 1, 2 mg/kg) eller et isotype kontroll-antistoff 3 minutter før induksjon av blødning. Blødningen ble observert i 1 time (3600 sek). Figur 18 viser blodplatetallet målt i individuelle dyr, etter stimulering med anti-fviii antistoff, administrering av et anti-tfpi-antistoff ("anti-tfpi ab", MuTFPI4F36) og deretter fått igang blødning. Dette ble utført i en kontroll hemofilimodell og i nærvær av det murine anti-tfpi-antistoffet 4F36 (MuTFPI4F36) som beskrevet heri. Figur 19 viser plasmakonsentrasjonen av fritt HzTFPI4F36 (mabtfpi 21) i
8 6 1 2 kaniner dosert med mg/kg HzTFPI4F36 ved 0 timer. Hud-blødningsforsøk ble utført ved 96 timer (4 dager), 168 timer (7 dager) og 240 timer ( dager). De prikkede linjene angir det "effektive konsentrasjon"-området for HzTFPI4F36 som funnet i dose-responsstudien (se fig. 17). Figur : Venstre panel : plasma HzTFPI4F36 (mabtfpi 21) (venstre akse: O) og blødningstiden i huden (middelverdi ± SEM; ). Høyre panel : plasma HzTFPI4F36 (mabtfpi 21) (venstre akse O) og blodtap (middelverdi+sem; ) hos kaniner med antistoff-indusert hemofili, når de er behandlet på forhånd med mg/kg HzTFPI4F36 (n=8) eller isotype kontroll-antistoff (n = 12) ved 4, 7 eller dager før induksjon av blødning. Blødningen ble observert i 1 time (3600 sek). Figur 21 viser plasmakonsentrasjonsnivåer etter IV og SC HzTFPI4F36 (mabtfpi 21) administrering til aper. I de tre nedre diagrammene ble to aper administrert tre doser av HzTFPI4F36 med to ukers intervall. Nede til venstre ble tre doser på 2, og 80 mg/kg administrert, nede i midten ble tre doser på, 80 og 160 mg/kg administrert; nede til høyre ble tre doser på 80, 160 og 0 mg/kg dosert. Øverst til venstre ble en enkelt dose på mg/kg administrert til tre aper; øverst til høyre ble enkelt IV dose administrert til tre aper. I diagrammer representerer punktene individuelle observasjoner mens linjen representerer modell tilpasningen. Figur 22 viser en simulering av 1 mg/kg HzTFPI4F36 (mabtfpi 21) administrert SC daglig. Den sammenhengende horisontale linjen representerer simulerte plasmakonsentrasjonsnivåer og den prikkete horisontale linjen den høyeste effektive konsentrasjonen som avledet fra effekt-data. Figur 23 viser en simulering av 1 mg/kg HzTFPI4F36 (mabtfpi 21), administrert intravenøst hver tredje uke. Den sammenhengende horisontale linjen representerer simulerte plasmakonsentrasjonsnivåer og den prikkete horisontale linjen den høyeste effektive konsentrasjonen som avledet fra effekt-data. Figur 24 viser en simulering av mg/kg HzTFPI4F36 (mabtfpi 21), administrert intravenøst annenhver uke. Den sammenhengende horisontale linjen representerer simulerte plasmakonsentrasjonsnivåer og den prikkete horisontale linjen den forventede mål metningen som avledet fra effekt-studien. Kort beskrivelse av sekvenslisten 3 utelatt). SEQ ID NO: 1 viser aminosyresekvensen av human TFPI (signalpeptidsekvens SEQ ID NO: 2 viser aminosyresekvensen for en konstruksjon anvendt for
9 bestemmelse av bindingsepitopen for et antistoff. Konstruksjonen omfatter aminosyrene 91 til fra human TFPI og et C-terminalt HiS 6 -merke. SEQ ID NO: 3, og 4 viser polynukleotid (sense og anti-sense) og polypeptid-sekvensene for lett kjede variabelt domene (VL) av MuTFPI4F36 (TFPI- 4F36A1B2) monoklonalt antistoff. SEQ ID NO: 6 viser aminosyresekvensen av den lette kjeden av MuTFPI4F36 (TFPI-4F36A1B2) monoklonalt antistoff. Signalpeptidsekvenser er utelatt. SEQ ID NO: 7, 9 og 8 viser polynukleotid (sense og anti-sense) og polypeptid-sekvensene for tung kjede variabelt domene (VH) fra MuTFPI4F36 (TFPI- 4F36A1B2) monoklonalt antistoff. SEQ ID NO: viser aminosyresekvensen av den tunge kjeden av det monoklonale antistoffet MuTFPI4F36 (TFPI-4F36A1B2). Signalpeptidsekvenser er utelatt. SEQ ID NO: 11 viser sekvensen av en revers primer anvendt for amplifikasjon av tung kjede variabelt domene og SEQ ID NO: 12 viser sekvensen av en revers primer anvendt for amplifikasjon av lett kjede. SEQ ID NO: 13-1 viser sense polynukleotid-, anti-sense polynukleotid- og polypeptidsekvensene, henholdsvis, for lett kjede variabelt domene (VL) av det humaniserte monoklonale antistoffet, HzTFPI4F36 (mabtfpi21). Signalpeptidsekvenser er utelatt. SEQ ID NO: viser sense polynukleotid-, anti-sense polynukleotid- og polypeptidsekvensene, henholdsvis, for tung kjede variabelt domene (VH) av det humaniserte monoklonale antistoffet, HzTFPI4F36 (mabtfpi21). SEQ ID NO: viser sense polynukleotid, anti-sense polynukleotid og polypeptidsekvensene, henholdsvis, for den lette kjeden (LC) av det humaniserte monoklonale antistoffet, HzTFPI4F36 (mabtfpi21). SEQ ID NO: viser sense polynukleotid, anti-sense polynukleotid og polypeptidsekvensene, henholdsvis, for den tunge kjeden (HC) av det humaniserte monoklonale antistoffet, HzTFPI4F36 (mabtfpi21). Signalpeptid-sekvenser er utelatt. SEQ ID NO: 2-26 viser nukleinsyre- og aminosyresekvenser, henholdsvis, for lett kjede variabelt domene av CDR-graftet HzTFPI4F36. Signalpeptidsekvenser er utelatt. SEQ ID NO: viser nukleinsyre- og aminosyresekvensene, henholdsvis, av tung kjede variabelt domene av CDR-graftet HzTFPI4F36. Signalpeptidsekvenser er utelatt. SEQ ID NO: 29 viser aminosyresekvensen av den lette kjeden av CDR-graftet
10 8 1 HzTFPI4F36 (human kappa-kjede). Signalpeptidsekvensen er utelatt. SEQ ID NO: viser aminosyresekvensen av den tunge kjeden av CDRgraftet HzTFPI4F36, som er en human IgG4 (S241P). Signalpeptidsekvensen er utelatt. SEQ ID NO: 31 viser kjønnscelle-sekvensen, VKII_A18/JK4, anvendt for humanisering av den lette kjeden av MuTFPI4F36. Signalpeptidsekvensen er utelatt. SEQ ID NO: 32 viser kjønnscellesekvensen, VH3_21/JH6, anvendt for humanisering av den tunge kjeden av MuTFPI4F36. Signalpeptidsekvensen er utelatt. SEQ ID NO: 33 viser aminosyresekvensen av MuTFPI4F36A1B2 tung kjede Fab. Signalpeptidet er utelatt. SEQ ID NO: 34 viser aminosyresekvensen for HzTFPI4F36 tung kjede Fab. Signalpeptidet er utelatt. 2 3 Detaljert beskrivelse av oppfinnelsen Foreliggende oppfinnelse angår antistoffer som binder til TFPI. Antistoffene binder foretrukket spesifikt til TFPI, dvs. de binder til TFPI men de binder ikke, eller bind med lavere affinitet, til andre molekyler. Speielt angår oppfinnelsen antistoffer som binder til TFPI og som modulerer dens aktivitet. Antistoffer ifølge oppfinnelsen kan følgelig ha evne til å redusere koagulasjonstid. For eksempel kan et antistoff ifølge oppfinnelsen ha evne til å redusere koagulasjonstid i humant plasma med FVIII-mangel eller til å redusere tid til koagulering som målt i en tromboelastografi (TEG)-analyse av humant helblod. Oppfinnelsen også angår anvendelser for slike antistoffer, slik som terapeutiske og farmasøytiske anvendelser. Betegnelsen TFPI som anvendt heri omfatter hvilken som helst naturlig forekommende form av TFPI som kan avledes fra hvilken som helst passende organisme. For eksempel kan TFPI for anvendelse som beskrevet heri være et mammalsk TFPI, slik som humant, fra mus, rotte, primat, bovint, ovint, eller TFPI fra svin. Foretrukket er TFPI human TFPI. TFPI kan være en moden form av TFPI slik som et TFPI-protein som har gjennomgått posttranslasjonell prosessering i en egnet celle. Et slikt modent TFPI-protein kan, for eksempel, være glykosylert. TFPI kan være et fullengde TFPI-protein. Betegnelsen TFPI omfatter også varianter, isoformer og andre homologer av slike TFPI-molekyler. Variant TFPI-molekyler vil generelt være karakterisert ved at de har samme type aktivitet som naturlig forekommende TFPI, slik som evne til å nøytralisere den katalytiske aktiviteten av FXa, eller evne til
11 å inhibere et kompleks av TF-FVIIa/FXa. Et antistoff ifølge oppfinnelsen vil ha evne til å binde til TFPI. Foretrukket vil et antistoff ifølge oppfinnelsen binde spesifikt til TFPI. Det vil si at et antistoff ifølge oppfinnelsen vil foretrukket binde til TFPI med høyere bindingsaffinitet enn den ved hvilken det binder til et annet molekyl. Et antistoff ifølge oppfinnelsen kan ha evne til å binde eller spesifikt binde til et TFPI-molekyl som beskrevet heri slik som hvilket som helst mål-molekyl som beskrevet heri. Betegnelsen "bindingsaffinitet" er heri anvendt som et mål på styrken av en ikke-kovalent interaksjon mellom to molekyler, f.eks. og antistoff, eller fragment derav, og et antigen. Betegnelsen "bindingsaffinitet" blir anvendt for å beskrive monovalente interaksjoner (intrinsisk aktivitet). Bindingsaffinitet mellom to molekyler, f.eks. et antistoff, eller fragment derav, og et antigen, gjennom en monovalent interaksjon kan kvantifiseres ved bestemmelse av dissosiasjonskonstanten (K D ). I sin tur kan K D bestemmes ved måling av kinetikken av kompleksdannelse og dissosiasjon, f.eks. ved SPR-metoden (Biacore). Hastighets-konstantene som svarer til assosiasjon og dissosiasjon av et monovalent kompleks er referert til som assosiasjonsrate-konstanten k a (eller k on ) og dissosiasjonsrate-konstanten k d. (eller k off ), henholdsvis. K D er relatert til k a og k d gjennom ligningen K D = k d / k a. Ifølge ovennevnte definisjon kan bindingsaffiniteter forbundet med forskjellige molekylære interaksjoner, f.eks. sammenligning av bindingsaffiniteten av forskjellige antistoffer for et gitt antigen, sammenlignes ved å sammenligne K D - verdiene for de individuelle antistoff/antigen-kompleksene. På lignende måte kan spesifisiteten av en interaksjon vurderes ved bestemmelse og sammenligning av K D -verdien for interaksjonen av interesse, f.eks. en spesifikk interaksjon mellom et antistoff og et antigen, med K D -verdien for en interaksjon som ikke er av interesse. Typisk vil K D for antistoffet med hensyn til målet være 2 ganger, foretrukket ganger, mer foretrukket ganger lavere enn K D med hensyn til det andre, ikkemål-molekylet slik som urelatert materiale eller ledsagende materiale i omgivelsene. Mer foretrukket vil K D være 0 ganger lavere, slik som 0 ganger lavere, eller 0 ganger lavere; enda mer foretrukket 00 ganger lavere, slik som 1,000 ganger lavere, eller,000 ganger lavere. Verdien av denne dissosiasjonskonstanten kan bestemmes direkte ved velkjente metoder, og kan beregnes til og med for komplekse blandinger ved metoder slik som de, for eksempel, vist i Caceci et al. (Byte 9: , 1984). For eksempel kan K D fastslås ved anvendelse av en dobbelt-filter nitrocellulosefilter bindingsanalyse slik som den beskrevet av Wong & Lohman (Proc. Natl. Acad. Sci.
12 1 2 3 USA 90, , 1993). Andre standardanalyser for å evaluere bindingsevnen av ligander slik som antistoffer overfor mål er kjent på området, inkludert for eksempel, ELISA, Western blot, RIA og flowcytometrianalyse. Bindingskinetikken og bindingsaffiniteten for antistoffet kan også vurderes ved standardanalyser kjent på området, slik som overflate plasmonresonans (SPR), f.eks. ved anvendelse av et Biacore TM system. En kompetitiv bindingsanalyse kan foretas i hvilken binding av antistoffet til målet blir sammenlignet med binding av målet ved en annen ligand for dette målet, slik som et annet antistoff. Konsentrasjonen ved hvilken 0 % inhibering finner sted er kjent som Ki. Under ideelle betingelser er Ki ekvivalent med K D. Ki-verdien vil aldri være lavere enn K D, slik at måling av Ki hensiktsmessig kan anvendes i stedet, for å gi en øvre grense for K D. Et antistoff ifølge oppfinnelsen kan ha en K D for dets mål på 1 x -7 M eller lavere, 1 x -8 M eller lavere, eller 1 x -9 M eller lavere, eller 1 x - M eller lavere, 1 x -11 M eller lavere, eller 1 x -12 M eller lavere. Et antistoff som spesifikt binder til dets mål kan binde dets mål med høy affinitet, det vil si at det framviser en lav K D som diskutert ovenfor, og kan binde til andre ikke-mål molekyler med en lavere affinitet. For eksempel kan antistoffet binde til ikke-mål molekyler med en K D på 1 x -6 M eller høyere, mer foretrukket 1 x - M eller høyere, mer foretrukket 1 x -4 M eller høyere, mer foretrukket 1 x -3 M eller høyere, enda mer foretrukket 1 x -2 M eller høyere. Et antistoff ifølge oppfinnelsen er foretrukket i stand til å binde til dets mål med en affinitet som er minst to ganger, ganger, 0 ganger, 0 ganger 0 ganger, 00 ganger, 1,000 ganger eller,000 ganger eller høyere enn dets affinitet for binding til et annet ikke-mål molekyl. Mål-molekylet kan være hvilket som helst TFPI-molekyl som beskrevet heri, slik som et naturlig forekommende TFPI-molekyl, et fullstendig modent TFPI molekyl eller et fullengde TFPI-molekyl. Foretrukne TFPI-molekyler er fullstendig modne, naturlig forekommende, fullengde mammalske TFPI-molekyler. For eksempel kan TFPI-molekylet bestå av, eller kan omfatte, aminosyresekvensen ifølge SEQ ID NO: 1 eller et fragment eller annen variant derav som beskrevet heri. Mål-molekylet kan være en variant av et TFPI-molekyl slik som et fragment av et TFPI-molekyl. For eksempel kan mål-molekylet være et fragment eller annen variant av TFPI som bevarer en egnet epitop for antistoffbinding. For eksempel kan mål-molekylet være et fragment eller annen variant av TFPI som bibeholder en epitop som beskrevet heri. Mål-molekylet kan omfatte en slik epitop. I én utførelsesform er mål-molekylet et fullengde TFPI-molekyl. Fullengde
13 TFPI-molekylet kan omfatte et første, andre og tredje Kunitz-domene som beskrevet heri. Fullengde TFPI-molekylet kan omfatte et første, andre og tredje Kunitz-domene som beskrevet heri og også en karboksy-terminal region som beskrevet heri. Fullengde TFPI-molekylet kan være et naturlig forekommende TFPI-molekyl slik som et fullengde TFPI polypeptid som uttrykt fra et TFPI-gen, eller som sekretert av TFPIuttrykkende celler. Fullengde TFPI-molekylet kan være et naturlig forekommende TFPI-molekyl som funnet sirkulerende i fri form i plasma eller bundet til celler slik som endotelceller. Fullengde TFPI-molekylet er ikke et trunkert TFPI-molekyl som for eksempel et naturlig forekommende trunkert TFPI-molekyl som beskrevet heri. I én utførelsesform er mål-molekylet et trunkert TFPI-molekyl. For eksempel kan det trunkerte TFPI-molekylet omfatte en karboksy-terminal trunkering. For eksempel er flere naturlig forekommende trunkerte former av TFPI kjent. Disse kan omfatte en trunkering av del av eller hele den karboksyterminale delen av TFPI. De kan videre omfatte trunkering av del av eller hele av ett eller flere av Kunitzdomenene. For eksempel kan en trunkert form av TFPI omfatte delesjon av den karboksy-terminale delen og del av, eller hele, det tredje Kunitz-domenet. For eksempel omfatter én naturlig forekommende trunkert form av TFPI kun aminosyrene 1 til 161 av det fullengde TFPI-molekylet (referert til heri som TFPI (1-161)). TFPI (1-161) er en aktiv form av TFPI som har redusert aktivitet sammenlignet med det fullengde molekylet. TFPI (1-161) avviker i struktur fra fullengde TFPI og antistoffer frembrakt mot TFPI (1-161) som et mål-molekyl kan derfor være forskjellig fra antistoffer frembrakt mot fullengde TFPI. En trunkert form av TFPI kan være et formålstjenlig mål-molekyl når det er ønsket å målrette antistoffer mot den regionen av fullengde TFPI som er til stede i TFPI (1-161). Imidlertid blir trunkert TFPI foretrukket anvendt som et mål-molekyl når det er ønsket å rette antistoffer mot spesifikke trunkerte former avtfpi slik som naturlig forekommende trunkert TFPI. I én utførelsesform er mål-molekylet en naturlig forekommende form av TFPI. Denne kan anvendes i en form i hvilken den er til stede in vivo. For eksempel kan mål-molekylet være et fullengde naturlig forekommende TFPI som diskutert ovenfor. Mål-molekylet kan være et trunkert naturlig forekommende TFPI som diskutert ovenfor. Mål-molekylet kan være TFPI i en form i hvilken den er til stede i plasma in vivo. Mål-molekylet kan være TFPI som er bundet til lipoprotein på samme måte som det er til stede i plasma in vivo. Mål-molekylet kan være TFPI som er bundet til celler på samme måte som det forekommer in vivo, slik som TFPI som er bundet til endotelceller. Et antistoff ifølge oppfinnelsen kan binde til en enkelt eller flere av disse naturlig forekommende formene av TFPI. Antistoffet ifølge oppfinnelsen kan være i stand til å binde til alle disse naturlig forekommende formene av TFPI, eller
14 kan være i stand til å skjelne mellom disse ulike formene, og binde til noen men ikke til andre. I én utførelsesform er mål-molekylet, eller omfatter, det andre Kunitzdomenet av TFPI. Et slikt mål-molekyl kan omfatte aminosyrene 97 til 147 av SEQ ID NO: 1 eller aminosyrene 91 til av SEQ ID NO: 1 eller en ekvivalent Kunitzdomene 2-region fra et annet TFPI polypeptid. Et slikt mål-molekyl kan omfatte SEQ ID NO: 2 eller aminosyrene 3 til 8 eller til 0 av SEQ ID NO: 2. Mål-molekylet kan være, eller kan omfatte, et fragment av det andre Kunitz-domenet av TFPI. For eksempel kan mål-molekylet omfatte fem eller flere, åtte eller flere, ti eller flere, tolv eller flere eller femten eller flere aminosyrer fra det andre Kunitz-domenet. Mål-molekylet kan omfatte fem eller flere, åtte eller flere, ti eller flere, tolv eller flere eller femten eller flere overflate-tilgjengelige rester av TFPI eller av en bestemt region av TFPI slik som et bestemt Kunitz-domene eller den C-terminale delen av TFPI. En overflate-tilgjengelig rest er en rest som har mer enn 40 % relativ tilgjengelighet. For eksempel har for Kunitz 2-domenet fra TFPI (SEQ ID NO: 1), følgende aminosyrer mer enn 40 % relativ tilgjengelighet: 94-9, 98, 0-1, , , 131, 134, og (se Figur ). Mål-molekylet kan omfatte fem eller flere, åtte eller flere, ti eller flere, tolv eller flere eller femten eller flere av disse restene, slik som et fragment av TFPI som omfatter fem eller flere, åtte eller flere, ti eller flere, tolv eller flere eller femten eller flere av disse restene. Mål-molekylet kan omfatte en kjent epitop fra TFPI. Betegnelsen "epitop", som anvendt heri, er definert i sammenheng med en molekylær interaksjon mellom et "antigenbindende polypeptid" (Ab) og dets korresponderende "antigen" (Ag). Som anvendt heri omfatter betegnelsen Ab et antistoff eller et fragment derav, som spesifikt binder det korresponderende Ag. Eksempler på antigenbindende fragmenter omfatter Fab, Fab', F(ab)2, F(ab')2, F(ab)S, Fv (typisk VL og VH-domenene fra en enkelt arm av et antistoff), enkeltkjede Fv (scfv; se f.eks.. Bird et al., Science 1988; 242:42S-426; og Huston et al. PNAS 1988; 8: ), dsfv, Fd (typisk VH og CHI domenet), og dab (typisk et VH domene)-fragmenter; VH, VL, VhH og V-NAR-domenene; monovalente molekyler omfattende en enkelt VH og en enkelt VL-kjede; minibodier, diabodier, triabodier, tetrabodier og kappa bodier (se, f.eks., Ill et al.. Protein Eng 1997; :949-7); kamel IgG; IgNAR; så vel som én eller flere isolerte CDR'er eller en funksjonell paratop, hvor de isolerte CDR'ene eller antigenbindende restene eller polypeptidene kan være forbundet eller koblet sammen for å danne et funksjonelt antistoffragment. Forskjellige typer antistoffragmenter har blitt beskrevet eller gjennomgått i, f.eks., Holliger og Hudson, Nat Biotechnol 0;2S: ; WO , og publiserte U.S. Patentsøknader og
15 Antistoffragmenter kan oppnås ved anvendelse av konvensjonelle rekombinante eller proteinkonstruksjons-teknikker, og fragmentene kan screenes for antigenbinding eller annen funksjon på samme måte som intakte antistoffer. Betegnelsen antigen (Ag) angir den molekylære enheten anvendt for immunisering av et immunkompetent virveldyr for å produsere antistoffet (Ab) som gjenkjenner Ag. Heri er Ag betegnet mer generelt og er generelt ment å omfatte mål-molekyler som blir spesifikt gjenkjent av Ab, hvilket følgelig inkluderer fragmenter eller etterligninger av molekylet anvendt i immuniseringsprosessen for å frembringe Ab. Følgelig blir, for Ab'er som binder til det andre Kunitz-domenet (K2) av TFPI, både isolert K2, fullengde TFPI inkludert trunkerte og andre varianter av TFPI referert til som et Ag. Generelt angir betegnelsen "epitop" området eller regionen av et Ag til hvilket et Ab binder spesifikt, dvs. området eller regionen i fysisk kontakt med Ab. En proteinepitop kan omfatte aminosyrerester i Ag som er direkte involvert i binding til et Ab (også kalt den immundominante komponenten av epitopen) og andre aminosyrerester, som ikke er direkte involvert i bindingen, slik som aminosyrerester av Ag som blir effektivt blokkert av Ab (med andre ord er aminosyreresten innenfor den "løsningsmiddel-ekskludert overflaten" og/eller "footprint" av Ab). Betegnelsen epitop heri omfatter begge typer bindingsseter i hvilken som helst bestemt region av K2 i TFPI som spesifikt binder til et anti-tfpi antistoff, eller et annet K2-spesifikt middel ifølge oppfinnelsen, dersom ikke annerledes angitt (f.eks., i noen sammenhenger angår oppfinnelsen antistoffer som binder direkte til bestemte aminosyrerester). K2 kan omfatte flere forskjellige epitoper, som kan omfatte, uten begrensning, (1) lineært peptid antigendeterminanter, (2) konformasjonelle antigene determinanter som består av én eller flere ikke-tilgrensende aminosyrer lokalisert nær hverandre i den modne K2-konformasjonen; og (3) posttranslasjonelle antigene determinanter som, enten i sin helhet eller delvis, består av molekylære strukturer kovalent bundet til K2, slik som karbohydratgrupper. Epitopen for et gitt antistoff (Ab)/antigen (Ag)-par kan defineres og karakteriseres på forskjellig detaljnivå ved anvendelse av mange forskjellige eksperimentelle og data-metoder for epitopkartlegging. De eksperimentelle metodene omfatter mutagenese, røntgenkrystallografi, kjernemagnetisk resonans (NMR) spektroskopi, Hydroged deuterium exchange Mass Spectrometry (HX-MS) og forskjellige konkurransebindings-metoder. Ettersom hver metode avhenger av et unikt prinsipp er beskrivelse av en epitop nøye forbundet med metoden ved hvilken den har blitt bestemt. Følgelig vil epitopen for et gitt Ab/ Ag-par bli definert ulikt avhengig av hvilken metode for epitopkartlegging som er anvendt. På sitt mest detaljerte nivå kan epitopen for interaksjonen mellom Ag og Ab
16 defineres ved de romlige koordinatene som definerer atom kontaktene til stede i Ag- Ab-interaksjonen, så vel som informasjon om deres relative bidrag til bindings termodynamikken. På et mindre detaljert nivå kan epitopen karakteriseres ved de romlige koordinatene som definerer atom kontaktene mellom Ag og Ab. På et ytterligere mindre detaljert nivå kan epitopen karakteriseres ved aminosyrerestene som den omfatter som definert ved et bestemt kriterium, f.eks. avstand mellom atomer i Ab og Ag. På et enda mindre detaljert nivå kan epitopen karakteriseres gjennom funksjon, f.eks. ved konkurransebinding med andre Ab'er. Epitopen kan også defineres mer generisk som å omfatte aminosyrerester for hvilke substitusjon ved en annen aminosyre vil endre egenskapene av interaksjonen mellom Ab og Ag. I konteksten til en røntgen-avledet krystallstruktur definert ved romlige koordinater av et kompleks mellom et Ab, f.eks. et Fab-fragment, og dets Ag, er betegnelsen epitop heri, dersom ikke annerledes angitt eller det motsies av sammenhengen, spesifikt definert som K2-rester karakterisert ved at de har et tungt atom (dvs. et ikke-hydrogen atom) innenfor en avstand på 4 Å fra et tungt atom i Ab. Ut fra det faktum at beskrivelsene og definisjonene av epitoper, avhengig av metoden for epitopkartlegging anvendt, er oppnådd ved forskjellige detaljnivåer, følger det at sammenligning av epitoper for ulike Ab på samme Ag på lignende måte kan foretas på forskjellige detaljnivåer. Epitoper beskrevet på aminosyrenivå, f.eks. bestemt fra en røntgenstrukturen, sies å være identiske dersom de inneholder samme sett av aminosyrerester. Epitoper sies å overlappe dersom minst én aminosyre deles av epitopene. Epitoper sies å være atskilte (unike) dersom ingen aminosyrerest deles av epitopene. Epitoper karakterisert ved konkurrerende binding sies å være overlappende dersom binding av de korresponderende Ab'er er gjensidig utelukkende, dvs. binding av ett Ab utelukker samtidig binding av det andre Ab. Epitopene sies å være atskilte (unike) dersom Ag er i stand til romme binding av begge korresponderende Ab'er samtidig. Definisjonen av betegnelsen "paratop" er avledet fra ovennevnte definisjon av "epitop" ved å snu opp/ned på perspektivet. Følgelig angir betegnelsen "paratop" området eller regionen på Ab til hvilken Ag spesifikt binder, dvs. til hvilken det kommer i fysisk kontakt med Ag. I konteksten til en røntgen-avledet krystallstruktur definert ved romlige koordinater av et kompleks mellom et Ab, f.eks. et Fab-fragment, og dets Ag, er betegnelsen paratop heri, dersom ikke annerledes angitt eller det klart motsies av sammenhengen, spesifikt definert som Ag-rester krakterisert ved at de har et tungt
17 1 1 2 atom (dvs. et ikke-hydrogen atom) innenfor en avstand på 4 Å fra et tungt atom i K2. Epitopen og paratopen for et gitt antistoff (Ab)/antigen (Ag)-par kan identifiseres ved rutinemessige metoder. For eksempel kan den generelle lokaliseringen av en epitop bestemmes ved å vurdere evnen av et antistoff til å binde til ulike fragmenter eller variant TFPI polypeptider. De spesifikke aminosyrene i TFPI som kommer i kontakt med et antistoff (epitop) og de spesifikke aminosyrene i et antistoff som kommer i kontakt med TFPI (paratop) kan også bestemmes ved anvendelse av rutinemessige metoder, slik som de beskrevet i eksemplene. For eksempel kan antistoffet og mål-molekylet kombineres og Ab/ Ag-komplekset kan krystalliseres. Krystallstrukturen av komplekset kan bestemmes og anvendes for å identifisere spesifikke seter for interaksjon mellom antistoffet og dets mål. Foreliggende oppfinnere har utført en slik analyse av interaksjon mellom det murine antistoffet MuTFPI4F36, så vel som det humaniserte antistoffet HzTFPI4F36, beskrevet heri, og Kunitz 2-domenet (K2) av TFPI. Denne analysen er mer detaljert beskrevet i eksemplene. Paratopen av et antistoff ifølge foreliggende oppfinnelse kan defineres som følger: den lette kjeden av nevnte antistoff omfatter restene E31, S32, D33, Y37, A96, T97 og F99 av SEQ ID NO: 1 og den tunge kjeden av nevnte antistoff omfatter restene N31, S2, R3, S4, Y7, Y9, F60, P61, D62, Q6, Y2, D3 og D6 av SEQ ID NO 18. Den lette kjeden av antistoffet ifølge foreliggende oppfinnelse kan følgelig omfatte aminosyrerestene: E, i posisjonen svarende til posisjon 31, S, i posisjonen svarende til posisjon 32, D, i posisjonen svarende til posisjon 33, Y, i posisjonen svarende til posisjon 37, A, i posisjonen svarende til posisjon 96, T, i posisjonen svarende til posisjon 97 og F, i posisjonen svarende til posisjon 99 av SEQ ID NO: 1; og den tunge kjeden av nevnte antistoff kan omfatte aminosyrerestene: 3 N, i posisjonen svarende til posisjon 31, R, i posisjonen svarende til posisjon 3, S, i posisjonen svarende til posisjon 4, Y, i posisjonen svarende til posisjon 7,
18 Y, i posisjonen svarende til posisjon 9, F, i posisjonen svarende til posisjon 60, P, i posisjonen svarende til posisjon 61, D, i posisjonen svarende til posisjon 62, Q, i posisjonen svarende til posisjon 6, Y, i posisjonen svarende til posisjon 2, D, i posisjonen svarende til posisjon 3 og D, i posisjonen svarende til posisjon 6 av SEQ ID NO 18. Den tunge kjeden kan videre omfatte en S i posisjonen svarende til posisjon 2 fra SEQ ID NO: 18. Den lette kjeden av et antistoff ifølge foreliggende oppfinnelse kan videre omfatte en H i posisjonen svarende til posisjon 98 av SEQ ID NO: 1 og den tunge kjeden kan videre omfatte an S, i posisjonen svarende til posisjon 6 av SEQ ID NO: 18. For MuTFPI4F36 (Eksempel 4) ble epitopen funnet å være sammensatt av aminosyrene E0, E1, P3, R7, Y9, T111, Y113, Q118, Q121, E123, R124, F12, K126 og L140 fra SEQ ID NO: 1, svarende til aminosyrene E, E11, P13, R17, Y19, T21, Y23, Q28, Q31, E33, R34, F3, K36 og L0 fra SEQ ID NO: 2. Paratopen ble funnet å være sammensatt av lett kjede aminosyrerestene E31, S32, D33, Y37, A96, T97, H98 og F99 av SEQ ID NO: 4 og tung kjede aminosyrerestene N31, R3, S4, S6, Y7, Y9, F60, P61, D62, Q6, Y2, D3 og D6 av SEQ ID NO 8. For HzTFPI4F36 (Eksempel ) ble epitopen funnet å være sammensatt av aminosyrene E0, E1, D2, P3, R7, Y9, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F12, K126 og L140 fra SEQ ID NO: 1, svarende til aminosyrene E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, K36 og L0 av SEQ ID NO: 2. Paratopen ble funnet å være sammensatt av lett kjede aminosyrerestene E31, S32, D33, Y37, A96, T97 og F99 av SEQ ID NO: 1 og tung kjede aminosyrerestene N31, S2, R3, S4, Y7, Y9, F60, P61, D62, Q6, Y2, D3 og D6 av SEQ ID NO 18. Et antistoff i henhold til foreliggende oppfinnelse kan binde til samme epitop eller domene av TFPI som antistoffene ifølge oppfinnelsen som er spesifikt beskrevet heri. For eksempel kan andre enda uidentifiserte antistoffer ifølge oppfinnelsen identifiseres ved å sammenligne deres binding til TFPI med den for de monoklonale antistoffene, MuTFPI4F36 og/eller HzTFPI4F36; eller ved å sammenligne funksjonen av enda uidentifiserte antistoffer med den av MuTFPI4F36 og/eller HzTFPI4F36.
19 Analyser og assays som kan anvendes med henblikk på slik identifikasjon omfatter TFPI nøytraliserings assays slik som: FXa inhiberings assay beskrevet i Eksempel 6 og FVIIa/TF/FXa inhiberings assay beskrevet i Eksempel 7; bindingsinteraksjonsanalyser slik som overflate plasmonresonans-analyse beskrevet i Eksempel 8; cellulære assays slik som nøytralisering av TFPI ved humane vaskulære endotelceller fra navlestreng (HUVEC), beskrevet i Eksempel 9, og nøytralisering av TFPI-inhibering av TF/FVIIa-aktivitet på MDA-MB 231 humane brystkarcinomceller, beskrevet i Eksempel. I én utførelsesform kan et antistoff ifølge oppfinnelsen binde til samme epitop eller region som antistoffene MuTFPI4F36 eller HzTFPI4F36 beskrevet heri. Binding av MuTFPI4F36 og HzTFPI4F36 til TFPI er beskrevet mer detaljert heri. Et antistoff ifølge oppfinnelsen kan være et antistoff som binder til samme epitop i TFPI som MuTFPI4F36 eller HzTFPI4F36-antistoffene. Dette kan omfatte at det er i kontakt med de bestemte aminosyrene av TFPI som beskrevet ovenfor. For eksempel kan et antistoff ifølge oppfinnelsen binde til TFPI på en slik måte at det er i kontakt med aminosyrene E, E11, P13, R17, Y19, T21, Y23, Q28, Q31, E33, R34, F3, K36 og L0 av SEQ ID NO: 2. eller på en slik måte at det er i kontakt med aminosyrene E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, K36 og L0 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende én eller flere rester valgt fra gruppen bestående av E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, F3, K36 og L0 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest E av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest E11 av SEQ ID NO: 2). Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest D12 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest P13 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest R17 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest Y19 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest T21 av SEQ ID NO: 2.
20 Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest Y23 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest F24 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest N26 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest Q28 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest Q31 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest C32 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest E33 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest R34 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest F3 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest K36 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende rest L0 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende restene E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, K36 og L0 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan være i stand til å binde en epitop omfattende restene E, E11, P13, R17, Y19, T21, Y23, Q28, Q31, E33, R34, F3, K36 og L0 av SEQ ID NO: 2. Et antistoff ifølge oppfinnelsen kan ha evne til å konkurrere med et annet antistoff ifølge oppfinnelsen om binding til TFPI eller et annet passende mål som beskrevet heri. For eksempel kan et antistoff ifølge oppfinnelsen kryss-konkurrere med antistoffene MuTFPI4F36 eller HzTFPI4F36 beskrevet heri om binding til TFPI, eller til et passende fragment eller variant av TFPI som bindes av MuTFPI4F36 eller HzTFPI4F36-antistoffene. Slik kryss-konkurrerende antistoffer kan identifiseres basert på deres evne til å kryss-konkurrere med et kjent antistoff ifølge oppfinnelsen i standard bindingsassays. For eksempel kan SPR f.eks. ved anvendelse av et Biacore TM system, ELISA assay eller flowcytometri, anvendes for å demonstrere
21 kryss-konkurranse. Slik kryss-konkurranse kan antyde at de to antistoffene binder til identiske, overlappende eller lignende epitoper. Følgelig kan antistoffet ifølge oppfinnelsen være i stand til å binde K2- domenet av TFPI med en høyere affinitet enn en enkelt eller flere av de følgende kommersielt tilgjengelige monoklonale antistoffene: mab0281 (Ab systems) og/eller mab4904 (American Diagnostica) og/eller mab2974 (R&D systems) og/eller mab29741 (R&D systems). Et antistoff ifølge oppfinnelsen kan derfor identifiseres ved en fremgangsmåte som omfatter en bindings-assay som vurderer hvorvidt et test-antistoff er i stand til å konkurrere med et kjent antistoff ifølge oppfinnelsen om et bindingssete på målmolekylet. Metoder for utførelse av kompetitiv bindings-assays er velkjent på området. For eksempel kan de omfatte binding av et kjent antistoff ifølge oppfinnelsen til et mål-molekyl ved anvendelse av betingelser under hvilke antistoffet kan binde til mål-molekylet. Antistoff/mål-komplekset kan deretter eksponeres for et test-antistoff og graden i hvilken test-antistoffet er i stand til å fortrenge antistoffet ifølge oppfinnelsen fra antistoff/mål-komplekser kan vurderes. En alternativ metode kan omfatte å bringe et test-antistoff i kontakt med et målmolekyl under betingelser som tillater antistoffbinding, deretter tilsette et antistoff ifølge oppfinnelsen som er i stand til å binde dette mål-molekylet og fastsette graden i hvilken antistoffet ifølge oppfinnelsen er i stand til å fortrenge test-antistoffet fra antistoff/mål-komplekser. Evnen av et test-antistoff til å inhibere binding av et antistoff ifølge oppfinnelsen til målet viser at test-forbindelsen kan konkurrere med et antistoff ifølge oppfinnelsen om binding til målet og følgelig at test-antistoffet binder til samme epitop eller region på TFPI-proteinet som det kjente antistoffet ifølge oppfinnelsen. Et test-antistoff som er identifisert som å konkurrere med et kjent antistoff ifølge oppfinnelsen i en slik fremgangsmåte er også et potensielt antistoff i henhold til foreliggende oppfinnelse. Det faktum at test-antistoffet kan binde TFPI i samme region som et kjent antistoff ifølge oppfinnelsen og konkurrere med det kjente antistoffet ifølge oppfinnelsen antyder at test-antistoffet kan fungere som en ligand ved samme bindingssete som det kjente antistoffet og at test-antistoffet derfor kan etterligne virkningen av det kjente antistoffet. Dette kan bekreftes ved å vurdere aktiviteten av TFPI i nærvær av test-forbindelsen som beskrevet heri. Det kjente antistoffet ifølge oppfinnelsen kan være et antistoff som beskrevet heri, slik som det murine TFPI-4F36A1B2 (også referert til som 4F36 og som MuTFPI4F36)-antistoffet, eller hvilken som helst variant eller fragment derav som beskrevet heri som bibeholder evnen til å binde til TFPI, slik som humaniserte TFPI- 4F36A1B2 antistoffer, av hvilke ett er heri referert til som HzTFPI4F36 (mabtfpi
22 ). Et antistoff ifølge oppfinnelsen kan binde til samme epitop som MuTFPI4F36- antistoffet som beskrevet heri eller hvilken som helst variant eller fragment derav som beskrevet heri som bibeholder evnen til å binde til TFPI, slik som HzTFPI4F36. Et antistoff ifølge oppfinnelsen kan binde en epitop som er identisk med, overlapper, eller er lik MuTFPI4F36-epitopen som er ytterligere beskrevet i eksemplene. Et antistoff ifølge oppfinnelsen kan binde til en epitop som er identisk med, overlapper eller er lik HzTFPI4F36 epitopen som er ytterligere beskrevet i eksemplene. Et antistoff ifølge oppfinnelsen kan binde, foretrukket spesifikt, én eller flere aminosyrerester som tilhører epitopene fra MuTFPI4F36 og/eller HzTFPI4F36. For eksempel kan et antistoff ifølge oppfinnelsen binde til fem eller flere, seks eller flere, sju eller flere, åtte eller flere eller ti eller flere av aminosyrerestene vist ovenfor for binding av MuTFPI4F36 eller HzTFPI4F36. For eksempel, når det bringes i kontakt med et polypeptid ifølge SEQ ID NO: 2, kan et antistoff ifølge oppfinnelsen binde til polypeptidet og komme i kontakt med aminosyrene E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, F3, K36 og L0, eller en undergruppe av disse aminosyrene, slik som minst 2, minst 3, minst 4, minst, minst 6, minst 7, minst 8, minst 9, minst, minst 11, minst 12, minst 13, minst 14, minst 1, minst 16, minst 17 eller minst 18 av disse aminosyrene. Spesifikk binding kan vurderes med referanse til binding av antistoffet til et molekyl som ikke er målet. Denne sammenligningen kan foretas ved å sammenligne evnen av et antistoff til å binde til målet og til et annet molekyl. Denne sammenligningen kan foretas som beskrevet ovenfor i en vurdering av K D eller Ki. Det andre molekylet anvendt i en slik sammenligning kan være hvilket som helst molekyl som ikke er mål-molekylet. Foretrukket er det andre molekylet ikke identisk med mål-molekylet. Foretrukket er mål-molekylet ikke et fragment av målmolekylet. K D for et antistoff ifølge foreliggende oppfinnelse kan være lavere enn 0,8 nm, slik som lavere enn 0,7 nm, slik som lavere enn 0,6 nm, slik som lavere enn 0, nm, slik som lavere enn 0,4 nm, slik som lavere enn 0,3 nm, slik som lavere enn 0,2 nm, slik som lavere enn 0,1 nm, slik som lavere enn 0,0 nm, slik som lavere enn 0,02 nm, slik som lavere enn 0,01 nm, slik som mellom 0,01 nm og 0 nm. Det andre molekylet anvendt for å bestemme spesifikk binding kan være urelatert i struktur eller funksjon med målet. For eksempel kan det andre molekylet være et urelatert materiale eller ledsagende materiale i omgivelsene. Det andre molekylet anvendt for å bestemme spesifikk binding kan være et annet molekyl involvert i samme in vivo reaksjonsvei som mål-molekylet. For eksempel kan, når målet er TFPI eller et fragment eller variant derav, det andre molekylet anvendt for sammenligning være et protein som danner del av
23 blodkoagulasjonskaskaden. Ved å sørge for at antistoffet ifølge oppfinnelsen har spesifisitet for TFPI fremfor et annet slikt molekyl, kan uønsket in vivo kryssreaktivitet unngås. Det andre molekylet anvendt for sammenligning kan være beslektet med mål-molekylet. Når det for eksempel er ønsket å identifisere et antistoff som binder kun til en spesifikk epitop, kan det andre molekylet for sammenligning være et TFPImolekyl i hvilket epitopen mangler eller er avbrutt. Det andre molekylet anvendt for sammenligning kan følgelig være et annet mål-molekyl som er forskjellig fra målmolekylet bundet av det aktuelle antistoffet. Antistoffet ifølge oppfinnelsen kan bibeholde evnen til å binde til noen molekyler som er beslektet med mål-molekylet. For eksempel kan en fullengde moden human TFPI anvendes som målet, men antistoffet kan også være i stand til å binde til, f.eks. umodne former av human TFPI, fragmenter eller trunkerte former av human TFPI, TFPI som er bundet til lipoprotein eller til en celle eller TFPI fra andre arter, slik som annen mammalsk TFPI. Alternativt kan antistoffet ifølge oppfinnelsen ha spesifisitet for et bestemt mål-molekyl. For eksempel kan det binde til ett mål-molekyl som beskrevet heri, men kan ikke binde, eller kan binde med betydelig redusert affinitet til et ulikt målmolekyl som beskrevet heri. For eksempel kan en fullengde moden human TFPI anvendes som målet, men antistoffet som binder til dette målet kan være ute av stand til å binde til eller kan binde med lavere affinitet til, f.eks. umodne former av human TFPI, fragmenter eller trunkerte former av human TFPI, TFPI som er bundet til lipoprotein eller til en celle eller TFPI fra annen art, slik som annen mammalsk TFPI. Et antistoff ifølge oppfinnelsen kan binde til TFPI og ved å gjøre dette kan det inhibere en aktivitet av TFPI. Som forklart ovenfor nedregulerer TFPI blodkoagulasjon. Den gjør det ved å hemme aktiviteten av FXa og ved å hemme TF-FVIIa-komplekset i nærvær av FXa. Aktiviteten av TFPI som inhiberes av et antistoff ifølge oppfinnelsen kan være hvilke som helst av disse aktivitetene eller hvilken som helst nedstrøms effekt derav. For eksempel kan et antistoff ifølge oppfinnelsen føre til en økning i blodkoagulasjon, økning i tilstedeværelse eller nivåer av FXa eller økt aktivitet av TF-FVIIa. Foretrukket reduserer et antistoff ifølge oppfinnelsen koagulasjonstid når det bringes i kontakt med (a) human plasma med FVIII-mangel eller (b) humant helblod. Måling av TFPI-aktivitet kan omfatte å vurdere aktiviteten av TFPI med hensyn til å inhibere koagulasjon eller redusere koagulasjonstid i en blodprøve. For eksempel kan en slik fremgangsmåte omfatte å bringe TFPI i kontakt med en prøve av blod eller et blodprodukt slik som plasma eller serum som omfatter
24 blodkoagulasjons-faktorer ved betingelser under hvilke koagulasjon skulle finne sted, og bestemme hvorvidt koagulasjon av blodet inhiberes eller koagulasjonstid reduseres ved nærvær av TFPI. Nivået av blodkoagulasjon eller koagulasjonstid i en slik prøve kan deretter sammenlignes med det i en ekvivalent prøve i hvilken et testantistoff også er til stede. Dersom nivået av koagulasjon er økt eller koagulasjonstid er redusert i antistoffprøven, antyder dette at antistoffet inhiberer aktiviteten av TFPI i prøven. Blodkoagulasjon kan påvises ved å se etter koagulasjon av blodet i seg selv, i plasma, eller én eller flere karakteristika for koagulasjonskaskaden som finnes nedstrøms for virkningspunktet for TFPI. For eksempel kan fremgangsmåten fastsette nivåer av FXa eller aktivering av TF-FVIIa i prøven. Forskjellige andre metoder for å vurdere blodkoagulasjon og koagulasjonstid er velkjent på området. For eksempel kan hvilken som helst effekt av et antistoff på blodkoagulasjonstiden vurderes ved anvendelse av "dilute" protrombintid-analyse (dpt analyse) som beskrevet i eksemplene. I korthet blir humant plasma brakt i kontakt med humant tromboplastin. Tiden det tar for plasma å koagulere blir målt i nærvær og fravær av test-antistoffet. En positiv kontroll kan anvendes i en slik analyse, slik som tilsetning av FVIIa (NovoSeven ) som ville forventes å redusere koagulasjonstid. Et antistoff ifølge oppfinnelsen skulle være i stand til redusere koagulasjonstid i en slik metode. Foretrukket skulle et antistoff ifølge oppfinnelsen være i stand ti å redusere koagulasjonstid på en doseavhengig måte. Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å inhibere TFPI i en plasmabasert koagulasjons-assay, slik som en dpt-analyse, betydelig bedre enn et enkelt eller flere av de følgende kommersielt tilgjengelige monoklonale antistoffene: mab0281 (Ab systems) og/eller mab4904 (American Diagnostica) og/eller mab2974 (R&D systems) og/eller mab29741 (R&D systems). Tromboelastografi kan anvendes for å vurdere kinetikken for koagulasjon og fibrinolyse i prøver av helblod. Evnen av et antistoff til å redusere koagulasjonstid eller til å stimulere blodkoagulasjon kan følgelig vurderes på lignende måte i en helblodprøve ved å sammenligne tiden det tar for dannelse av klumper i nærvær og fravær av antistoffet. Metoder for å vurdere de funksjonelle effektene av et antistoff ifølge oppfinnelsen kan følgelig utføres in vitro. Slike metoder blir foretrukket utført for prøver av humant blod eller plasma. Slike prøver kan være normalt humant blod eller plasma eller kan ha mangel av, eller være supplert med, én eller flere faktorer involvert i blodkoagulasjon. For eksempel kan disse metodene utføres ved bruk av normalt humant helblod, normalt humant plasma eller plasma eller helblod med FVIII-mangel. Blod eller plasma med FVIII- mangel kan frembringes ved å bringe
25 egnet blod- eller plasmaprøve i kontakt med nøytraliserende anti-fviii antistoff. Slike in vitro metoder kan være bindingsinteraksjonsanalyser eller TFPI nøytraliserings-analyser, slik som de beskrevet i eksemplene Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å inhibere blodplate-assosiert TFPI. Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å inhibere løselig TFPI. Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å inhibere lipoprotein-bundet TFPI. Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å inhibere celle-bundet TFPI, slik som TFPI som er bundet til endotelceller. Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å binde TFPI slik at FXa bibeholder sin aktivitet med minst 91 %, slik som minst 92 %, slik som minst 93 %, slik som minst 94 %, slik som minst 9 %, slik som minst 96 %, slik som minst 97 %, slik som minst 98 %, slik som minst 99 %, slik som 99-0 % som målt i en FXa inhiberings-assay. Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å nøytralisere TFPI-inhiberingen av membranbundet FVIIa/TF/FXa, når TFPI er mettet med nevnte antistoff, ved minst %, slik som minst 60 %, slik som minst 6 %, slik som minst 70 %, slik som minst 7 %, slik som minst 80 %, slik som minst 8 %, slik som minst 90 %, slik som minst 9 %, slik som opp til 0 %, slik som 0 %, som målt i en FVIIa/TF/FXa inhibitor-assay. Foretrukket er et antistoff ifølge oppfinnelsen i stand til å redusere koagulasjonstid og/eller stimulere blodkoagulasjon i en prøve av (a) humant helblod, (b) humant plasma, (c) humant helblod med FVIII-mangel, (d) humant plasma med FVIII-mangel, (e) humant helblod med FIX-mangel eller (f) humant plasma med FIX-mangel. Metoder for å bestemme evnen av et antistoff til å stimulere blodkoagulasjon eller redusere koagulasjonstid kan også utføres in vivo. For eksempel kan in vivo studier utføres i transient hemofile kaniner som beskrevet i eksemplene. I korthet kan kaniner gjøres transient hemofile ved administrering av anti-fviii antistoff. Test-antistoffet kan deretter administreres og blødningstiden i huden og/eller blodplatetallet fastsettes. En reduksjon i blødningstiden i huden i nærvær av et testantistoff indikerer at antistoffet er i stand til å redusere koagulasjonstid og stimulere blodkoagulasjon. Et antistoff som har en slik effekt kan derfor være et antistoff ifølge foreliggende oppfinnelse. Antistoffet ifølge foreliggende oppfinnelse kan være i stand til å binde K2- domenet av TFPI slik at prosentandelen av fri TFPI i et subjekt blir redusert til
26 mindre enn %, slik som mindre enn 29 %, slik som mindre enn 28 %, slik som mindre enn 27 %, slik som mindre enn 26 %, slik som mindre enn 2 %, slik som mindre enn 24 %, slik som mindre enn 23 %, slik som mindre enn 22 %, slik som mindre enn 21 %, slik som mindre enn %, slik som mindre enn 19 %, slik som mindre enn 18 %, slik som mindre enn 17 %, slik som mindre enn 16 %, slik som mindre enn 1 %, slik som mindre enn 14 %, slik som mindre enn 13 %, slik som mindre enn 12 %, slik som mindre enn 11 %, slik som mindre enn %, slik som mindre enn 9 %, slik som mindre enn 8 %, slik som mindre enn 7 %, slik som mindre enn 6 %, slik som mindre enn %, slik som mindre enn 4 %, slik som mindre enn 3 %, slik som mindre enn 2 %, slik som mindre enn 1 %, slik som 0 %. Videre kan antistoffet ifølge foreliggende oppfinnelse være i stand til å binde K2-domenet av TFPI slik at mengden av fri TFPI i et subjekt blir redusert i løpet av de første 28 dagene, slik som i løpet av de første 27 dager, slik som i løpet av de første 26 dagene, slik som i løpet av de første 2 dagene, slik som i løpet av de første 24 dagene, slik som i løpet av de første 23 dagene, slik som i løpet av de første 22 dagene, slik som i løpet av de første 21 dagene, slik som i løpet av de første dagene, slik som i løpet av de første 19 dagene, slik som i løpet av de første 18 dagene, slik som i løpet av de første 17 dagene, slik som i løpet av de første 16 dagene, slik som i løpet av de første 1 dagene, slik som i løpet av de første 14 dagene, slik som i løpet av de første 13 dagene, slik som i løpet av de første 12 dagene, slik som i løpet av de første 11 dagene, slik som i løpet av de første dagene, slik som i løpet av de første 9 dagene, slik som i løpet av de første 8 dagene, slik som i løpet av de første 7 dagene, slik som i løpet av de første 6 dagene, slik som i løpet av de første dagene, slik som i løpet av de første 4 dagene, slik som i løpet av de første 3 dagene, slik som i løpet av de første 2 dagene, slik som i løpet av den første dagen etter administrering av nevnte monoklonale antistoff til nevnte subjekt. Et antistoff ifølge foreliggende oppfinnelse kan også føre til ingen signifikant reduksjon i blodplatetallene. Spesielt kan et antistoff ifølge oppfinnelsen være i stand til å redusere koagulasjonstid og/eller stimulere blodkoagulasjon i en prøve av (a) humant helblod, (b) humant plasma, (c) humant helblod med FVIII-mangel (d) humant plasma med FVIII-mangel, (e) humant helblod med FIX-mangel eller (f) humant plasma med FIX-mangel, eller i et dyr in vivo, uten å føre til noen signifikant reduksjon i blodplatetall. Blodplatetall kan vurderes i samme prøve eller dyr som de andre effektene diskutert ovenfor, eller kan vurderes separat. For eksempel kan blodplatetall vurderes i en blodprøve slik som en prøve av blod oppnådd fra en pasient eller forsøksdyr. Blodplatetall kan vurderes etter administrering av antistoffet til en kanin med transient hemofili som beskrevet ovenfor. Antistoffer ifølge
27 oppfinnelsen kan være i stand til å redusere blødningstiden i huden uten å føre til en samtidig reduksjon i blodplatetall, som eksemplifisert ved in vivo studier i transient hemofile kaniner. En endring i blodplatetall kan vurderes ved å sammenligne blodplatetall før og etter administrering av antistoffet eller ved å sammenligne blodplatetall mellom en prøve eller dyr behandlet med antistoffet av interesse og en kontrollprøve eller dyr ikke behandlet med dette antistoffet. Et antistoff ifølge foreliggende oppfinnelse kan være i stand til å binde K2-domenet av TFPI, slik at et subjekts in vivo koagulasjonstid blir redusert og blodplatetallet hos nevnte subjekt ikke blir betydelig redusert. For eksempel kan nevnte subjekts blodplatetall ikke synke til omtrent 80 %, slik som omtrent 7 %, slik som omtrent 70 %, slik som omtrent 6 %, slik som omtrent 60 %, slik som omtrent %, slik som omtrent 0 %, slik som omtrent 4 %, slik som omtrent 40 %, slik som omtrent 3 %, slik som omtrent %, slik som omtrent 2 % av den opprinnelige blodplatetellingen. Foretrukket vil det ikke være noen forskjell eller ingen statistisk signifikant forskjell i blodplatetall når slike sammenligninger foretas. Det vil si at antistoffet ifølge oppfinnelsen ikke vil ha forårsaket noen reduksjon i blodplatetall. Betegnelsen "antistoff" som referert til heri omfatter hele antistoffer og hvilket som helst antigenbindende fragment (dvs., "antigen-bindende del") eller enkeltkjeder derav. Et antistoff angir et glykoprotein omfattende minst to tunge kjeder (HC) og to lette kjeder (LC) innbyrdes forbundet ved disulfidbindinger, eller en antigenbindende del derav. Hver tunge kjede er sammensatt av en tung kjede variabel region (forkortet heri som VH) og en tung kjede konstant region (CH). Hver lette kjede er sammensatt av en lett kjede variabel region (forkortet heri som VL) og en lett kjede konstant region (CL). De variable regionene av de tunge og lette kjedene inneholder et bindingsdomene som interagerer med et antigen. VH og VLregionene kan deles videre opp i regioner av hypervariabilitet, betegnet komplementaritetsbestemmende regioner (CDR), med innlagte regioner som er mer konserverte, betegnet struktur ( framework )-regioner (FR). De konstante regionene av antistoffene kan mediere binding av immunglobulinet til vertsvev eller faktorer, inkludert ulike celler av immunsystemet (f.eks., effektorceller) og den første komponenten (CIq) av det klassiske komplementsystemet. Betegnelsen "komplementaritetsbestemmende region" eller "hypervariabel region" angir, når anvendt heri, de aminosyrerestene av et antistoff som er ansvarlig for antigenbinding. De komplementaritetsbestemmende regionene "CDR'ene" utgjøres generelt av aminosyrerestene (L1), 0-6 (L2) og (L3) i lett kjede variabelt domene og 31-3 (Hl), 0-6 (H2) og 9-2 (H3) i tung kjede variabelt domene; (Kabat et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH
28 Publication No ) og/eller restene fra en "hypervariabel løkke" (restene (L1), 0-2 (L2) og (L3) i lett kjede variabelt domene og (H1), 3- (H2) og 96-1 (H3) i tung kjede variabelt domene; Chothia og Lesk, J. MoI. Biol 1987; 196: ). Typisk blir nummereringen av aminosyrerestene i denne regionen foretatt ved metoden beskrevet i Kabat et al., supra. Uttrykk slik som "Kabat posisjon", "Kabat-rest", og "i henhold til Kabat" refererer til dette nummereringssystemet for tung kjede variable domener eller lett kjede variable domener. Ved anvendelse av Kabat nummereringssystemet, kan den reelle lineære aminosyresekvensen av et peptid inneholde færre eller ekstra aminosyrer samsvarende med en forkortelse av, eller insersjon inn i, en FR eller CDR av det variable domenet. For eksempel kan et tung kjede variabelt domene omfatte aminosyreinsersjoner (rest 2a, 2b og 2c i henhold til Kabat) etter rest 2 av CDR H2 og inserterte rester (f.eks. restene 82a, 82b og 82c, osv. i henhold til Kabat) etter tung kjede FR rest 82. Kabat-nummereringen av rester kan bestemmes for et gitt antistoff ved sammenstilling ved regioner med homologi av sekvensen av antistoffet med en "standard" Kabat-nummerert sekvens. Betegnelsen " struktur ( framework )-region" eller "FR"-rester angir de VH eller VL-aminosyrerestene som ikke er innenfor CDR'ene, som definert heri. Et antistoff ifølge oppfinnelsen kan være et monoklonalt antistoff eller et polyklonalt antistoff. I én utførelsesform er et antistoff ifølge oppfinnelsen et monoklonalt antistoff. Et antistoff ifølge oppfinnelsen kan være et kimært antistoff, et CDR-bundet antistoff, et humant eller humanisert antistoff eller en antigenbindende del av hvilke som helst derav. For produksjon av både monoklonale og polyklonale antistoffer, er forsøksdyret et passende pattedyr slik som, men ikke begrenset til, en geit, kanin, rotte eller mus. Polyklonale antistoffer er antistoffer som er avledet fra forskjellige B cellelinjer. Et polyklonalt antistoff kan omfatte en blanding av forskjellige immunglobulinmolekyler som er rettet mot et spesifikt antigen. Det polyklonale antistoffet kan omfatte en blanding av forskjellige immunglobulin-molekyler som binder til én eller flere ulike epitoper på et antigenmolekyl. Polyklonale antistoffer kan frembringes ved rutinemessige metoder slik som immunisering av et egnet dyr, med antigenet av interesse. Blod kan senere fjernes fra dyret og immunglobulinfraksjonen renses. Monoklonale antistoffer er immunglobulinmolekyler som er identiske med hverandre og har en enkelt bindingsspesifisitet og affinitet for en bestemt epitop. Monoklonale antistoffer (mabs) ifølge foreliggende oppfinnelse kan frembringes ved mange forskjellige teknikker, inkludert konvensjonell monoklonalt antistoffmetodologi f.eks., standard somatisk celle hybridiseringsteknikk ifølge Kohler og
29 Milstein (197) Nature 26: 49, eller viral eller oncogen transformasjon av B- lymfocytter. Det foretrukne animalske systemet for fremstilling av hybridomer er det murine systemet. Hybridom produksjon i mus er en svært veletablert metode. Immuniseringsmetoder og teknikker for isolering av immuniserte splenocytter for fusjon er kjent på området. Fusjonspartnere (f.eks., murine myelomceller) og fusjonsmetoder er også kjent. For å fremstille hybridomer som produserer monoklonale antistoffer ifølge oppfinnelsen kan splenocytter og/eller lymfekjertel-celler fra immuniserte mus isoleres og fusjoneres med en egnet immortalisert cellelinje, slik som en mus myelom cellelinje. De resulterende hybridomene kan screenes for produksjon av antigen-spesifikke antistoffer. De antistoff-sekreterende hybridomene kan kan plates ut på nytt, screenes igjen, og dersom fortsatt positive for passende IgG, kan de monoklonale antistoffeme subklones minst to ganger ved begrensende fortynning. De stabile subklonene kan deretter dyrkes in vitro for å fremstille små mengder av antistoff i vevskulturmedium for karakterisering. Betegnelsen "antigenbindende del" av et antistoff angir ett eller flere fragmenter av et antistoff som bibeholder evne til å spesifikt binde til et antigen, slik som TFPI eller et annet mål-protein som beskrevet heri. Det er vist at den antigenbindende funksjonen av et antistoff kan utføres av fragmenter av et fullengde antistoff. Eksempler på bindende fragmenter omfattet innenfor betegnelsen "antigen-bindende del" av et antistoff omfatter et Fab-fragment, et F(ab') 2 -fragment, et Fab'-fragment, et Fd-fragment, et Fv-fragment, et dab-fragment og en isolert komplementaritets-bestemmende region (CDR). Enkeltkjede antistoffer slik som scfv og tung kjede antistoffer slik som VHH og kamel antistoffer er også ment å være omfattet innenfor betegnelsen "antigen-bindende del" av et antistoff. Disse antistoffragmentene kan oppnås ved anvendelse av konvensjonelle teknikker kjent for fagfolk på området, og fragmentene kan screenes for anvendelighet på samme måte som intakte antistoffer. Et antistoff ifølge oppfinnelsen kan fremstilles, uttrykkes, dannes eller isoleres ved rekombinante metoder, slik som (a) antistoffer isolert fra et dyr (f.eks., en mus) som er transgent eller transkromosomalt for immunglobulingenene av interesse eller et hybridom fremstilt derfra, (b) antistoffer isolert fra en vertscelle transformert for å uttrykke antistoffet av interesse, f.eks., fra et transfectom, (c) antistoffer isolert fra et rekombinant, kombinatorisk antistoff-bibliotek, og (d) antistoffer fremstilt, uttrykt, dannet eller isolert ved hvilke som helst andre metoder som omfatter splicing av immunglobulin gensekvenser til andre DNA-sekvenser. Et antistoff ifølge oppfinnelsen kan være et humant antistoff eller et humanisert antistoff. Betegnelsen "humant antistoff", som anvendt heri, skal
30 omfatte antistoffer som har variable regioner i hvilke både struktur ( framework )- og CDR-regionene er avledet fra human kjønnscelle immunglobulinsekvenser. Dersom antistoffet inneholder en konstant region er den konstante regionen også avledet fra immunglobulinsekvenser fra human kjønnscelle. De humane antistoffene ifølge oppfinnelsen kan omfatte aminosyrerester som ikke kodes for av human kjønnscelle immunglobulinsekvenser (f.eks., mutasjoner innført ved tilfeldig eller sete-rettet mutagenese in vitro eller ved somatisk mutasjon in vivo). Imidlertid er betegnelsen "humant antistoff", som anvendt heri, ikke ment å omfatte antistoffer i hvilke CDR-sekvenser avledet fra kjønnscellen fra en annen pattedyrart, slik som en mus, har blitt bundet ("grafted") til humane struktur ( framework )-sekvenser. Et slikt humant antistoff kan være et humant monoklonalt antistoff. Et slikt humant monoklonalt antistoff kan produseres av et hybridom som omfatter en B- celle oppnådd fra et transgent ikke-humant dyr, f.eks., en transgen mus, som har et genom omfattende et human tung kjede transgen og et lett kjede transgen fusjonert til en immortalisert celle. Humane antistoffer kan isoleres fra sekvensbibliotek bygget på utvalg av human kjønnscellesekvenser ytterligere diversifisert med naturlig og syntetisk sekvens diversitet. Humane antistoffer kan fremstilles ved in vitro immunisering av humane lymfocytter fulgt av transformasjon av lymfocyttene med Epstein-Barr virus. Betegnelsen "humane antistoff-derivater" angir hvilken som helst modifisert form av det humane antistoffet, f.eks., et konjugat av antistoffet og et annet middel eller antistoff. Betegnelsen "humanisert antistoff" er ment å angi et humant/ ikke-humant kimært antistoff som inneholder en minimal sekvens (CDR-regioner) avledet fra ikke-humant immunglobulin. Humaniserte antistoffer er følgelig humane immunglobuliner (recipient antistoff) i hvilke rester fra en hypervariabel region av recipienten er erstattet av rester fra en hypervariabel region fra en ikke-human art (donor-antistoff) slik som mus, rotte, kanin, eller ikke-human primat som har den ønskede spesifisiteten, affiniteten og kapasitet. I noen tilfeller er FR-rester fra det humane immunglobulinet erstattet av tilsvarende ikke-humane rester. Et eksempel på en slik modifikasjon er innføring av én eller flere såkalte tilbake-mutasjoner, slik som beskrevet i Eksempel 2. Videre kan humaniserte antistoffer omfatte rester som ikke finnes i recipientantistoffet eller i donor-antistoffet. Disse modifikasjonene foretas for å ytterligere foredle antistoff ytelse. Generelt vil et humanisert antistoff omfatte hovedsakelig alt av minst ett, og typisk to, variable domener, i hvilke alt eller hovedsakelig alt av de hypervariable løkkene svarer til slike som er fra et ikke-humant immunglobulin og
31 alt eller hovedsakelig alt av FR-restene er fra en human immunglobulinsekvens. Det humaniserte antistoffet kan eventuelt også omfatte minst en del av en immunglobulin konstant region (Fc), typisk den fra et humant immunglobulin. Antistoffer ifølge oppfinnelsen kan testes for binding til målproteinet ved, for eksempel, standard ELISA eller Western blotting. En ELISA assay kan også anvendes for å screene for hybridomer som oppviser positiv reaktivitet med målproteinet. Bindingsspesifisiteten av et antistoff kan også bestemmes ved å overvåke binding av antistoffet til celler som uttrykker målproteinet, for eksempel ved flowcytometri. Spesifisiteten av et antistoff ifølge oppfinnelsen for mål-protein kan studeres ytterligere ved å bestemme om antistoffet binder til andre proteiner eller ikke. Når det for eksempel er ønsket å fremstille et antistoff som spesifikt binder TFPI eller en bestemt del, f.eks. epitop, av TFPI, kan spesifisiteten av antistoffet vurderes ved å bestemme hvorvidt antistoffet også binder til andre molekyler eller modifiserte former av TFPI som mangler delen av interesse. Som forklart ovenfor kan antistoffer ifølge oppfinnelsen modifisere aktiviteten av TFPI. Antistoffer som har de nødvendige bindingsegenskapene kan følgelig testes ytterligere for å bestemme deres effekter på aktiviteten av TFPI. Følgelig kan det anvendes metoder for å identifisere egnede antistoffer som er i stand til å binde til TFPI og som er i stand til å modifisere, og spesielt redusere dens aktivitet. Når et passende antistoff først har blitt identifisert og selektert, kan aminosyresekvensen av antistoffet identifiseres ved metoder kjent på området. Genene som koder for antistoffet kan klones ved anvendelse av spesifikke og/eller degenererte primere. Antistoffet kan fremstilles rekombinant ved rutinemessige metoder. Et "polypeptid" blir anvendt heri i videste forstand til å referere til en forbindelse av to eller flere aminosyre subenheter, aminosyreanaloger, eller andre peptidomimetika. Betegnelsen "polypeptid" omfatter følgelig korte peptidsekvenser og også lengre polypeptider og proteiner. Som anvendt heri kan betegnelsen "aminosyre" referere til naturlige og/eller ikke-naturlige eller syntetiske aminosyrer, D og/eller L optiske isomerer, og aminosyreanaloger og peptidomimetika. Foreliggende oppfinnere har identifisert et murint antistoff som beskrevet i eksemplene. Dette antistoffet er referert til heri som TFPI-4F36A1B2 (Alternativt, 4F36 eller MuTFPI4F36). Foreliggende oppfinnelse omfatter dette antistoffet, varianter og fragmenter derav - inkludert kimære antistoffer og humaniserte antistoffer - som bibeholder én eller flere aktiviteter av det murine antistoffet og som også er beskrevet i eksemplene. Aktivitetene av dette antistoffet omfatter evne til å binde til TFPI, evne til å binde til spesifikke lokaliseringer i TFPI-molekylet og evne til å inhibere aktiviteten av TFPI.
32 1 2 3 Et egnet fragment eller variant av dette antistoff vil bibeholde evnen til å binde til TFPI. Det vil foretrukket bibeholde evnen til å spesifikt binde til TFPI. Det vil foretrukket bibeholde evnen til å spesifikt binde til samme eller lignende epitop eller region av TFPI-molekylet som antistoffet (MuTFPI4F36) fra hvilket det er avledet. Det vil foretrukket bibeholde én eller flere ytterligere funksjoner av antistoffet fra hvilket det er avledet, slik som evne til å inhibere TFPI-aktivitet eller evnen til å redusere koagulasjonstid, eventuelt uten at det fører til at blodplatetallet synker. Polypeptid- eller antistoff-"fragmenter" ifølge oppfinnelsen kan fremstilles ved trunkering, f.eks. ved fjerning av én eller flere aminosyrer fra de N- og/eller C- terminale endene av et polypeptid. Opp til, opp til, opp til, opptil 40 eller flere aminosyrer kan fjernes fra den N- og/eller C-terminale enden på denne måten. Fragmenter kan også fremstilles ved én eller flere indre delesjoner. Et antistoff ifølge oppfinnelsen kan være, eller kan omfatte, et fragment av antistoffet MuTFPI4F36 eller en variant derav. Antistoffet ifølge oppfinnelsen kan være eller kan omfatte en antigenbindende andel av dette antistoffet eller en variant derav som ytterligere diskutert ovenfor. For eksempel kan antistoffet ifølge oppfinnelsen være et Fab-fragment av dette antistoffet eller en variant derav eller kan være et enkeltkjede antistoff avledet fra dette antistoffet eller en variant derav. Aminosyresekvensene av de lette og tunge kjedene av MuTFPI4F36- antistoffet er gitt i SEQ ID NO: 6 og henholdsvis. Aminosyresekvensene for VL og VH-kjedene av antistoffet MuTFPI4F36 er gitt i SEQ ID NO: 4 og 8, henholdsvis. Aminosyresekvensene av de lette og tunge kjedene av ett humanisert antistoff, HzTFPI4F36, er gitt i SEQ ID NO: 21 og 24, henholdsvis. Aminosyresekvensene for VL og VH-kjedene av HzTFPI4F36 er gitt i SEQ ID NO: 1 og 18, henholdsvis. Et antistoff ifølge oppfinnelsen kan omfatte lett kjede aminosyresekvensen fra MuTFPI4F36 vist i SEQ ID NO: 6 eller et fragment eller variant derav. Et antistoff kan i tillegg eller alternativt omfatte MuTFPI4F36 tung kjede aminosyresekvensen vist i SEQ ID NO: eller et fragment eller variant derav som beskrevet heri. Et antistoff ifølge oppfinnelsen kan omfatte VL aminosyresekvensen av SEQ ID No: 4, eller et fragment eller variant derav. Et antistoff ifølge oppfinnelsen kan omfatte VH-aminosyresekvensen ifølge SEQ ID No: 8, eller et fragment eller variant derav. Et antistoff ifølge oppfinnelsen kan omfatte både (a) aminosyresekvensen for VL av SEQ ID No: 4, eller et fragment eller variant derav og (b) aminosyresekvensen for VH av SEQ ID No: 8, eller et fragment eller variant derav. Et antistoff ifølge oppfinnelsen kan omfatte et fragment av én av VL eller VH aminosyresekvensene vist i Figur 2. For eksempel kan et antistoff ifølge oppfinnelsen omfatte et fragment med minst 7, minst 8, minst 9, minst, minst 12, minst 1, minst 18, minst eller minst 2 etterfølgende aminosyrer av SEQ ID No: 4 eller 8.
33 Et slikt fragment vil foretrukket bibeholde én eller flere av funksjonene diskutert ovenfor, slik som evne til å binde til TFPI. Et egnet fragment eller variant av hvilke som helst av disse VH eller VLsekvensene vil bibeholde evnen til å binde til TFPI. Det vil foretrukket bibeholde evnen til å spesifikt binde til TFPI. Det vil foretrukket bibeholde evnen til å spesifikt binde til samme eller lignende epitop eller region av TFPI-molekylet som antistoffet (MuTFPI4F36) fra hvilket det er avledet. Det vil foretrukket bibeholde én eller flere ytterligere funksjoner av antistoffet fra hvilket det er avledet, slik som evnen til å inhibere TFPI-aktivitet eller evnen til å redusere koagulasjonstid, eventuelt uten å føre til at blodplatetallet synker. Et passende fragment eller variant VL-sekvens vil foretrukket bibeholde aminosyrene ved posisjonene E31, S32, D33, Y37, A96, T97, H98 og F99 i SEQ ID NO: 4. Et passende fragment eller variant VH-sekvens vil foretrukket bibeholde aminosyrene ved posisjonene N31, R3, S4, S6, Y7, Y9, F60, P61, D62, Q6, Y2, D3 og D6 i SEQ ID NO: 8. Et passende fragment eller variant antistoff vil foretrukket bibeholde aminosyrene ved posisjonene E31, S32, D33, Y37, A96, T97, H98 og F99 i SEQ ID NO: 4 og aminosyrene ved posisjonene N31, R3, S4, S6, Y7, Y9, F60, P61, D62, Q6, Y2, D3 og D6 i SEQ ID NO: 8. Som identifisert i Figur 3 er disse restene i MuTFPI4F36 lett og tung kjede-sekvenser som har et tungt atom innenfor en avstand på 4 Å fra et tungt atom når MuTFPI4F36 er bundet til K2-domenet av TFPI. Et antistoff ifølge oppfinnelsen kan omfatte en CDR-region fra det spesifikke antistoffet identifisert heri slik som en CDR-region fra innenfor SEQ ID NO: 4 eller 8. Et slikt antistoff vil foretrukket bibeholde evnen til å binde til TFPI som beskrevet heri. For eksempel kan, som vist i Figur 3, ved anvendelse av Kabat-definisjonen, CDR-sekvensene innenfor den lette kjeden av MuTFPI4F36 identifiseres ved aminosyrene 24 til 39, til 61 og 94 til 2 av SEQ ID NO: 4 eller SEQ ID NO: 6. CDR-sekvensene innenfor den tunge kjeden av MuTFPI4F36 kan identifiseres ved aminosyrene 31 til 3, 0 til 66 og 99 til 1 av SEQ er NO: 8 eller SEQ ID NO:. Et antistoff ifølge oppfinnelsen kan omfatte én eller flere av CDR-sekvensene vist i Figur 3. For eksempel kan et antistoff ifølge oppfinnelsen omfatte én, to eller alle tre av aminosyresekvensene vist ved restene 24 til 39, til 61 og 94 til 2 av SEQ ID NO: 6. Et antistoff ifølge oppfinnelsen kan alternativt eller i tillegg omfatte én, to eller alle tre av aminosyresekvensene vist ved restene 31 til 3, 0 til 66 og 99 til 1 av SEQ ID NO:. Et antistoff ifølge oppfinnelsen kan omfatte alle seks aminosyresekvensene vist ved restene 24 til 39, til 61 og 94 til 2 av SEQ ID NO: 6 og 31 til 3, 0 til 66 og 99 til 1 av SEQ ID NO:.
34 Et antistoff ifølge oppfinnelsen kan være et humanisert antistoff, slik som antistoffet heri referert til som HzTFPI4F36 (mabtfpi 21). Et slikt antistoff kan omfatte én eller flere CDR-regioner fra innenfor SEQ ID NO: 1 eller 18. Den tunge kjeden av et antistoff ifølge oppfinnelsen kan omfatte en CDR1- sekvens av aminosyrene 31 til 3 (NYAMS) av SEQ ID NO: 18, hvori én av disse aminosyrene kan være erstattet av en ulik aminosyre. Den tunge kjeden av et antistoff ifølge oppfinnelsen kan omfatte en CDR2- sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) av SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være erstattet av en annen aminosyre. Den tunge kjeden av et antistoff ifølge oppfinnelsen kan omfatte en CDR3- sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) av SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være erstattet av en ulik aminosyre. Den lette kjeden av et antistoff ifølge oppfinnelsen kan omfatte en CDR1- sekvens av aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) av SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være erstattet av en annen aminosyre. Den lette kjeden av et antistoff ifølge oppfinnelsen kan omfatte en CDR2- sekvens av aminosyrene til 61 (LVSILDS) av SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være erstattet av en ulik aminosyre. Den lette kjeden av et antistoff ifølge oppfinnelsen kan omfatte en CDR3- sekvens av aminosyrene 94 til 2 (LQATHFPQT) av SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være erstattet av en annen aminosyre. Nærmere bestemt kan et antistoff ifølge oppfinnelsen ha en tung kjede som omfatter: en CDR1 sekvens som, i sin tur, omfatter aminosyrene 31 til 3 (NYAMS) av SEQ ID NO: 18; og en CDR2-sekvens som, i sin tur, omfatter aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) av SEQ ID NO: 18; og en CDR3-sekvens som, i sin tur, omfatter aminosyrene 99 til 1 (LGGYDEGDAMDS) av SEQ ID NO: 18. Et antistoff ifølge oppfinnelsen kan ha en lett kjede som omfatter: en CDR1-sekvens som, i sin tur, omfatter aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) av SEQ ID NO: 1; og en CDR2-sekvens som, i sin tur, omfatter aminosyrene til 61 (LVSILDS) av SEQ ID NO: 1; og en CDR3-sekvens som, i sin tur, omfatter aminosyrene 94 til 2 (LQATHFPQT) av SEQ ID NO: 1. Et antistoff ifølge oppfinnelsen kan omfatte hvilken som helst kombinasjon av de ovennevnte CDR-regionene.
35 Nærmere bestemt kan struktur ( framework )-region 2 (FR2) av den tunge kjeden av et antistoff ifølge oppfinnelsen omfatte aminosyrene: T, i posisjonen svarende til posisjon 40, E, i posisjonen svarende til posisjon 42, R, i posisjonen svarende til posisjon 44 og A, i posisjonen svarende til posisjon 49 av SEQ ID NO: 18. Alternativt kan nevnte FR2 av den tunge kjeden omfatte aminosyrene 36 til 49 av SEQ ID NO: 18. Et antistoff ifølge oppfinnelsen kan omfatte hvilke som helst av de følgende: VL-aminosyresekvensen ifølge SEQ ID NO: 1. VH-aminosyresekvensen av SEQ ID NO: 18. SEQ ID NO: 1 og 18. lett kjede aminosyresekvensen av SEQ ID NO: 21. tung kjede aminosyresekvensen av SEQ ID NO: 24. SEQ ID NO: 21 og 24. Et antistoff ifølge oppfinnelsen kan alternativt være eller kan omfatte en variant av én av disse spesifikke sekvensene slik som en variant av antistoffet MuTFPI4F36 eller en variant av HzTFPI4F36. For eksempel kan en variant være en substitusjon, delesjon eller addisjons-variant av hvilke som helst av de ovennevnte aminosyresekvensene. En variant i henhold til foreliggende oppfinnelse kan være et antistoff som ikke omfatter: N, i posisjonen svarende til posisjon 31 av CDR1-regionen av SEQ ID NO: 18; 3 R, i posisjonen svarende til posisjon 3; S, i posisjonen svarende til posisjon 4; S, i posisjonen svarende til posisjon 6; Y, i posisjonen svarende til posisjon 7; Y, i posisjonen svarende til posisjon 9; F, i posisjonen svarende til posisjon 60; P, i posisjonen svarende til posisjon 61; D, i posisjonen svarende til posisjon 62; og Q, i posisjonen svarende til posisjon 6; av CDR2-regionen av SEQ ID NO: 18.
36 34 Y, i posisjonen svarende til posisjon 2; D, i posisjonen svarende til posisjon 3; og D, i posisjonen svarende til posisjon 6; av CDR3-regionen av SEQ ID NO: 18. E, i posisjonen svarende til posisjon 31; S, i posisjonen svarende til posisjon 32; D, i posisjonen svarende til posisjon 33; og Y, i posisjonen svarende til posisjon 37; av CDR1-regionen av SEQ ID NO: 1. 1 A, i posisjonen svarende til posisjon 96; T, i posisjonen svarende til posisjon 97; H, i posisjonen svarende til posisjon 98; og F, i posisjonen svarende til posisjon 99; av CDR3-regionen av SEQ ID NO: Et variant antistoff kan omfatte 1, 2, 3, 4,, opp til, opp til, opp til eller flere aminosyresubstitusjoner og/eller delesjoner og/eller insersjoner fra de spesifikke sekvensene og fragmentene diskutert ovenfor. "Delesjons-"varianter kan omfatte delesjon av individuelle aminosyrer, delesjon av små grupper av aminosyrer slik som 2, 3, 4 eller aminosyrer, eller delesjon av større aminosyreregioner, slik som delesjon av spesifikke aminosyre-domener eller andre trekk. "Insersjons-" varianter kan omfatte insersjon av individuelle aminosyrer, insersjon av små grupper av aminosyrer slik som 2, 3, 4 eller aminosyrer, eller insersjon av større aminosyreregioner, slik som insersjon av spesifikke aminosyre-domener eller andre trekk. "Substitusjons"-varianter omfatter foretrukket å erstatte én eller flere aminosyrer med samme antall aminosyrer og foreta konservative aminosyresubstitusjoner. For eksempel kan en aminosyre erstattes av en alternativ aminosyre som har lignende egenskaper, for eksempel, en annen basisk aminosyre, en annen sur aminosyre, en annen nøytral aminosyre, en annen ladet aminosyre, en annen hydrofil aminosyre, en annen hydrofob aminosyre, en annen polar aminosyre, en annen aromatisk aminosyre eller en annen alifatisk aminosyre. Noen egenskaper av de viktigste aminosyrene som kan anvendes for å velge egnede substituenter er som følger: Ala alifatisk, hydrofob, nøytral Met hydrofob, nøytral
37 3 Cys polar, hydrofob, nøytral Asn polar, hydrofil, nøytral Asp polar, hydrofil, ladet (-) Pro hydrofob, nøytral Glu polar, hydrofil, ladet (-) Gln polar, hydrofil, nøytral Phe aromatisk, hydrofob, nøytral Arg polar, hydrofil, ladet (+) Gly alifatisk, nøytral Ser polar, hydrofil, nøytral His aromatisk, polar, hydrofil, ladet (+) Thr polar, hydrofil, nøytral Ile alifatisk, hydrofob, nøytral Val alifatisk, hydrofob, nøytral Lys polar, hydrofil, ladet (+) Trp aromatisk, hydrofob, nøytral Leu alifatisk, hydrofob, nøytral Tyr aromatisk, polar, hydrofob 1 2 Foretrukne "derivater" eller "varianter" omfatter slike som i hvilke aminosyren som finnes i sekvensen, i stedet for den naturlig forekommende aminosyren, er en strukturell analog derav. Aminosyrer anvendt i sekvensene kan også være derivatisert eller modifisert, f.eks. merket, under forutsetning av at funksjonen av antistoffet ikke blir betydelig negativt påvirket. Substitusjoner kan være, men er ikke begrenset til, konservative substitusjoner. Derivater og varianter som beskrevet ovenfor kan fremstilles under syntese av antistoffet eller ved modifikasjon etter produksjon, eller når antistoffet er i rekombinant form ved anvendelse av de kjente teknikkene for seterettet mutagenese, tilfeldig mutagenese, eller enzymatisk kløyving og/eller ligering av nukleinsyrer. I et annet aspekt viser foreliggende oppfinnelse multispesifikke molekyler omfattende et anti-tfpi antistoff, eller et antigen-fragment derav, ifølge oppfinnelsen. Slike multispesifikke molekyler omfatter bispesifikke molekyler omfattende minst én første bindingsspesifisitet for TFPI og en andre bindingsspesifisitet for en andre mål-epitop. En type bispesifikke molekyler er bispesifikke antistoffer som kjent på området. Bispesifikke antistoffer, eller faktisk multispesifikke antistoffer, kan fremstilles som fullengde antistoffer eller antistoffragmenter (f.eks. F(ab')2 bispesifikke antistoffer) eller hvilke som helst andre antigen-bindende fragmenter beskrevet heri. I ett aspekt viser foreliggende oppfinnelse antistoffderivater (eller immunkonjugater), slik som anfi-tfpi antistoffer konjugert eller kovalent bundet til et andre middel. Det andre midlet kan være forbundet med antistoffet direkte eller indirekte, ved anvendelse av hvilke som helst av et stort antall tilgjengelige metoder kjent for fagfolk på området. For eksempel kan et middel være bundet ved hengselsregionen av den reduserte antistoff-komponenten via dannelse av disulfidbinding, ved anvendelse av kryssbindere slik som N- succinyl S-(2-
38 pyridylditio) proprionat (SPDP), eller via en karbohydratgruppe i Fc-regionen av antistoffet. I ett aspekt kan antistoffer ifølge oppfinnelsen konstrueres til å omfatte modifikasjoner i Fc-regionen, typisk for å endre én eller flere funksjonelle egenskaper av antistoffet, slik som halveringstid i serum, komplementfiksering, Fcreseptorbinding, proteinstabilitet og/eller antigen-avhengig cellulær cytotoksisitet, eller mangel derav. Videre kan et antistoff ifølge oppfinnelsen være kjemisk modifisert (f.eks., én eller flere kjemiske grupper kan være bundet til antistoffet) eller være modifisert for å endre dets glykosylering, igjen for å endre én eller flere funksjonelle egenskaper av antistoffet. Om ønsket kan klassen av et antistoff "switches" ved kjente teknikker. For eksempel kan et antistoff som opprinnelig ble produsert som et IgM-molekyl underlegges klasse-switching for å danne et IgG-antistoff. Teknikker for klasse switching kan også anvendes for å omdanne én IgG subklasse til en annen, for eksempel: fra IgG1 til IgG2 eller IgG4; fra IgG2 til IgG1 eller IgG4; eller fra IgG4 til IgG1 eller IgG2. Konstruksjon av antistoffer for å danne konstant region kimære molekyler, ved kombinasjon av regioner fra forskjellige IgG subklasser, kan også foretas. I én utførelsesform er hengselsregionen av CHI modifisert slik at antall cysteinrester i hengselsregionen er endret, f.eks., økt eller redusert. Denne metoden er beskrevet ytterligere for eksempel i U.S. Patent Nr.,677,42 ved Bodmer et al. Den konstante regionen kan videre modifiseres for å stabilisere antistoffet, f.eks., for å redusere risikoen for at et bivalent antistoff deles opp til to monovalent VH-VL fragmenter. I en lgg4 konstant region kan for eksempel rest S241 muteres til en prolin (P)-rest for å muliggjøre komplett disulfidbro-dannelse ved hengselen (se, f.eks., Angal et al., Mol Immunol. 199S; : -8). Variant antistoffer ifølge oppfinnelsen kan ha aminosyresekvenser som er mer enn 60 %, eller mer enn 6 %, eller mer enn 70 %, eller mer enn 7 %, eller mer enn 80 %, foretrukket mer enn 8 %, slik som mer enn 90 %, slik som mer enn 9 % identisk med SEQ ID NO: 4 eller 8, eller fragmenter derav. Andre variant antistoffer i henhold til oppfinnelsen kan ha aminosyresekvenser som er mer enn 60 %, eller mer enn 6 %, eller mer enn 70 %, eller mer enn 7 %, eller mer enn 80 %, foretrukket mer enn 8 %, slik som mer enn 90 %, slik som mer enn 9 % identisk med SEQ ID NO: 1 eller 18, eller et fragment derav. Dette nivået av aminosyreidentitet kan ses gjennom hele den fulle lengden av den relevante SEQ ID NO-sekvensen eller over en del av sekvensen, slik som over,, 40, 0, 60, 70,
39 37 1 7, 80, 90, 0,, 0 eller flere aminosyrer, avhengig av størrelsen av fullengde polypeptidet. I forbindelse med aminosyresekvenser angir "sekvensidentitet" sekvenser som har den angitte verdien når vurdert ved anvendelse av ClustalW (Thompson et al., 1994, supra) med følgende parametere: Parvis sammenstillingsparametere -Metode: nøyaktig, Matrise: PAM, Gap åpne straff:.00, Gap utvidelse straff: 0.; Multiple sammenstillings parametere -Matrise: PAM, Gap åpne straff:.00, % identitet for forsinkelse:, Straff ende gaps: på, Gap separasjon avstand: 0, Negativ matrise: nei, Gap utvidelse straff: 0., Rest-spesifikk gap straff: på, Hydrofil gap straff: på, Hydrofile rester: GPSNDQEKR. Sekvensidentitet ved en bestemt rest er ment å inkludere identiske rester som ganske enkelt har blitt derivatisert. Foreliggende oppfinnelse tilveiebringer følgelig antistoffer som har spesifikke VH og VL-aminosyresekvenser og varianter og fragmenter derav som opprettholder funksjonen eller aktiviteten av disse VH og VL-domenene. Følgelig kan et antistoff ifølge oppfinnelsen omfatte: (a) en lett kjede variabel region aminosyresekvens fra SEQ ID NO: 4; (b) et fragment på minst 7 aminosyrer av (a) som bibeholder evnen til å spesifikt binde til TFPI; eller (c) en variant av (a) med minst 70 % aminosyresekvensidentitet med en sekvens ifølge (a) og bibeholder evnen til å spesifikt binde til TFPI. 2 Et antistoff ifølge oppfinnelsen kan omfatte: (a) en tung kjede variabel region aminosyresekvens av SEQ ID NO: 8; (b) et fragment på minst 7 aminosyrer ifølge (a) som bibeholder evnen til å spesifikt binde til TFPI; eller (c) en variant av (a) med minst 70 % aminosyresekvensidentitet med en sekvens ifølge (a) og bibeholder evne til å spesifikt binde til TFPI. Et antistoff ifølge oppfinnelsen kan omfatte lett kjede variabel region av SEQ ID NO: 4 og tung kjede variabel region av SEQ ID NO: 8. 3 Et antistoff ifølge oppfinnelsen kan omfatte: (a) lett kjede variabel region fra SEQ ID NO: 4 og tung kjede variabel region fra SEQ ID NO: 8; (b) en variant av (a) i hvilken én eller begge av tung kjede og lett kjedesekvensene er modifisert slik at de omfatter et fragment på minst 7 aminosyrer av sekvensen angitt i (a); eller
40 (c) en variant av (a) eller (b) i hvilken én eller begge av tung og lett kjede sekvensene er modifisert slik at de har minst 70 % aminosyresekvensidentitet med en sekvens ifølge (a) eller (b); hvori antistoffet bibeholder evnen til å spesifikt binde til TFPI. Antistoffet kan også bibeholde én eller flere ytterligere funksjoner eller aktiviteter av et antistoff ifølge oppfinnelsen som beskrevet heri slik som evne til å inhibere TFPI eller evne til å forkorte koagulasjonstid, eventuelt uten at det fører til at blodplatetallet synker. Foretrukne fragmenter og varianter av SEQ ID NO: 4 vil omfatte (i) aminosyrene 24 til 39 fra SEQ ID NO: 6; og/eller (ii) aminosyrene til 61 fra SEQ ID NO: 6; og/eller (iii) aminosyrene 94 til 2 fra SEQ ID NO: 6. Foretrukne fragmenter og varianter av SEQ ID NO: 8 vil omfatte (i) aminosyrene 31 til 3 fra SEQ ID NO: ; og/eller (ii) aminosyrene 0 til 66 fra SEQ ID NO: ; og/eller (iii) aminosyrene 99 til 1 fra SEQ ID NO:. Ytterligere foretrukne varianter av SEQ ID NO: 4 vil omfatte aminosyrene 31 til 33, 37 og 96 til 99 fra SEQ ID NO: 6. Ytterligere foretrukne varianter av SEQ ID NO: 8 vil omfatte aminosyrene 31, 3, 4, 6, 7, 9, 60, 61, 62, 6, 2, 3 og 6 fra SEQ ID NO:. Et antistoff ifølge oppfinnelsen kan omfatte: (a) en lett kjede variabel region aminosyresekvens fra SEQ ID NO: 1; (b) et fragment på minst 7 aminosyrer fra (a) som bibeholder evne til å spesifikt binde til TFPI; eller (c) en variant av (a) som har minst 70 % aminosyresekvensidentitet med en sekvens ifølge (a) og bibeholder evne til å spesifikt binde til TFPI. Et antistoff ifølge oppfinnelsen kan omfatte: (a) en tung kjede variabel region aminosyresekvens fra SEQ ID NO: 18; (b) et fragment på minst 7 aminosyrer ifølge (a) som bibeholder evne til å spesifikt binde til TFPI; eller (c) en variant av (a) som har minst 70 % aminosyresekvensidentitet med en sekvens ifølge (a) og bibeholder evne til å spesifikt binde til TFPI. Et antistoff ifølge oppfinnelsen kan omfatte lett kjede variabel region fra SEQ ID NO: 1 og tung kjede variabel region fra SEQ ID NO: 18. Et antistoff ifølge oppfinnelsen kan omfatte: (a) lett kjede variabel region fra SEQ ID NO: 1 og tung kjede variabel region fra SEQ ID NO: 18; (b) en variant av (a) hvilken én eller begge av tung kjede og lett kjedesekvensene er modifisert slik at de omfatter et fragment på minst 7 aminosyrer av sekvensen angitt i (a); eller
41 (c) en variant av (a) eller (b) hvilken én eller begge av tung og lett kjedesekvensene er modifisert slik at de har minst 70 % aminosyresekvensidentitet med en sekvens ifølge (a) eller (b); hvori antistoffet bibeholder evne til å spesifikt binde til TFPI. Antistoffet kan også bibeholde én eller flere ytterligere funksjoner eller aktiviteter av et antistoff ifølge oppfinnelsen som beskrevet heri slik som evne til å inhibere TFPI eller evne til å forkorte koagulasjonstid, eventuelt uten at det fører til at blodplatetallet synker. Som forklart ovenfor kan et antistoff ifølge oppfinnelsen binde til samme epitop eller region som et annet antistoff ifølge oppfinnelsen. Følgelig vil det ses at et slikt antistoff kan binde til samme epitop eller region av TFPI som hvilke som helst av de spesifikke antistoffene, fragmentene og variantene beskrevet heri. Oppfinnelsen også angår polynukleotider som koder for antistoffer ifølge oppfinnelsen. Følgelig kan et polynukleotid ifølge oppfinnelsen kode for hvilket som helst antistoff som beskrevet heri. Betegnelsene "nukleinsyremolekyl" og "polynukleotid" blir anvendt om hverandre heri og refererer til en polymer form av nukleotider av hvilken som helst lengde, enten deoksyribonukleotider eller ribonukleotider, eller analoger derav. Ikke-begrensende eksempler på polynukleotider omfatter et gen, et genfragment, messenger RNA (mrna), cdna, rekombinante polynukleotider, plasmider, vektorer, isolert DNA av hvilken som helst sekvens, isolert RNA av hvilken som helst sekvens, nukleinsyreprober og primere. Et polynukleotid ifølge oppfinnelsen kan tilveiebringes i isolert eller renset form. En nukleinsyresekvens som "koder for" et valgt polypeptid er et nukleinsyremolekyl som blir transkribert (i tilfelle DNA) og translatert (i tilfelle mrna) til et polypeptid in vivo når den blir plassert under kontroll av egnede regulatoriske sekvenser. Grensene for den kodende sekvensen bestemmes av et startkodon ved den ' (amino) terminale enden og et translasjonsstopp-kodon ved 3' (karboksy)- enden. For formålene ifølge oppfinnelsen, kan slike nukleinsyresekvenser omfatte, men er ikke begrenset til, cdna fra viralt, prokaryot eller eukaryot mrna, genomiske sekvenser fra viralt eller prokaryot DNA eller RNA, og til og med syntetiske DNA-sekvenser. En transkripsjonstermineringssekvens kan være lokalisert 3' for den kodende sekvensen. I én utførelsesform omfatter et polynukleotid ifølge oppfinnelsen en sekvens som koder for en VH eller VL aminosyresekvens som beskrevet ovenfor. For eksempel kan et polynukleotid ifølge oppfinnelsen kode for et polypeptid omfattende sekvensen ifølge SEQ ID NO: 4 eller 8, eller en variant eller fragment derav som beskrevet ovenfor. Et slikt polynukleotid kan bestå av eller omfatte en nukleinsyresekvens fra hvilken som helst av SEQ ID NO: 3,, 7 og 9. En egnet polynukleotidsekvens kan alternativt være en variant av én av disse spesifikke
42 polynukleotidsekvensene. For eksempel kan en variant være en substitusjon, delesjon eller addisjonsvariant av hvilke som helst av de ovennevnte nukleinsyresekvensene. Et variant polynukleotid kan omfatte 1, 2, 3, 4,, opp til, opp til, opp til, opp til 40, opp til 0, opp til 7 eller flere nukleinsyresubstitusjoner og/eller delesjoner fra sekvensene gitt i sekvenslisten. Egnede varianter kan være minst 70 % homologe med et polynukleotid ifølge hvilken som helst av SEQ ID NO: 3,, 7 og 9 foretrukket minst 80 eller 90 % og mer foretrukket minst 9 %, 97 % eller 99 % homolog dertil. Metoder for måling av homologi er velkjent på området og det vil forstås av fagfolk på området at i foreliggende kontekst, blir homologi beregnet på grunnlag av nukleinsyreidentitet. Slik homologi kan eksistere over en region på minst 1, foretrukket minst, for eksempel minst 40, 60, 0, 0 eller flere sammenhengende nukleotider. Slik homologi kan eksistere over hele lengden av den umodifiserte polynukleotidsekvensen. Metoder for måling av homologi eller identitet av polynukleotider er kjent på området. For eksempel tilveiebringer UWGCG Package BESTFIT-programmet som kan anvendes for å beregne homologi (f.eks. anvendt ved dets standard innstillinger) (Devereux et al (1984) Nucleic Acids Research 12, s ). PILEUP og BLAST-algoritmene kan også anvendes for å beregne homologi eller stille opp sekvenser (typisk ved deres standard innstillinger), for eksempel som beskrevet i Altschul S. F. (1993) J MoI Evol 36: 290-0; Altschul, S, F et al (1990) J MoI Biol 21:403-. Programvare for utførelse av BLAST-analyser er offentlig tilgjengelig gjennom National Centre for Biotechnology Information ( Denne algoritmen omfatter først å identifisere sekvenspar med høyt score (HSP) ved å identifisere korte ord med lengde W i spørre ("query")-sekvensen som enten matcher eller tilfredsstiller en eller annen positivt-vurdert terskel-score T ved sammenstilling med et ord av samme lengde i en databasesekvens. T blir referert til som naboskap ("neighbourhood") ord score-terskelen (Altschul et al, supra). Disse initielle naboskap ord-treffene fungerer som såkorn for initiering av søk for å finne HSP'er som inneholder dem. Ord-treffene utvides i begge retninger langs hver sekvens for så langt som det kumulative sammenstilling-scoret kan økes. Utvidelser for ord-treffene i hver retning blir stanset når: det kumulative sammenstillings-score går til null eller lavere, beroende på akkumulering av én eller flere negativt-scorende rest sammenstillinger; eller enden av den ene eller andre sekvensen er nådd. BLAST algoritme-parametrene W, T og X bestemmer sensitiviteten og hastigheten av sammenstillingen. BLAST-programmet anvender som standarder en ordlengde (W) på 11, BLOSUM62 scoringsmatrisen (se Henikoff og Henikoff (1992) Proc. Natl. Acad.
43 Sci. USA 89: ) sammenstillinger (B) på 0, forventning ("expectation") (E) på, M =, N=4, og en sammenligning av begge trådene. BLAST-algoritmen foretar en en statistisk analyse av likheten mellom to sekvenser; se f.eks., Karlin og Altschul (1993) Proc. Natl. Acad. Sci. USA 90: Ett mål for likheten gitt ved BLAST-algoritmen er den minste sum sannsynlighet (P(N)), som gir en indikasjon på sannsynligheten ved hvilken en match mellom to nukleotid- eller aminosyresekvenser ville forekomme tilfeldig. For eksempel bli en sekvens ansett for å være lik en annen sekvens dersom den minste sum sannsynlighet ved sammenligning av den første sekvensen med den andre sekvensen er mindre enn omtrent 1, foretrukket mindre enn omtrent 0,1, mer foretrukket mindre enn omtrent 0,01, og mest foretrukket mindre enn omtrent 0,001. En homolog sekvens kan skille seg fra en sekvens i det relevante polynukleotidet ved mindre enn 3,,, 1, eller flere mutasjoner (som hver kan være en substitusjon, delesjon eller insersjon). Disse mutasjonene kan måles over en region på minst, for eksempel minst 40, 60 eller 0 eller flere sammenhengende nukleotider av den homologe sekvensen. I én utførelsesform kan en variant sekvens variere fra de spesifikke sekvensene gitt i sekvenslisten i kraft av redundansen for den genetiske koden. DNA-koden har 4 primære nukleinsyrerester (A, T, C og G) og anvender disse til å "stave" trebokstavs kodoner som representerer aminosyrene av proteinene som kodes for i en organismes gener. Den lineære sekvensen av kodoner langs DNAmolekylet blir translatert til den lineære sekvensen av aminosyrer i proteinet(/proteinene) kodet for av disse genene. Koden er høyst degenerert, med 61 kodoner som koder for de naturlige aminosyrene og 3 kodoner som representerer "stopp"-signaler. Følgelig blir de fleste aminosyrer kodet for av mer enn ett kodon - faktisk blir flere kodet for av fire eller flere forskjellige kodoner. Et variant polynukleotid ifølge oppfinnelsen kan derfor kode for den samme polypeptidsekvensen som et annet polynukleotid ifølge oppfinnelsen, men kan ha en ulik nukleinsyresekvens på grunn av bruk av ulike kodoner til å kode for de samme aminosyrene. Polynukleotid-"fragmenter" i henhold til oppfinnelsen kan fremstilles ved trunkering, f.eks. ved å fjerne ett eller flere nukleotider fra én eller begge ender av et polynukleotid. Opp til, opp til, opp til, opp til 40, opp til 0, opp til 7, opp til 0, opp til 0 eller flere aminosyrer kan fjernes fra 3'- og/eller '-enden av polynukleotidet på denne måten. Fragmenter kan også genereres ved én eller flere interne delesjoner. Slike fragmenter kan være avledet fra en sekvens ifølge SEQ ID NO: 3,, 7 og 9 eller kan være avledet fra en variant polynukleotid som beskrevet
44 heri. Foretrukket har slike fragmenter en lengde på mellom og 0 rester, f.eks. til 0, til 0, til 0, 0 til 0 eller 0 til 0 rester. Alternativt kan fragmenter ifølge oppfinnelsen være lengre sekvenser, for eksempel omfattende minst 0 %, minst 60 %, minst 70 %, minst 80 % eller minst 90 % av et fullengde polynukleotid ifølge oppfinnelsen. Et antistoff ifølge oppfinnelsen kan følgelig bli fremstilt fra eller levert i form av et polynukleotid som koder for, og er i stand til å uttrykke, det. Når antistoffet omfatter to eller flere kjeder, kan et polynukleotid ifølge oppfinnelsen kode for ett eller flere antistoffkjeder. For eksempel kan et polynukleotid ifølge oppfinnelsen kode for en lett kjede av et antistoff, en tung kjede av et antistoff eller begge. To polynukleotider kan tilveiebringes, av hvilke ett koder for en lett kjede av et antistoff og av hvilke det andre koder for den korresponderende tunge kjeden av et antistoff. Et slikt polynukleotid eller par av polynukleotider kan uttrykkes sammen slik at et antistoff ifølge oppfinnelsen blir fremstilt. Polynukleotider ifølge oppfinnelsen kan syntetiseres i henhold til metoder velkjent på området, som beskrevet som eksempel i Sambrook et al (1989, Molecular Cloning - a laboratory manual; Cold Spring Harbor Press). Nukleinsyremolekyler ifølge foreliggende oppfinnelse kan tilveiebringes i form av en ekspresjonskassett som omfatter kontrollsekvenser, signalpeptidsekvenser operabelt bundet til den inserterte sekvensen, hvilket følgelig muliggjør ekspresjon av antistoffet ifølge oppfinnelsen in vivo. Disse ekspresjonskassettene blir, i sin tur, typisk tilveiebrakt i vektorer (f.eks., plasmider eller rekombinante virale vektorer). En slik ekspresjons-kassett kan administreres direkte til et verts-subjekt. Alternativt kan en vektor omfattende et polynukleotid ifølge oppfinnelsen administreres til et verts-subjekt. Foretrukket blir polynukleotidet fremstilt og/eller administrert ved anvendelse av en genetisk vektor. En egnet vektor kan være hvilken som helst vektor som er i stand til å romme en tilstrekkelig mengde genetisk informasjon, og tillate ekspresjon av et polypeptid ifølge oppfinnelsen. Foreliggende oppfinnelse omfatter følgelig ekspresjonsvektorer som omfatter slike polynukleotidsekvenser. Slike ekspresjonsvektorer blir rutinemessig konstruert innenfor området molekylærbiologi og kan for eksempel omfatte anvendelse av plasmid-dna og passende initiatorer, promotere, enhancere, signalpeptidsekvenser og andre elementer, slik som for eksempel polyadenyleringssignaler som kan være nødvendig, og som er plassert i riktig orientering, for å muliggjøre ekspresjon av et peptid ifølge oppfinnelsen. Andre egnede vektorer ville være innlysende for fagfolk på området. Som ytterligere esempel i dette henseende viser vi til Sambrook et al. Oppfinnelsen omfatter også celler som har blitt modifisert til å uttrykke et antistoff ifølge oppfinnelsen. Slike celler omfatter transiente, eller foretrukket stabile
45 høyere eukaryote cellelinjer, slik som mammalske celler eller insektceller, lavere eukaryote celler, slik som gjær eller prokaryote celler slik som bakterieceller. Spesielle eksempler på celler som kan modifiseres ved insersjon av vektorer eller ekspresjonskassetter som koder for et antistoff ifølge oppfinnelsen omfatter mammalsk HEK293, CHO, BHK, NSO og human retinaceller. Foretrukket vil cellelinjen valgt være en som ikke bare er stabil, men også tillater moden glykosylering og celleoverflateekspresjon av et polypeptid. Slike cellelinjer ifølge oppfinnelsen kan dyrkes ved anvendelse av rutinemessige metoder for å fremstille et antistoff ifølge oppfinnelsen, eller kan anvendes terapeutisk eller profylaktisk til å levere antistoffer ifølge oppfinnelsen til et subjekt. Alternativt kan polynukleotider, ekspresjonskassetter eller vektorer ifølge oppfinnelsen administreres til en celle fra et subjekt ex vivo og cellen deretter returneres til kroppen til subjektet. I et annet aspekt tilveiebringer foreliggende oppfinnelse preparater og formuleringer omfattende molekyler ifølge oppfinnelsen, slik som antistoffene, polynukleotidene, vektorene og cellene beskrevet heri. For eksempel tilveiebringer oppfinnelsen et farmasøytisk preparat omfattende ett eller flere molekyler ifølge oppfinnelsen, slik som ett eller flere antistoffer ifølge oppfinnelsen, formulert sammen med en farmasøytisk akseptabel bærer. Som anvendt heri omfatter "farmasøytisk akseptabel bærer" hvilke som helst og alle løsningsmidler, dispersjonsmedier, drasjeringer, antibakterielle og antifungale midler, isotoniske og absorpsjonsforsinkende midler, og lignende som er fysiologisk kompatible. Foretrukket er bæreren egnet for parenteral, f.eks. intravenøs, intramuskulær eller subkutan administrering (f.eks., ved injeksjon eller infusjon). Avhengig av administreringsveien, kan antistoffet bli overtrukket i et materiale for å beskytte antistoffet mot virkningen av syrer og andre naturlige betingelser som kan inaktivere eller denaturere antistoffet. Foretrukne farmasøytisk akseptable bærere omfatter vandige bærere eller fortynningsmidler. Eksempler på egnede vandige bærere som kan anvendes i de farmasøytiske preparatene ifølge oppfinnelsen omfatter vann, bufret vann og saltoppløsning. Eksempler på andre bærere omfatter etanol, polyoler (slik som glyserol, propylenglykol, polyetylenglykol, og lignende), og passende blandinger derav, vegetabilske oljer, slik som olivenolje, og injiserbare organiske estere, slik som etyloleat. Riktig fluiditet kan opprettholdes, for eksempel, ved anvendelse av drasjeringsmaterialer, slik som lecitin, ved å opprettholde den nødvendige partikkelstørrelsen i tilfelle av dispersjoner, og ved anvendelse av surfaktanter. I mange tilfeller vil det være foretrukket å inkludere isotoniske midler, for eksempel, sukker, polyalkoholer slik som mannitol, sorbitol, eller natriumklorid i preparatet.
46 Et farmasøytisk preparat ifølge oppfinnelsen kan også omfatte en farmasøytisk akseptabel antioksidant. Disse preparatene kan også inneholde adjuvanser slik som konserveringsmidler, fuktemidler, emulgeringsmidler og dispergeringsmidler. Forhindring av tilstedeværelse av mikroorganismer kan ivaretas både ved steriliseringsmetoder, ovenfor, og ved inkludering av forskjellige antibakterielle og antifungale midler, for eksempel, paraben, klorbutanol, fenol sorbinsyre, og lignende. Det kan også være ønskelig å inkludere isotoniske midler, slik som sukker, natriumklorid, og lignende i preparatene. I tillegg kan forlenget absorpsjon av den injiserbare farmasøytiske formen fås i stand ved å inkludere midler som forsinker absorpsjon slik som aluminum monostearat og gelatin. Terapeutiske preparater må typisk være sterile og stabile under betingelsene for fremstilling og lagring. Preparatet kan formuleres som en løsning, mikroemulsjon, liposom, eller annen ordinert struktur egnet for høy medikamentkonsentrasjon. Sterile injiserbare løsninger kan fremstilles ved å innarbeide det aktive midlet (f.eks. antistoff) i den nødvendige mengden i et egnet løsningsmiddel med én eller en kombinasjon av ingredienser spesifisert ovenfor, som nødvendig, fulgt av sterilisering mikrofiltrering. Generelt blir dispersjoner fremstilt ved å innarbeide det aktive midlet i en steril konstituent som inneholder et basisk dispersjonsmedium og de andre nødvendige ingrediensene fra de spesifisert ovenfor. I tilfelle av sterile pulvere for fremstilling av sterile injiserbare løsninger, er de foretrukne metodene for fremstilling vakuumtørking og frysetørking (lyofilisering) som gir et pulver av det aktive midlet pluss hvilket som helst ytterligere ønsket ingrediens fra en tidligere sterilfiltrert løsning derav. Farmasøytiske preparater ifølge oppfinnelsen kan omfatte ytterligere aktive ingredienser så vel som et antistoff ifølge oppfinnelsen. Som nevnt ovenfor kan preparater ifølge oppfinnelsen omfatte ett eller flere antistoffer ifølge oppfinnelsen. De kan også omfatte ytterligere terapeutiske eller profylaktisk midler. For eksempel når et farmasøytisk preparat ifølge oppfinnelsen er ment for anvendelse ved behandling av en blødningsforstyrrelse, kan det ytterligere omfatte ett eller flere midler ment å redusere symptomene på blødningsforstyrrelsen. For eksempel kan preparatet omfatte én eller flere koagulasjonsfaktorer. Preparatet kan omfatte én eller flere andre komponenter ment å forbedre tilstanden til pasienten. Når for eksempel preparatet er ment for anvendelse ved behandling av pasienter som lider av uønsket blødning slik som pasienter som gjennomgår kirurgi eller pasienter som lider av traume, kan preparatet omfatte ett eller flere analgetika, anestetika, immunosuppressive stoffer eller antiinflammatoriske midler. Også innenfor omfanget av foreliggende oppfinnelse er sett omfattende antistoffer eller andre preparater
47 ifølge oppfinnelsen og instruksjoner for anvendelse. Et slikt sett kan videre inneholde én eller flere ytterligere reagenser, slik som et ytterligere terapeutisk eller profylaktisk middel som diskutert ovenfor. Antistoffene, andre molekyler og preparater ifølge foreliggende oppfinnelse har mange in vitro og in vivo terapeutiske anvendbarheter omfattende behandling og forebygging av koagulasjonsrelaterte forstyrrelser. For eksempel kan disse antistoffene og preparatene administreres til celler i kultur, in vitro eller ex vivo, eller til humane subjekter, f.eks., in vivo, for å forebygge eller behandle mange forskjellige forstyrrelser. Spesielt tilveiebringer foreliggende oppfinnelse fremgangsmåter for behandling av blødningsforstyrrelse eller for forøkelse av blodkoagulasjon omfattende å administere til en pasient med behov derfor en effektiv mengde av et antistoff eller annet molekyl eller preparat ifølge oppfinnelsen. For eksempel kan slike fremgangsmåter være for behandling av mangel på koagulasjonsfaktor slik som hemofili A, hemofili B, Faktor XI-mangel, Faktor VII-mangel, trombocytopeni eller von Willebrands sykdom. Slike fremgangsmåter kan være for behandling av tilstander forbundet med nærvær av en koagulasjonsfaktor-inhibitor. Slike fremgangsmåter kan være for behandling av overdreven blødning. Antistoffene og preparatene ifølge oppfinnelsen kan anvendes for å behandle pasienter før, under, eller etter kirurgi eller antikoagulant behandling eller etter traume. Antistoffene og preparatene beskrevet heri kan anvendes ved hvilken som helst slik behandling eller kan anvendes ved fremstilling av et medikament for anvendelse i hvilken som helst slik behandling. Antistoffene og preparatene ifølge foreliggende oppfinnelse kan administreres for profylaktisk/forbyggende og/eller terapeutiske behandlinger. Ved terapeutiske anvendelser blir antistoffer eller preparater administrert til et subjekt som allerede lider av en forstyrrelse eller lidelse som beskrevet ovenfor, i en mengde tilstrekkelig til å kurere, lindre eller delvis stanse lidelsen eller ett eller flere av dens symptomer. Slik terapeutisk behandling kan føre til en reduksjon i alvorlighets-graden av sykdomssymptomer, eller en økning i hyppighet eller varighet av symptomfrie perioder. En mengde passende til å oppnå dette er definert som "terapeutisk effektiv mengde". For eksempel når behandlingen er for uønsket blødning, kan behandling defineres som en reduksjon i mengden av blødning eller passende koagulasjon for å stanse blødningen fullstendig. Ved profylaktiske eller forebyggende anvendelser, blir antistoffer eller preparater administrert til et subjekt med risiko for en forstyrrelse eller lidelse som beskrevet ovenfor, i en mengde tilstrekkelig til å forebygge eller redusere de senere effektene av lidelsen eller ett eller flere av dens symptomer. En mengde passende til
48 å oppnå dette er definert som en "profylaktisk effektiv mengde". For eksempel når behandlingrn er for å forhindre uønsket blødning, kan en profylaktisk effekt defineres som forebygging av blødning eller en redusert periode eller mengde av blødning sammenlignet med den som ville blitt sett i fravær av modulatoren. Effektive mengder for hvert formål vil avhenge av alvorlighetsgraden av sykdommen eller skaden så vel som vekt og generell tilstand av subjektet. Som anvendt heri omfatter betegnelsen "subjekt" hvilket som helst menneske eller ikke-humant dyr. Betegnelsen "ikke-humant dyr" omfatter alle virveldyr, f.eks., pattedyr og ikke-pattedyr, slik som ikke-humane primater, sauer, hunder, katter, hester, kyr, kyllinger, amfibier, reptiler, osv. Et antistoff eller preparat ifølge foreliggende oppfinnelse kan administreres via én eller flere administreringsveier ved anvendelse av én eller flere av mange forskjellige metoder kjent på området. Hvilket vil være forstått av fagfolk på området, vil administreringsveien og/eller administreringsmetoden variere avhengig av de ønskede resultatene. Foretrukne administreringsveier for antistoffer eller preparater ifølge oppfinnelsen omfatter intravenøs, intramuskulær, intradermal, intraperitoneal, subkutan, spinal eller andre parenterale administreringsveier, for eksempel ved injeksjon eller infusjon. Uttrykket "parenteral administrering" som anvendt heri betyr metoder for administrering andre enn enteral og topikal administrering, vanligvis ved injeksjon. Alternativt kan et antistoff eller preparat ifølge oppfinnelsen administreres via en ikke-parenteral administreringsvei, slik som en topikal, epidermal eller mukosal administreringsvei. På lignende måte kan et antistoff ifølge oppfinnelsen anvendes for fremstilling av et medikament egnet for parenteral administrering. Et antistoff ifølge oppfinnelsen kan anvendes for fremstilling av et medikament egnet for intravenøs administrering. Et antistoff ifølge oppfinnelsen kan anvendes for fremstilling av et medikament egnet for intramuskulær administrering. Et antistoff ifølge oppfinnelsen kan anvendes for fremstilling av et medikament egnet for subkutan administrering. En passende dose av et antistoff ifølge oppfinnelsen kan bestemmes av en praktiserende lege. Reelle doseringsnivåer av de aktive ingrediensene i de farmasøytiske preparatene ifølge foreliggende oppfinnelse kan varieres for å oppnå en mengde av den aktive ingrediensen som er effektiv for å oppnå den ønskede terapeutiske responsen for en bestemt pasient, preparat, og metode for administrering, uten å være toksisk for pasienten. Det valgte doseringsnivået vil avhenge av mange forskjellige farmakokinetiske faktorer inkludert aktiviteten av de bestemte antistoffet anvendt, administreringsveien, tiden for administrering,
49 ekskresjonsraten for antistoffet, varigheten av behandlingen, andre medikamenter, forbindelser og/eller materialer anvendt i kombinasjon med de bestemte preparatene anvendt, alder, kjønn, vekt, tilstand, generell helse og tidligere medisinsk historie for pasienten som behandles, og lignende faktorer velkjent på det medisinske området. En passende dose av et antistoff ifølge oppfinnelsen kan være, for eksempel, i området fra omtrent 0,1 μg/kg til omtrent 0 mg/kg kroppsvekt av pasienten som skal behandles. For eksempel kan en passende dose være fra omtrent 1 μg/kg til omtrent mg/kg kroppsvekt per dag eller fra omtrent 1 mg/kg til omtrent mg/kg kroppsvekt per dag. En passende dose av et antistoff ifølge oppfinnelsen kan være i området fra 2 til 0 mg/kg, slik som omtrent -0 mg/kg, slik som omtrent -170 mg/kg, slik som omtrent 0- mg/kg, slik som omtrent 0-0 mg/kg, slik som omtrent mg/kg, slik som omtrent -0 mg/kg, slik som omtrent - mg/kg. Andre passende doser kan være omtrent 0,1- mg/kg, slik som omtrent 0,1-1 mg/kg, slik som omtrent 1-2 mg/kg eller omtrent 2-3 mg/kg eller omtrent 4- mg/kg eller omtrent -6 mg/kg eller omtrent 6-7 mg/kg eller omtrent 7-8 mg/kg eller omtrent 8-9 mg/kg eller omtrent 9- mg/kg; eller omtrent -21 mg/kg, slik som omtrent -11 mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent 14-1 mg/kg, eller omtrent 1-16 mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent 19- mg/kg eller omtrent -21 mg/kg. Mengden av monoklonalt antistoff administrert til et subjekt kan være slik at dets administrering fører til en plasmakonsentrasjon hos subjektet på omtrent μg/ml til omtrent 40 μg/ml, slik som omtrent 1-3 μg/ml, slik som omtrent -1 μg/ml, slik som omtrent 1- μg/ml, slik som omtrent -2 μg/ml, slik som omtrent 2- μg/ml, slik som omtrent -3 μg/ml, slik som omtrent 3-40 μg/ml, av nevnte monoklonale antistoff. Doseringsregimer kan justeres for å gi optimal ønsket respons (f.eks., en terapeutisk respons). For eksempel kan en enkelt bolus administreres, flere oppdelte doser kan administreres over tid eller dosen kan proporsjonelt reduseres eller økes som indikert av kravene for den terapeutiske situasjonen. Det er spesielt fordelaktig å formulere parenterale preparater i doseenhetsform for enkel administrering og ensartet dose. Doseenhetsform som anvendt heri angir fysisk atskilte enheter ordnet som enhetsdoser for subjektene som skal behandles; hver enhet inneholder en på forhånd bestemt mengde av den aktive forbindelsen beregnet å frembringe den ønskede terapeutiske effekten i assosiasjon med den nødvendige farmasøytiske bæreren.
50 Antistoffer kan administreres i en enkelt dose eller i multiple doser. De multiple dosene kan administreres via samme eller forskjellige administreringsveier og til samme eller forskjellige lokaliseringer. Alternativt kan antistoffer administreres som en formulering med vedvarende frigjøring, i hvilket tilfelle mindre hyppig administrering er nødvendig. Dose og frekvens kan variere avhengig av halveringstiden av antistoffet i pasienten og den behandlingsvarigheten som er ønsket. Dosen og frekvensen av administrering kan også variere avhengig av om behandlingen er profylaktisk eller terapeutisk. Ved profylaktiske anvendelser kan en relativt lav dose administreres ved relativt lange mellomrom over en lang tidsperiode. Ved terapeutiske anvendelser kan en relativt høy dose administreres, for eksempel helt til pasienten viser delvis eller fullstendig bedring av symptomer på sykdom. Følgelig kan et antistoff ifølge oppfinnelsen administreres: omtrent daglig, omtrent annenhver dag, omtrent hver tredje dag, omtrent hver fjerde dag, omtrent hver femte dag, omtrent hver sjette dag; omtrent hver uke, slik som hver., 6., 7., 8., 9. eller. dag; omtrent annenhver uke, slik som hver 11., 12., 13., 14., 1., 16. eller 17. dag; omtrent hver tredje uke, slik som hver 18., 19.,., 21., 22., 23. eller 24. dag; omtrent hver fjerde uke, slik som hver 2., 26., 27., 28., 29.,. eller 31. dag. Et antistoff ifølge oppfinnelsen kan også administreres "på forespørsel". Som nevnt ovenfor kan antistoffer ifølge oppfinnelsen administreres sammen med ett eller flere andre terapeutiske midler. Det andre midlet kan være et middel som vil fremme effektene av modulatoren. Det andre midlet kan være et middel som virker ved å fremme blodkoagulasjon, slik som en blodkoagulasjonsfaktor. Spesielt kan modulatorene ifølge oppfinnelsen administreres sammen med Faktor VII(a) eller FVIII(a). Det andre midlet kan være ment for å behandle andre symptomer eller lidelser hos pasienten. For eksempel kan det andre midlet være et analgetisk, anestetisk, immunosuppressivt eller antiinflammatorisk middel. Kombinert administrering av to eller flere midler kan oppnås på flere forskjellige måter. I én utførelsesform kan antistoffet og det andre midlet administreres sammen i et enkelt preparat. I en annen utførelsesform kan antistoffet og det andre midlet administreres i separate preparater som del av en kombinert behandling. For eksempel kan modulatoren administreres før, etter eller samtidig med det andre midlet. Betegnelsen "behandling", som anvendt heri, refererer til medisinsk behandling av hvilket som helst menneske eller annet dyre-subjekt med behov derfor. Nevnte subjekt er forventet å ha gjennomgått fysisk undersøkelse ved en praktiserende lege eller en praktiserende veterinær, som har gitt en foreløpig eller endelig diagnose som ville indikere at anvendelse av nevnte spesifikke behandling er
51 fordelaktig for helsen til nevnte humane eller andre animalske subjekt. Reguleringen og formålet med nevnte behandling kan variere fra ett individ til et annet, i henhold til status quo av subjektets helse. Følgelig kan nevnte behandling være profylaktisk, palliativ, symptomatisk og/eller kurativ. Når det gjelder foreliggende oppfinnelse kan profylaktiske, palliative, symptomatiske og/eller kurative behandlinger representere separate aspekter av oppfinnelsen. Følgelig kan et antistoff ifølge oppfinnelsen administreres parenteralt. Et antistoff ifølge oppfinnelsen kan administreres intravenøst. Et antistoff ifølge oppfinnelsen kan administreres intramuskulært. Et antistoff ifølge oppfinnelsen kan administreres subkutant. Et antistoff ifølge oppfinnelsen kan administreres profylaktisk, Et antistoff ifølge oppfinnelsen kan administreres terapeutisk ("på forespørsel"). Et antistoff ifølge oppfinnelsen kan være i stand til å redusere blodtad i betydelig grad. Et antistoff ifølge oppfinnelsen kan være i stand til, i betydelig grad, å redusere blødningstid. Følgelig er oppfinnelsen også en fremgangsmåte for behandling av et subjekt med behov derfor med et monoklonalt antistoff som er i stand til å binde K2- domenet av TFPI, hvori mengden av monoklonalt antistoff administrert er slik at den metter dets mål. Mengden av monoklonalt antistoff administrert kan være slik at den metter løselig TFPI. Mengden av monoklonalt antistoff administrert kan være slik at den metter endotel-bundet TFPI. Betegnelsen "koagulopati", som anvendt heri, angir en økt blødningstendens som kan være forårsaket av hvilken som helst kvalitativ eller kvantitativ mangel på hvilken som helst pro-koagulativ komponent av den normale koagulasjonskaskaden, eller hvilken som helst oppregulering av fibrinolyse. Slike koagulopatier kan være medfødte og/eller ervervet og/eller iatrogene og blir identifisert av fagfolk på området. Ikke-begrensende eksempler på medfødte hypokoagulopatier er hemofili A, hemofili B, Faktor VII-mangel, Faktor XI -mangel, von Willebrands sykdom og trombocytopeni slik som Glanzmanns trombasteni og Bernard-Soulier syndrom. Et ikke-begrensende eksempel på en ervervet koagulopati er serinproteasemangel forårsaket av vitamin K-mangel; slik vitamin K-mangel kan være forårsaket av administrering av en vitamin K-antagonist, slik som warfarin. Ervervet koagulopati kan også oppstå etter omfattende traume. I dette tilfelle ellers kjent som "bloody vicious cycle", er det karakterisert ved hemodilusjon (dilutional trombocytopeni og fortynning av koagulasjonsfaktorer), hypotermi, forbruk av
52 koagulasjonsfaktorer og metabolske fortyrrelser (acidose). Væske("fluid")-terapi og økt fibrinolyse kan forverre denne situasjonen. Nevnte hemoragi kan være fra hvilken som helst del av kroppen. Hemofili A med "inhibitorer" (det vil si allo-antistoffer mot faktor VIII) og hemofili B med "inhibitorer" (det vil si, allo-antistoffer mot faktor IX) er ikkebegrensende eksempler på koagulopatier som er delvis medfødte og delvis ervervet. Et ikke-begrensende eksempel på en iatrogen koagulopati er en overdosering av antikoagulant medikament - slik som heparin, aspirin, warfarin og andre hemmere av blodplateaggregasjon - som kan preskriberes for behandling av tromboembolisk sykdom. Et annet, ikke-begrensende eksempel på iatrogen koagulopati er den som blir indusert ved overdreven og/eller upassende væsketerapi, slik som den som kan bli indusert ved en blodtransfusjon. I én utførelsesform ifølge foreliggende oppfinnelse er hemoragi assosiert med hemofili A eller B. I en annen utførelsesform er hemoragi forbundet med hemofili A eller B med ervervede inhibitorer. I en annen utførelsesform er hemoragi forbundet med trombocytopeni. I en annen utførelsesform er hemoragi forbundet med von Willebrands sykdom. I en annen utførelsesform er hemoragi assosiert med alvorlig vevskade. I en annen utførelsesform er hemoragi assosiert med alvorlig traume. I en annen utførelsesform er hemoragi assosiert med kirurgi. I en annen utførelsesform er hemoragi forbundet med hemoragisk gastritt og/eller enteritt. I en annen utførelsesform er hemoragi overstrømmende uterin blødning, slik som ved morkakeløsning. I en annen utførelsesform oppstår hemoragi i organer med begrenset mulighet for mekanisk hemostase, slik som intrakranielt, intraauralt eller intraokulært. I en annen utførelsesform er hemoragi forbundet med antikoagulantbehandling. Et antistoff ifølge foreliggende oppfinnelse kan anvendes for å behandle et subjekt med koagulopati. Følgelig er oppfinnelsen også anvendelse av et monoklonalt antistoff, som er i stand til å binde til K2-domenet av TFPI, for behandling av et subjekt med behov derfor; så vel som anvendelse av nevnte antistoff for fremstilling av et medikament for behandling av et subjekt med behov derfor. Videre er oppfinnelsen en fremgangsmåte for behandling av et subjekt med behov derfor med et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI. Anvendelse av nevnte monoklonale antistoff ifølge oppfinnelsen kan, i betydelig grad, redusere blodtap. Anvendelse av nevnte monoklonale antistoff ifølge oppfinnelsen kan redusere blødningstid i betydelig grad.
53 1 Videre kan anvendelse av nevnte monoklonale antistoff ifølge oppfinnelsen redusere in vivo koagulasjonstid uten å forårsake transient trombocytopeni. UTFØRELSESFORMER Det følgende er en ikke-begrensende liste over utførelsesformer ifølge foreliggende beskrivelse: Utførelsesform 1 : Et monoklonalt antistoff som er i stand til å spesifikt binde til K2- domenet av TFPI, hvori nevnte antistoff er i stand til å binde til en epitop omfattende én eller flere rester valgt fra gruppen bestående av E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, F3, K36 og L0 fra SEQ ID NO: 2. 1 Utførelsesform 2: Det monoklonale antistoffet ifølge utførelsesform 1, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende en epitop omfattende rest E fra SEQ ID NO: 2. Utførelsesform 3: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene ovenfor, hvori nevnte antistoff er i stand til å spesifikt binde en epitop omfattende en epitop omfattende rest E11 fra SEQ ID NO: 2. Utførelsesform 4: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest D12 fra SEQ ID NO: 2. 2 Utførelsesform : Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å spesifikt binde en epitop omfattende rest P13 fra SEQ ID NO: 2. Utførelsesform 6: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest R17 fra SEQ ID NO: 2. 3 Utførelsesform 7: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest Y19 fra SEQ ID NO: 2.
54 2 Utførelsesform 8: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å spesifikt binde en epitop omfattende rest T21 fra SEQ ID NO: 2. Utførelsesform 9: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest Y23 fra SEQ ID NO: 2. Utførelsesform : Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å spesifikt binde en epitop omfattende rest F24 fra SEQ ID NO: 2. 1 Utførelsesform 11: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å spesifikt binde en epitop omfattende rest N26 fra SEQ ID NO: 2. Utførelsesform 12: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest Q28 fra SEQ ID NO: 2. Utførelsesform 13: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å spesifikt binde en epitop omfattende rest Q31 fra SEQ ID NO: 2. 2 Utførelsesform 14: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest C32 fra SEQ ID NO: 2. Utførelsesform 1: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest E33 fra SEQ ID NO: 2. 3 Utførelsesform 16: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å spesifikt binde en epitop omfattende rest R34 fra SEQ ID NO: 2.
55 3 Utførelsesform 17: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest F3 fra SEQ ID NO: 2. Utførelsesform 18: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest K36 fra SEQ ID NO: 2. Utførelsesform 19: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevte utførelsesformene, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende rest L0 fra SEQ ID NO: 2. 1 Utførelsesform : Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-16 og 18-19, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende restene E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, K36 og L0 fra SEQ ID NO: 2. Utførelsesform 21: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-3, -9, og 1-19, hvori nevnte antistoff er i stand til å binde spesifikt til en epitop omfattende restene E, E11, P13, R17, Y19, T21, Y23, Q28, Q31, E33, R34, F3, K36 og L0 fra SEQ ID NO: Utførelsesform 22: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI, hvori den lette kjeden av nevnte antistoff omfatter aminosyrerestene: E, i posisjonen svarende til posisjon 31, S, i posisjonen svarende til posisjon 32, D, i posisjonen svarende til posisjon 33, Y, i posisjonen svarende til posisjon 37, A, i posisjonen svarende til posisjon 96, T, i posisjonen svarende til posisjon 97 og F, i posisjonen svarende til posisjon 99 fra SEQ ID NO: 1; og hvori den tunge kjeden av nevnte antistoff omfatter aminosyrerestene: N, i posisjonen svarende til posisjon 31, R, i posisjonen svarende til posisjon 3, S, i posisjonen svarende til posisjon 4, Y, i posisjonen svarende til posisjon 7, Y, i posisjonen svarende til posisjon 9,
56 4 F, i posisjonen svarende til posisjon 60, P, i posisjonen svarende til posisjon 61, D, i posisjonen svarende til posisjon 62, Q, i posisjonen svarende til posisjon 6, Y, i posisjonen svarende til posisjon 2, D, i posisjonen svarende til posisjon 3 og D, i posisjonen svarende til posisjon 6 fra SEQ ID NO: Utførelsesform 23: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1-21, hvori den lette kjeden av nevnte antistoff omfatter aminosyrerestene: E, i posisjonen svarende til posisjon 31, S, i posisjonen svarende til posisjon 32, D, i posisjonen svarende til posisjon 33, Y, i posisjonen svarende til posisjon 37, A, i posisjonen svarende til posisjon 96, T, i posisjonen svarende til posisjon 97 og F, i posisjonen svarende til posisjon 99 fra SEQ ID NO: 1; og hvori den tunge kjeden av nevnte antistoff omfatter aminosyrerestene: N, i posisjonen svarende til posisjon 31, R, i posisjonen svarende til posisjon 3, S, i posisjonen svarende til posisjon 4, Y, i posisjonen svarende til posisjon 7, Y, i posisjonen svarende til posisjon 9, F, i posisjonen svarende til posisjon 60, P, i posisjonen svarende til posisjon 61, D, i posisjonen svarende til posisjon 62, Q, i posisjonen svarende til posisjon 6, Y, i posisjonen svarende til posisjon 2, D, i posisjonen svarende til posisjon 3 og D, i posisjonen svarende til posisjon 6 fra SEQ ID NO: Utførelsesform 24: Det monoklonale antistoffet ifølge utførelsesform 22 eller utførelsesform 23, hvori nevnte tunge kjede videre omfatter en S i posisjonen svarende til posisjon 2 fra SEQ ID NO: 18.
57 Utførelsesform 2: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 22-23, hvori nevnte lette kjede videre omfatter en H i posisjonen svarende til posisjon 98 fra SEQ ID NO: 1 og nevnte tunge kjede videre omfatter en S i posisjonen svarende til posisjon 6 fra SEQ ID NO: 18. Utførelsesform 26: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter en CDR1-sekvens med aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18, hvori én av disse aminosyrene kan være substituert med en annen aminosyre. 1 Utførelsesform 27: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21 som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter en CDR1-sekvens med aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18, hvori én av disse aminosyrene kan være substituert med en annen aminosyre. Utførelsesform 28: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter en CDR2-sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. 2 3 Utførelsesform 29: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21 som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter en CDR2-sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18, én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. Utførelsesform : Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. Utførelsesform 31: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21 som er i stand til å binde til Kunitz 2 (K2)-domenet av
58 6 vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. Utførelsesform 32: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den lette kjeden av nevnte antistoff omfatter en CDR1-sekvens med aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. 1 Utførelsesform 33: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21 som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den lette kjeden av nevnte antistoff omfatter en CDR1-sekvens med aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. Utførelsesform 34: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den lette kjeden av nevnte antistoff omfatter en CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre. 2 Utførelsesform 3: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21 som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den lette kjeden av nevnte antistoff omfatter en CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre. 3 Utførelsesform 36: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den lette kjeden av nevnte antistoff omfatter en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre.
59 7 Utførelsesform 37: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21 som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den lette kjeden av nevnte antistoff omfatter en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre. 1 2 Utførelsesform 38: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18, hvori én av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR2-sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. Utførelsesform 39: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21, hvori den tunge kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18, hvori én av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR2-sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre. 3 Utførelsesform 40: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den lette kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller
60 8 en CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre. 1 Utførelsesform 41: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21, hvori den lette kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være substituert med en ulik aminosyre; og/eller en CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en ulik aminosyre; og/eller en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en ulik aminosyre. 2 3 Utførelsesform 42: Et monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18, hvori én av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR2-sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og hvori den lette kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre; og/eller
61 9 en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre. 1 2 Utførelsesform 43: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1 til 21, hvori den tunge kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18, hvori én av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR2-sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og hvori den lette kjeden av nevnte antistoff omfatter: en CDR1-sekvens med aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre. 3 Utførelsesform 44: Et monoklonalt antistoff ifølge hvilke som helst av utførelsesformene 26-43, hvori nevnte aminosyresubstitusjoner ikke omfatter aminosyrene: N, i posisjonen svarende til posisjon 31 av CDR1-regionen av SEQ ID NO: 18; R, i posisjonen svarende til posisjon 3; S, i posisjonen svarende til posisjon 4; S, i posisjonen svarende til posisjon 6; Y, i posisjonen svarende til posisjon 7; Y, i posisjonen svarende til posisjon 9; F, i posisjonen svarende til posisjon 60; P, i posisjonen svarende til posisjon 61;
62 60 1 D, i posisjonen svarende til posisjon 62; og Q, i posisjonen svarende til posisjon 6; av CDR2-regionen av SEQ ID NO: 18. Y, i posisjonen svarende til posisjon 2; D, i posisjonen svarende til posisjon 3; og D, i posisjonen svarende til posisjon 6; av CDR3-regionen fra SEQ ID NO: 18. E, i posisjonen svarende til posisjon 31; S, i posisjonen svarende til posisjon 32; D, i posisjonen svarende til posisjon 33; og Y, i posisjonen svarende til posisjon 37; av CDR1-regionen fra SEQ ID NO: 1. A, i posisjonen svarende til posisjon 96; T, i posisjonen svarende til posisjon 97; H, i posisjonen svarende til posisjon 98; og F, i posisjonen svarende til posisjon 99; av CDR3-regionen fra SEQ ID NO: 1. Utførelsesform 4: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 26-44, hvori nevnte aminosyresubstitusjon er en konservativ substitusjon. 2 Utførelsesform 46: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 26-4, hvori den tunge kjeden av nevnte antistoff omfatter: en CDR1-sekvens som omfatter aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18; og en CDR2-sekvens som omfatter aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18; og en CDR3-sekvens som omfatter aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: Utførelsesform 47: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 26-46, hvori den lette kjeden av nevnte antistoff omfatter: en CDR1-sekvens som omfatter aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1; og en CDR2-sekvens som omfatter aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1; og
63 61 en CDR3-sekvens som omfatter aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1. 1 Utførelsesform 48: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 46-47, hvori den tunge kjeden omfatter: en CDR1-sekvens som omfatter aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18; og en CDR2-sekvens som omfatter aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18; og en CDR3-sekvens som omfatter aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18; og hvori den lette kjeden omfatter: en CDR1-sekvens som omfatter aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1; og en CDR2-sekvens som omfatter aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1; og en CDR3-sekvens som omfatter aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1. Utførelsesform 49: Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene, hvori den lette kjeden av nevnte antistoff omfatter SEQ ID NO: 1. 2 Utførelsesform 0: Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene, hvori den tunge kjeden av nevnte antistoff omfatter SEQ ID NO: 18. Utførelsesform 1: Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene, hvori nevnte antistoff omfatter SEQ ID NO: 1 og SEQ ID NO: Utførelsesform 2: Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene, hvori nevnte antistoff omfatter den lette kjeden av SEQ ID NO: 21. Utførelsesform 3: Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene, hvori nevnte antistoff omfatter den tunge kjeden fra SEQ ID NO: 24.
64 62 Utførelsesform 4: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 2-3, hvori nevnte antistoff omfatter SEQ ID NO: 21 og SEQ ID NO: 24. Utførelsesform : Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene, hvilket er et humanisert antistoff. 1 Utførelsesform 6: Det monoklonale antistoffet ifølge utførelsesform, i hvilken struktur ( framework )-region 2 av den tunge kjeden omfatter aminosyrene: T, i posisjonen svarende til posisjon 40, E, i posisjonen svarende til posisjon 42, R, i posisjonen svarende til posisjon 44 og A, i posisjonen svarende til posisjon 49 av SEQ ID NO: 18. Utførelsesform 7: Det monoklonale antistoffet ifølge utførelsesform, i hvilken struktur ( framework )-region 2 av den tunge kjeden omfatter aminosyrene svarende til posisjonene 36 til 49 (WVRQTPEKRLEWVA) fra SEQ ID NO: 18. Utførelsesform 8: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-4, hvilket er et humant antistoff. 2 Utførelsesform 9: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-4, hvilket er et kimært antistoff. Utførelsesform 60: Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene, hvori isotypen av nevnte antistoff er IgG. Utførelsesform 61: Det monoklonale antistoffet ifølge utførelsesform 60, hvori nevnte isotype er IgG1, IgG2 eller IgG4. 3 Utførelsesform 62: Det monoklonale antistoffet ifølge utførelsesform 61, hvori isotypen av nevnte antistoff er IgG4. Utførelsesform 63: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 60-62, hvori minst én aminosyre fra Fc-regionen av nevnte antistoff har blitt erstattet av en annen aminosyre.
65 63 Utførelsesform 64: Det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene hvori Fc-regionen av nevnte antistoff er minst 80 %, slik som minst 8 %, slik som minst 90 %, slik som minst 9 %, slik som 9-0 % identisk med aminosyrene fra SEQ ID NO: 24. Utførelsesform 6: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab0281. Utførelsesform 66: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-64, som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab Utførelsesform 67: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab4904. Utførelsesform 68: Et monoklonalt antistoff ifølge hvilke som helst av utførelsesformene 1-66, som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab4904. Utførelsesform 69: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab Utførelsesform 70: Et monoklonalt antistoff, ifølge hvilke som helst av utførelsesformene 1-68, som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab2974. Utførelsesform 71: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab Utførelsesform 72: Et monoklonalt antistoff ifølge hvilke som helst av utførelsesformene 1-70, som er i stand til å binde til K2-domenet av TFPI med en høyere affinitet enn mab Utførelsesform 73: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI, hvori K D for nevnte antistoff er mindre enn 0,8 nm, slik som mindre enn 0,7 nm, slik som mindre enn 0,6 nm, slik som mindre enn 0, nm, slik som mindre enn
66 64 0,4 nm, slik som mindre enn 0,3 nm, slik som mindre enn 0,2 nm, slik som mindre enn 0,1 nm, slik som mindre enn 0,0 nm, slik som mindre enn 0,02 nm. Utførelsesform 74: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-73, hvori K D for nevnte antistoff er mindre enn 0,8 nm, slik som mindre enn 0,7 nm, slik som mindre enn 0,6 nm, slik som mindre enn 0, nm, slik som mindre enn 0,4 nm, slik som mindre enn 0,3 nm, slik som mindre enn 0,2 nm, slik som mindre enn 0,1 nm, slik som mindre enn 0,0 nm, slik som mindre enn 0,02 nm. Utførelsesform 7: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevnte utførelsesformene som er i stand til å binde til K2-domenet av blodplateassosiert TFPI. 1 Utførelsesform 76: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevnte utførelsesformene som er i stand til å inhibere løselig TFPI. Utførelsesform 77: Det monoklonale antistoffet ifølge utførelsesform 76, hvori nevnte løselige TFPI kan bli fullstendig inhibert. Utførelsesform 78: Det monoklonale antistoffet ifølge hvilke som helst av de ovennevnte utførelsesformene som er i stand til å inhibere lipoprotein-bundet TFPI. 2 Utførelsesform 79: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene ovenfor som er i stand til å inhibere endotelcelle-bundet TFPI. Utførelsesform 80: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI slik at FXa bibeholder sin aktivitet med minst 91 %, slik som minst 92 %, slik som minst 93 %, slik som minst 94 %, slik som minst 9 %, slik som minst 96 %, slik som minst 97 %, slik som minst 98 %, slik som minst 99 %, slik som 99-0 %, som målt i en FXa inhiberings-assay. 3 Utførelsesform 81: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-79, som er i stand til å binde til TFPI slik at FXa bibeholder sin aktivitet med minst 91 %, slik som minst 92 %, slik som minst 93 %, slik som minst 94 %, slik som minst 9 %, slik som minst 96 %, slik som minst 97 %, slik som minst 98 %, slik som minst 99 %, slik som 99-0 % som målt i en FXa inhiberingsassay.
67 6 Utførelsesform 82: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI slik at prosentandelen av fri TFPI i et subjekt blir redusert til mindre enn %, slik som mindre enn 29 %, slik som mindre enn 28 %, slik som mindre enn 27 %, slik som mindre enn 26 %, slik som mindre enn 2 %, slik som mindre enn 24 %, slik som mindre enn 23 %, slik som mindre enn 22 %, slik som mindre enn 21 %, slik som mindre enn %, slik som mindre enn 19 %, slik som mindre enn 18 %, slik som mindre enn 17 %, slik som mindre enn 16 %, slik som mindre enn 1 %, slik som mindre enn 14 %, slik som mindre enn 13 %, slik som mindre enn 12 %, slik som mindre enn 11 %, slik som mindre enn %, slik som mindre enn 9 %, slik som mindre enn 8 %, slik som mindre enn 7 %, slik som mindre enn 6 %, slik som mindre enn %, slik som mindre enn 4 %, slik som mindre enn 3 %, slik som mindre enn 2 %, slik som mindre enn 1 %, slik som mellom 1 % og 0 %. 1 2 Utførelsesform 83: Et monoklonalt antistoff ifølge hvilke som helst av kravene 1-81, som er i stand til å binde til K2-domenet av TFPI slik at prosentandelen av fri TFPI i et subjekt blir redusert til mindre enn %, slik som mindre enn 29 %, slik som mindre enn 28 %, slik som mindre enn 27 %, slik som mindre enn 26 %, slik som mindre enn 2 %, slik som mindre enn 24 %, slik som mindre enn 23 %, slik som mindre enn 22 %, slik som mindre enn 21 %, slik som mindre enn %, slik som mindre enn 19 %, slik som mindre enn 18 %, slik som mindre enn 17 %, slik som mindre enn 16 %, slik som mindre enn 1 %, slik som mindre enn 14 %, slik som mindre enn 13 %, slik som mindre enn 12 %, slik som mindre enn 11 %, slik som mindre enn %, slik som mindre enn 9 %, slik som mindre enn 8 %, slik som mindre enn 7 %, slik som mindre enn 6 %, slik som mindre enn %, slik som mindre enn 4 %, slik som mindre enn 3 %, slik som mindre enn 2 %, slik som mindre enn 1 %, slik som mellom 1 % og 0 %. 3 Utførelsesform 84: Det monoklonale antistoffet ifølge utførelsesform 83, hvori mengden av fri TFPI i et subjekt blir redusert til nevnte prosentandel i løpet av de første 28 dagene, slik som i løpet av de første 27 dagene, slik som i løpet av de første 26 dagene, slik som i løpet av de første 2 dagene, slik som i løpet av de første 24 dagene, slik som i løpet av de første 23 dagene, slik som i løpet av de første 22 dagene, slik som i løpet av de første 21 dagene, slik som i løpet av de første dagene, slik som i løpet av de første 19 dagene, slik som i løpet av de første 18 dagene, slik som i løpet av de første 17 dagene, slik som i løpet av de første 16 dagene, slik som i løpet av de første 1 dagene, slik som i løpet av de første 14 dagene, slik som i løpet av de første 13 dagene, slik som i løpet av de
68 66 første 12 dagene, slik som i løpet av de første 11 dagene, slik som i løpet av de første dagene, slik som i løpet av de første 9 dagene, slik som i løpet av de første 8 dagene, slik som i løpet av de første 7 dagene, slik som i løpet av de første 6 dagene, slik som i løpet av de første dagene, slik som i løpet av de første 4 dagene, slik som i løpet av de første 3 dagene, slik som i løpet av de første 2 dagene, slik som i løpet av den første dagen etter administrering av nevnte monoklonale antistoff til nevnte individ. 1 Utførelsesform 8: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI og som er i stand til å nøytralisere TFPI-inhiberingen av membranbundet FVIIa/TF/FXa med minst %, slik som minst 60 %, slik som minst 6 %, slik som minst 70 %, slik som minst 7 %, slik som minst 80 %, slik som minst 8 %, slik som minst 90 %, slik som minst 9 %, slik som opp til 0 %, slik som 0 %, som målt i en FVIIa/TF/FXa inhibitor assay, når TFPI er mettet med nevnte antistoff. Utførelsesform 86: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-84, hvori nevnte antistoff er i stand til å nøytralisere TFPIinhiberingen av membranbundet FVIIa/TF/FXa med minst %, slik som minst 60 %, slik som minst 6 %, slik som minst 70 %, slik som minst 7 %, slik som minst 80 %, slik som minst 8 %, slik som minst 90 %, slik som minst 9 %, slik som opp til 0 %, slik som 0 %, som målt i en FVIIa/TF/FXa inhibitor assay, når TFPI er mettet med nevnte antistoff. 2 Utførelsesform 87: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI og som reduserer in vivo koagulasjonstid uten å redusere blodplatetelling i betydelig grad. Utførelsesform 88: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-86, hvori nevnte antistoff reduserer in vivo koagulasjonstid uten å redusere blodplatetelling i betydelig grad. 3 Utførelsesform 89: Det monoklonale antistoffet ifølge utførelsesformene 88, hvori nevnte blodplatetelling ikke synker til omtrent 80 %, slik som omtrent 7 %, slik som omtrent 70 %, slik som omtrent 6 %, slik som omtrent 60 %, slik som omtrent %, slik som omtrent 0 %, slik som omtrent 4 %, slik som omtrent 40 %, slik som omtrent 3 %, slik som omtrent %, slik som omtrent 2 % av den opprinnelige blodplatetellingen.
69 67 Utførelsesform 90: Et monoklonalt antistoff som er i stand til å binde til K2-domenet av TFPI og som reduserer in vivo koagulasjonstid uten å forårsake transient trombocytopeni. Utførelsesform 91: Det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-89, hvori nevnte antistoff reduserer in vivo koagulasjonstid uten å forårsake transient trombocytopeni. Utførelsesform 92: Et fragment av det monoklonale antistoffet ifølge hvilke som helst av de foregående utførelsesformene. 1 Utførelsesform 93: Fragmentet ifølge utførelsesform 92, hvilket er et Fab-fragment, et F(ab') 2 -fragment, et Fab'-fragment, et Fd-fragment, et Fv-fragment eller et dabfragment. Utførelsesform 94: En variant av det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene, hvilken er en delesjonsvariant eller en insersjonsvariant. Utførelsesform 9: En farmasøytisk formulering omfattende det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene Utførelsesform 96: En farmasøytisk formulering omfattende det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94, hvori nevnte formulering er egnet for parenteral anvendelse. Utførelsesform 97: En farmasøytisk formulering omfattende det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94, hvori nevnte antistoff er egnet for intravenøs anvendelse. Utførelsesform 98: En farmasøytisk formulering omfattende det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94, hvori nevnte antistoff er egnet for intramuskulær anvendelse. 3 Utførelsesform 99: En farmasøytisk formulering omfattende det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94, hvori nevnte antistoff er egnet for subkutan anvendelse.
70 68 Utførelsesform 0: Anvendelse av det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94 for fremstilling av et medikament egnet for parenteral administrering. Utførelsesform 1: Anvendelse av det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94 for fremstilling av et medikament egnet for intravenøs administrering. Utførelsesform 2: Anvendelse av det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94 for fremstilling av et medikament egnet for intramuskulær administrering. 1 Utførelsesform 3: Anvendelse av det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene 1-94 for fremstilling av et medikament egnet for subkutan administrering. Utførelsesform 4: Anvendelse av et monoklonalt antistoff ifølge hvilke som helst av utførelsesformene 1-94, for behandling av et subjekt med en koagulopati. Utførelsesform : Anvendelse ifølge utførelsesform 4, hvori nevnte subjekt har hvilken som helst medfødt, ervervet og/eller iatrogen koagulopati, som for eksempel kan være valgt fra gruppen bestående av hemofili A, med eller uten inhibitorer, og hemofili B, med eller uten inhibitorer. 2 Utførelsesform 6: Anvendelse ifølge hvilke som helst av utførelsesformene 9-, hvori nevnte monoklonale antistoff reduserer blodtap i betydelig grad. 3 Utførelsesform 7: Anvendelse ifølge hvilke som helst av utførelsesformene 9-6, hvori nevnte monoklonale antistoff reduserer blødningstid i betydelig grad. Utførelsesform 8: Anvendelse ifølge hvilke som helst av utførelsesformene 9-7, hvori mengden av monoklonalt antistoff administrert resulterer i en plasmakonsentrasjon på omtrent μg/ml til omtrent 40 μg/ml, slik som omtrent 1-3 μg/ml, slik som omtrent -1 μg/ml, slik som omtrent 1- μg/ml, slik som omtrent -2 μg/ml, slik som omtrent 2- μg/ml, slik som omtrent -3 μg/ml, slik som omtrent 3-40 μg/ml, av nevnte monoklonale antistoff.
71 69 Utførelsesform 9: Fremgangsmåte for behandling av et subjekt med en koagulopati, omfattende å administrere til nevnte subjekt det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene Utførelsesform 1: Fremgangsmåte ifølge utførelsesform 9, hvori nevnte koagulopati er hvilken som helst av medfødt, ervervet og/eller iatrogen koagulopati, som for eksempel kan være valgt fra gruppen bestående av hemofili A, med eller uten inhibitorer, og hemofili B, med eller uten inhibitorer. Utførelsesform 111: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-1, hvori nevnte monoklonale antistoff er i stand til å redusere blodtap i betydelig grad. 1 Utførelsesform 112: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-111, hvori nevnte monoklonale antistoff er i stand til å redusere blødningstid i betydelig grad. Utførelsesform 113: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-112, hvori mengden av monoklonalt antistoff administrert er slik at den metter dets mål. 2 Utførelsesform 114: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-113, hvori mengden av monoklonalt antistoff administrert er slik at den metter løselig TFPI. Utførelsesform 11: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-114, hvori nevnte administrerte antistoff er i stand til å fullstendig inhibere løselig TFPI. Utførelsesform 116: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-11, hvori nevnte monoklonale antistoff blir administrert i en mengde tilstrekkelig til å mette endotel-bundet TFPI. 3 Utførelsesform 117: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-116, hvori mengden av monoklonalt antistoff administrert fører til en plasmakonsentrasjon på omtrent μg/ml til omtrent 40 μg/ml, slik som omtrent 1-3 μg/ml, slik som omtrent -1 μg/ml, slik som omtrent 1- μg/ml, slik som
72 70 omtrent -2 μg/ml, slik som omtrent 2- μg/ml, slik som omtrent -3 μg/ml, slik som omtrent 3-40 μg/ml, av nevnte monoklonale antistoff. Utførelsesform 118: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-117, hvori en enkelt dose kan administreres. Utførelsesform 119: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-118, hvori multiple doser kan administreres. Utførelsesform 1: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-119, hvori nevnte antistoff kan administreres daglig. 1 Utførelsesform 121: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-1, hvori nevnte antistoff kan administreres annenhver dag. Utførelsesform 122: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-121, hvori nevnte antistoff kan administreres hver tredje dag. Utførelsesform 123: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-122, hvori nevnte antistoff kan administreres hver fjerde dag. Utførelsesform 124: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-123, hvori nevnte antistoff kan administreres hver femte dag. 2 Utførelsesform 12: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-124, hvori nevnte antistoff kan administreres hver sjette dag. Utførelsesform 126: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-12, hvori nevnte monoklonale antistoff kan administreres omtrent hver uke, slik som hver., 6., 7., 8., 9. eller. dag. 3 Utførelsesform 127: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-126, hvori nevnte monoklonale antistoff kan administreres omtrent annenhver uke, slik som hver 11., 12., 13., 14., 1., 16. eller 17. dag. Utførelsesform 128: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-127, hvori nevnte monoklonale antistoff kan administreres omtrent hver tredje uke, slik som hver 18., 19.,., 21., 22., 23. eller 24. dag.
73 71 1 Utførelsesform 129: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-128, hvori nevnte monoklonale antistoff kan administreres omtrent hver fjerde uke, slik som hver 2., 26., 27., 28., 29.,. eller 31. dag. Utførelsesform 1: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-129, hvori dosen kan være omtrent 0,1- mg/kg, slik som omtrent 0,1-1 mg/kg, slik som omtrent 1-2 mg/kg eller omtrent 2-3 mg/kg eller omtrent 4- mg/kg eller omtrent -6 mg/kg eller omtrent 6-7 mg/kg eller omtrent 7-8 mg/kg eller omtrent 8-9 mg/kg eller omtrent 9- mg/kg; eller omtrent -21 mg/kg, slik som omtrent -11 mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent 14-1 mg/kg, eller omtrent 1-16 mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent mg/kg, eller omtrent 19- mg/kg eller omtrent -21 mg/kg. Utførelsesform 131: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-1, hvori dosen kan være omtrent 2 til 0 mg/kg, slik som omtrent -0 mg/kg, slik som omtrent -170 mg/kg, slik som omtrent 0- mg/kg, slik som omtrent 0-0 mg/kg, slik som omtrent mg/kg, slik som omtrent -0 mg/kg, slik som omtrent - mg/kg. Utførelsesform 132: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-131, hvori nevnte monoklonale antistoff kan administreres parenteralt. 2 Utførelsesform 133: Fremgangsmåte ifølge utførelsesform 132, hvori nevnte monoklonale antistoff kan administreres intravenøst. Utførelsesform 134: Fremgangsmåte ifølge utførelsesform 133, hvori dosen av nevnte monoklonale antistoff kan være omtrent - mg/kg. Utførelsesform 13: Fremgangsmåte ifølge hvilke som helst av utførelsesformene , hvori det monoklonale antistoffet kan administreres annenhver uke. 3 Utførelsesform 136: Fremgangsmåte ifølge hvilke som helst av utførelsesformene , hvori det monoklonale antistoffet kan administreres hver tredje uke. Utførelsesform 137: Fremgangsmåte ifølge hvilke som helst av utførelsesformene , hvori det monoklonale antistoffet kan administreres hver fjerde uke.
74 72 Utførelsesform 138: Fremgangsmåte ifølge utførelsesform 133, hvori dosen av nevnte monoklonale antistoff kan være omtrent - mg/kg og nevnte monoklonale antistoff kan administreres annenhver uke. Utførelsesform 139: Fremgangsmåte ifølge utførelsesform 133, hvori dosen av nevnte monoklonale antistoff kan være omtrent - mg/kg og nevnte monoklonale antistoff kan administreres hver tredje uke. Utførelsesform 140: Fremgangsmåte ifølge utførelsesform 133, hvori dosen av nevnte monoklonale antistoff kan være omtrent - mg/kg og nevnte monoklonale antistoff kan administreres hver fjerde uke. 1 Utførelsesform 141: Fremgangsmåte ifølge utførelsesform 132, hvori nevnte monoklonale antistoff kan administreres intramuskulært. Utførelsesform 142: Fremgangsmåte ifølge utførelsesform 132, hvori nevnte monoklonale antistoff kan administreres subkutant. Utførelsesform 143: Fremgangsmåte ifølge utførelsesform 132, hvori dosen av nevnte monoklonale antistoff kan være omtrent 1 mg/kg. 2 Utførelsesform 144: Fremgangsmåte ifølge hvilke som helst av utførelsesformene , hvori det monoklonale antistoffet kan administreres daglig. Utførelsesform 14: Fremgangsmåte ifølge hvilke som helst av utførelsesformene , hvori det monoklonale antistoffet kan administreres annenhver dag. Utførelsesform 146: Fremgangsmåte ifølge hvilke som helst av utførelsesformene , hvori dosen av nevnte monoklonale antistoff kan være omtrent 1 mg/kg og hvori nevnte monoklonale antistoff kan administreres daglig. 3 Utførelsesform 147: Fremgangsmåte ifølge hvilke som helst av utførelsesformene , hvori dosen av nevnte monoklonale antistoff kan være omtrent 1 mg/kg og hvori nevnte monoklonale antistoff kan administreres annenhver dag. Utførelsesform 148: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-147, hvori nevnte antistoff kan administreres profylaktisk.
75 73 Utførelsesform 149: Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-148, hvori nevnte antistoff kan administreres terapeutisk ("på forespørsel"). Utførelsesform : Fremgangsmåte ifølge hvilke som helst av utførelsesformene 9-149, hvori nevnte administrerte antistoff er i stand til å fullstendig (0 %) inhibere løselig TFPI. Utførelsesform 11: Et polynukleotid som koder for det monoklonale antistoffet ifølge hvilke som helst av utførelsesformene Utførelsesform 12: Et polynukleotid ifølge utførelsesform 11, hvilket omfatter minst én sekvens valgt fra gruppen bestående av SEQ ID NO: 13, 16, 19 og Utførelsesform 13: Et polynukleotid ifølge utførelsesform 12, hvilket omfatter SEQ ID NO: 19. Utførelsesform 14: Et polynukleotid ifølge utførelsesform 12, hvilket omfatter SEQ ID NO. 22. Utførelsesform 1: Et polynukleotid ifølge utførelsesform 12, hvilket omfatter SEQ ID NO: 19 og Utførelsesform 16: En eukaryot celle som omfatter polynukleotidet ifølge hvilke som helst av utførelsesformene Utførelsesform 17: En eukaryot celle som uttrykker det monoklonale antistoffet, eller fragment derav, ifølge hvilke som helst av utførelsesformene Utførelsesform 18: Den eukaryote cellen ifølge utførelsesform 17, hvilken er en mammalsk celle. 3 Utførelsesform 19: Den eukaryote cellen ifølge utførelsesform 17, hvilken er en gjærcelle. Utførelsesform 160: Den mammalske cellen ifølge utførelsesform 18, hvilken er valgt fra gruppen bestående av HEK293, CHO, BHK, NSO og human retina-celler.
76 74 EKSEMPLER Foreliggende oppfinnelse er ytterligere illustrert ved de følgende eksemplene som ikke skal oppfattes som ytterligere begrensende. Eksempel 1: Produksjon og karakterisering av monoklonale antistoffer rettet mot TFPI Monoklonale antistoffer ble fremkalt mot vevsfaktorsignalveisinhibitor (TFPI). Et monoklonalt antistoff som har den ønskede bindingsspesifisiteten ble identifisert, klonet og sekvensert. Dette antistoffet ble funnet å betydelig redusere blødningstiden i huden in vivo og førte ikke til signifikant reduksjon i blodplatetallet. 1 Metoder og resultater Alle sett ble anvendt i henhold til fabrikantens instruksjoner. Forkortelser: HC: tung kjede; LC: lett kjede; VH : variabelt domene - tung kjede; VL: variabelt domene - lett kjede; PCR: polymerasekjedereaksjon. 2 Immunisering og fusjon Mus ble immunisert med både fullengde TFPI og den korte versjonen TFPIB161B som inneholder kun de første to Kunitz-domenene. RBF-mus ble anvendt for immunisering og produksjon av mus monoklonale antistoffer. Injeksjoner ble foretatt subkutant i ryggen hos musene. μg protein ble blandet med complete Freund's adjuvans for den første injeksjonen. I de følgende immuniseringene ble incomplete Freund's adjuvans anvendt med samme konsentrasjon av antigenet. Ti dager etter siste immunisering, ble blod fra øyet fra mus screenet ved ELISA for TFPI-spesifikke antistoffer. Mus med positiv serum-titer ble boostet med μg TFPI ved intravenøs injeksjon, og avlivet etter tre dager. Miltene ble fjernet aseptisk og dispergert til en enkeltcellesuspensjon. Fusjon av miltceller og myelomceller ble utført ved PEG-metoden eller ved elektrofusjon. Bindings assay: ELISA Immunoplater ble belagt med anti-mus IgG. Kultursupernatanter fra hybridomcellene ble tilsatt til platene og, etter vasking, ble løselig biotinylert human TFPI eller TFPIB161B tilsatt for å teste for spesifikk binding. 3 Nøytralisering-assays: FXa assay og TF/FVIIa/FXa assay FXa inhiberings-assay: en fast/fiksert konsentrasjon av TFPI som fører til 90 % inhibering av FXa ble preinkubert med kultursupernatanter fra hybridomceller inneholdende anti TFPI monoklonale antistoffer og tilsatt til FXa pluss FXa-spesifikt
77 7 kromogent substrat. Denne analysen angår TFPI binding til FXa (beskrevet i nærmere detaljer i Eksempel 6). FVIIa/TF/FXa inhiberings-assay: 1) Inkubering av kultursupernatanter fra hybridomceller inneholdende anti TFPI monoklonale antistoffer anti og fiksert TFPI (90 % inhibering av FVIIa/TF); 2) Inkubering av TFPI + FVIIa + TF + FXa; 3) tilsetning av FX (FX>>FXa) fulgt av inkubering med FXa kromogent substrat (beskrevet i nærmere detaljer i Eksempel 7). 1 Fortynnet ("dilute") protrombintid (dpt) En fortynnet protrombin (PT)-analyse: humant plasma i kombinasjon med fortynnet humant tromboplastin (TF kilde). Koagulasjonstid i plasma ble målt ved tilsetning av økende konsentrasjoner av protein A renset TFPI monoklonalt antistoff for å se etter doseavhengig reduksjon av koagulasjonstid. FVIIa (2 nm) var positiv kontroll og må forkorte denne koagulasjonstiden. Bindingsinteraksjonsanalyse Bindingsinteraksjonsanalyse ble oppnådd ved overflate plasmonresonans i et Biacore 00. Binding av det relevante monoklonale antistoffet ved en fast konsentrasjon ble oppnådd med immobilisert mus anti-igg. Forskjellige konsentrasjoner av TFPI ble testet. Bestemmelse av bindingskonstanter (k on, k off, K D ) ble oppnådd ved å forutsette en 1:1-interaksjon av TFPI og antistoffet av interesse (beskrevet i nærmere detaljer i Eksempel 8). 2 Trombelastografi Dette registrerer kinetikken av koagulasjonen og fibrinolyse i helblod. Hemofili A-lignende lidelse induseres ved preinkubasjon av blodet med nøytraliserende anti-fviii IgG. 3 Antistoff-kloning og sekvensering Murine tung kjede og lett kjede sekvenser for et anti-tfpi antistoff ble klonet fra et hybridom : TFPI-4F36A1B2 (forkortet heri til 4F36). Totalt RNA, ekstrahert fra hybridomceller ved anvendelse av RNeasy-Mini Kit fra Qiagen, ble anvendt som templater for cdna-syntese. cdna ble syntetisert i en '-RACE reaksjon ved anvendelse av SMART TM RACE cdna amplifikasjons-kit fra Clontech. Deretter ble mål-amplifikasjon av HC- og LC-sekvenser utført ved PCR ved anvendelse av Phusion Hot Star polymerase (Finnzymes) og universell primer mix (UPM) inkludert i SMART TM RACE -settet som en forover-primer. En revers primer med følgende sekvens ble anvendt for HC (VH-domene)-amplifikasjon: '-
78 76 1 CCCTTGACCAGGCATCCCAG-S' (primer nr. 129). En revers primer med følgende sekvens ble anvendt for LC-amplifikasjon : '- GCTCTAGACTAACACTCATTCCTGTTGAAGCTCTTG-3' (primer nr. 69). PCR-produkter ble separert ved gelelektroforese, ekstrahert ved anvendelse av GFX PCR DNA og Gel Band Purification Kit fra GE Healthcare Bio-Sciences og klonet for sekvensering ved anvendelse av et Zero Blunt TOPO PCR Cloning Kit og kjemisk kompetent TOPlO E.coli fra Invitrogen. Koloni PCR ble utført for selekterte kolonier ved anvendelse av en AmpliTaq Gold Mas-ter Mix fra Applied Biosystems og M13uni/M13rev primers. Koloni PCR clean-up ble utført ved anvendelse av ExoSAP- IT enzyme mix (usb). Sekvensering ble utført ved MWG Biotech, Martinsried Tyskland ved anvendelse av enten M13uni(-21)/M13rev(-29) eller T3/T7 sekvenseringsprimere. Sekvenser ble analysert og annotert ved anvendelse av VectorNTI programmet Fra hybridom TFPI-4F36A1B2 ble en enkelt unik murin kappa type LC identifisert og en enkelt unik murin HC, subklasse IgG1. LC-sekvens er gitt i SEQ ID NO: 6 og HC-sekvens er gitt i SEQ ID NO:. VH og VL-sekvenser er vist i Figur 2, lederpeptid-sekvenser er ikke inkludert. 2 3 Epitoper TFPI1 omfatter tre Kunitz-domener (se Figur 4). Overflate-tilgjengelige rester av Kunitz-domenene av TFPI1 ble identifisert fra eksisterende strukturer av TFPI1-2. Spesielt er rester med en relativ tilgjengelighet høyere enn 40 % ansett å være overflate-tilgjengelige. For TFPI1-2 omfatter dette (se Figur ) : aminosyrene 94-9, 98, 0-1, , , 131, 134, og Eksempel 2: Kloning og sekvensering av mus TFPI4F36A1B2 mab Dette eksemplet beskriver kloning og sekvensering av de murine tung kjede og lett kjede sekvensene av anti-tfpi antistoff: TFPI4F36A1B2. Totalt RNA ble ekstrahert fra hybridomceller ved anvendelse av RNeasy-Mini Kit fra Qiagen og anvendt som templat for cdna-syntese. cdna ble syntetisert i en '-RACE reaksjon ved anvendelse av SMART TM RACE cdna amplifikasjonskit fra Clontech. Etterfølgende mål-amplifikasjon av HC og LC-sekvenser ble utført ved PCR ved anvendelse av Phusion Hot Start polymerase (Finnzymes) og universell primer mix (UPM) inkludert i SMART TM RACE kit som forover primer. Revers-primeren identifisert som SEQ ID NO: 11 ble anvendt for HC (VH domene)-amplifikasjon og revers-primeren identifisert som SEQ ID NO: 12 ble anvendt for LC amplifikasjon. PCR-produkter ble separert ved gelelektroforese, ekstrahert ved anvendelse av GFX PCR DNA & Gel Band Purification Kit fra GE Healthcare Bio-Sciences og klonet for
79 77 sekvensering ved anvendelse av et Zero Blunt TOPO PCR Cloning Kit og kjemisk kompetent TOPlO E.coli (Invitrogen). Koloni-PCR ble utført på utvalgte kolonier ved anvendelse av en AmpliTaq Gold Master Mix fra Applied Biosystems og M13uni/M13rev primere. Koloni-PCR clean-up ble utført ved anvendelse av ExoSAP- IT enzyme mix (USB). Sekvensering ble utført ved MWG Biotech, Martinsried Tyskland ved anvendelse av enten M13uni(-21)/M13rev(-29) eller T3/T7 sekvenseringsprimere. Sekvenser ble analysert og annotert ved anvendelse av VectorNTI programmet. Alle sett og reagenser ble anvendt i henhold til fabrikantens instruksjoner. En enkelt unik murin kappa type LC og en enkelt unik murin HC, subklasse IgG1 ble identifisert. Nukleinsyre- og aminosyresekvensene for den variable lette kjeden er vist i SEQ ID NO: 3 og, henholdsvis. Nukleinsyre- og aminosyresekvenser for den variable tunge kjeden er vist i SEQ ID NO: 7 og 9, henholdsvis. Lederpeptid-sekvenser er ikke inkludert i disse sekvensene. 1 2 BLAST-søk De translaterte anti-tfpi4f36a1b2 VL og VH aminosyresekvensene ble anvendt som spørre ("query")-sekvenser. BLAST-søk ble foretatt mot sekvenser i Uniprot databasen ved anvendelse av BLASTp translasjonsprogram. Output for anti- TFPI4F36A1B2 VH frembringer sammenstillinger av hvilke > av de 0 høyeste identitet-scores var murine Ig tung kjede sekvenser. De høyeste identitet-scores var 81 % (99/121) mot en mus Ig tung kjede. Output for anti-tfpi4f36a1b2 VL frembringer sammenstillinger av hvilke > av de 0 høyeste identitet-scores var murine Ig kappa lett kjede sekvenser. Det høyeste identitet-score var 92 % (/113) mot en mus Ig kappa lett kjede. Til slutt representerer VH og VLsekvensene for anti- TFPI4F36A1B2 nye unike sekvenser. 3 Fremstilling av mus anti-tfpi4f36a1b2 ekspresjonsvektorer En rekke CMV promotor-baserte ekspresjonsvektorer (ptt vektorer) ble fremstilt for transient ekspresjon av mus TFPI4F36-antistoffet i det HEK293-6E EBNA-baserte ekspresjonssystemet utviklet av Yves Durocher (Durocher et al. Nucleic Acid Research, 02). I tillegg til CMV-promotoren, inneholder vektorene et pmb1 origo, et EBV origo og Amp resistensgenet. Regionen som svarer til fullengde anti-tfpi4f36a1b2 LC (inkludert den opprinnelige signalpeptidsekvensen) ble PCR-amplifisert fra de opprinnelige TOPO sekvenseringsklonene ved anvendelse av primere spesifikke for de N og C-terminale sekvensene. Sense-primeren inneholdt et terminalt HindIII restriksjonssetesekvenser for kloningsformål og en Kozak-sekvens ('- GCCGCCACC- 3') umiddelbart oppstrøms for ATG startkodonet. Anti-sense primeren inneholdt et stopp-kodon fulgt
80 78 1 av en Xbal restriksjonssetesekvens, umiddelbart nedstrøms for den kodende sekvensen. Det fremstilte PCR-fragmentet ble underlagt restriksjonskløyving, klonet inn i det multiple kloningssetet (MCS) av en linearisert ptt-basert vektor og transformert inn i E. coli for seleksjon. Sekvensen av den endelige konstruksjonen ble bekreftet ved DNA-sekvensering. Regionen svarende til VH-domenet (inkludert den opprinnelige signalpeptidsekvensen) ble PCR-amplifisert fra de opprinnelige TOPO sekvenseringsklonene ved anvendelse av primere spesifikke for den N-terminale sekvensen og VH/CH transisjonssekvensen. Sense-primeren inneholdt et terminalt NotI restriksjonssetesekvenser for kloningsformål og en Kozak-sekvens ('- GCCGCCACC- 3') umiddelbart oppstrøms for ATG startkodonet. Anti-sense primeren inneholdt et Nhel restriksjonssete i leseramme nedstrøms for VH/CH transisjonen. Det fremstilte PCRfragmentet av VH-domenet ble underlagt restriksjonskløyving, klonet inn i en linearisert vektor inneholdende CH domene-sekvensen for en murin IgG1 og transformert inn i E. coli for seleksjon. Sekvensen av den endelige konstruksjonen ble bekreftet ved DNA-sekvensering. Det klonede og rekombinant uttrykte anti-tfpi4f36a1b2 antistoffet hadde samme profil og affinitet i alle analyser anvendt, som det opprinnelige hybridomavledete antistoffet. Fremgangsmåter anvendt for transient ekspresjon i HEK293-6Eceller er beskrevet i Eksempel Eksempel 3: Design og konstruksjon av en humanisert TFPI4F36 mab Mus anti-tfpi4f36a1b2 CDR-sekvensen ble annotert i henhold til Kabatdefinisjon og funnet å være som følger: CDR-H1 : NYAMS (aminosyrene 31-3 fra SEQ ID NO: 8). CDR-H2: TISRSGSYSYFPDSVQG (aminosyrene 0-66 fra SEQ ID NO: 8). CDR-H3 : LGGYDEGDAMDS (aminosyrene 99-1 fra SEQ ID NO: 8). CDR-L1 : KSSQSLLESDGKTYLN (aminosyrene fra SEQ ID NO: 4). CDR-L2: LVSILDS (aminosyrene -61 fra SEQ ID NO: 4). CDR-L3 : LQATHFPQT (aminosyrene 94-2 fra SEQ ID NO: 4). En 3D-modell av anti-tfpi4f36a1b2 ble bygget i Modeller (www, salilab.org/modeller/) basert på de strukturelle templatene 2GJJ (mab mot Her2erbb2) og 1X9Q (hab mot flourescein). Et BLASTp-søk i en human kjønnscelle V database med anti-tfpi4f36a1b2 VL og VH meldte tilbake de følgende fire potensielle kjønnscellesekvensene: Tung kjede : VH3_21 eller VH (E-verdier < 1e-4) Lett kjede : VKII_A18 eller VKII_A1 (E-verdier < 3e-4)
81 Etter manuell undesøkelse av treff og sammenstillinger, ble kjønnscellesekvensene VH3_21 og VKII_A18 valgt ut som HC og LC humanisering strukturer ("frameworks"), henholdsvis. De korresponderende kjønnscelle J- segmentene ble valgt ut basert på sekvenssammenstilling som JH6 og JK4. Sammenstillingen mellom anti-tfpi4f36a1b2 og de selekterte kjønnscellesekvensene er vist i kombinasjon med den første CDR graftede versjonen av den humaniserte TFPI4F36. Sekvensidentiteten mellom anti- TFPI4F36A1B2 og de humane skaffold (HC: VH3_21/JH6 og LC: VKII_A18/JK4) er svært høy som illustrert med stjerner under sekvensen. Hver stjerne markerer en posisjon med sekvensidentitet. Den initielle humaniserte VH-konstruksjonen ble utformet i henhold til en minimal CDR grafting strategi, i hvilken CDR-H2 er graftet i en kortere versjon (rest 0-8) enn Kabat-definisjonen (rest 0-66). De resterende CDR'ene ble graftet i henhold til Kabat-definisjonen. CDR'ene (Kabat definisjon) er listet opp som graftet nedenfor; restene vist i fet skrift for CDR-H2 er humane kjønnscellerester. CDR-H1 : NYAMS (aminosyrene 31-3 fra SEQ ID NO: 18). CDR-H2: TISRSGSYSYYADSVKG (aminosyrene 0-66 fra SEQ ID NO: 28). CDR-H3 : LGGYDEGDAMDS (aminosyrene 99-1 fra SEQ ID NO: 18). CDR-L1 : KSSQSLLESDGKTYLN (aminosyrene fra SEQ ID NO: 1). CDR-L2: LVSILDS (aminosyrene -61 fra SEQ ID NO: 1). CDR-L3: LQATHFPQT (aminosyrene 94-2 fra SEQ ID NO: 1). Sammensetningen av CDR-H2 i den endelige humaniserte varianten HzTFPI4F36 er listet opp nedenfor og matchet CDR-H2 listet opp for mus antistoffet anti- TFPI4F36A1B2. CDR-H2: TISRSGSYSYFPDSVQG (aminosyrene 0-66 fra SEQ ID NO: 18). Figur 1 viser sekvensene av VH (A) og VL (B)-domenene av mus anti- TFPI4F36A1B2 (SEQ ID NO: 8 og 4, henholdsvis) sammenstilt med humane kjønnscellesekvenser (SEQ ID NO: 32 og 31, henholdsvis) og de CDR graftede humaniserte TFPI4F36 sekvensene (SEQ ID NO: 28 og 26, henholdsvis). Kabat nummereringsskjemaet er anvendt, som vist over sekvensene i figuren, og CDR'ene i henhold til Kabat-definisjonen er vist i fet skrift. Forskjeller i struktur ("framework")-regionene mellom mus anti-tfpi4f36a1b2 og kjønnscellesekvensene er fremhevet i grått i anti-tfpi4f36a1b2-sekvensen. Stjerner angir posisjonene med sekvensidentitet mellom mus TFPI4F36 og humane kjønnscellesekvenser. Potensielle tilbakemutasjoner er fremhevet i grått i HzTFPI4F36-CDR-graftet sekvens (listet opp som hz4f36cdrgraft). Potensielle tilbakemutasjoner for de HzTFPI4F36-CDRgraftede konstruksjonene ble identifisert basert på de posisjonelle forskjellene funnet i
82 80 struktur ("framework")-regionene av mus TFPI4F36 og kjønnscellesekvensen. En 3D Figur 1 viser sekvens modellene for TFPI4F36 Fab-fragment ble også anvendt for å identifisere og prioritere potensielle tilbakemutasjoner. Listene over genererte tilbakemutasjoner i de humaniserte TFPI4F36 LC og HC er vist i tabellene 2 og 3, henholdsvis. 1 2 Fremstilling av ekspresjonsvektorer for humanisert TFPI4F36 DNA-sekvenser for humaniserte TFPI4F36 VH og VL-regioner ble syntetisert (GENEART AG) i henhold til humaniserings-designet for antistoffet beskrevet ovenfor. Sekvensene ble oppnådd med den grunnleggende minimal CDR grafting og ingen ytterligere tilbakemutasjoner. De respektive LC og HC kjønnscelle lederpeptidsekvensene ble tatt med i konstruksjonene så vel som en Kozak-sekvens ('- GCCGCCACC-3') umiddelbart oppstrøms for ATG startkodonet. ptt-baserte ekspresjonsvektorer ble fremstilt for transient ekspresjon av det humaniserte TFPI4F36 antistoffet som en human kappa/igg4(s241p) isotype. Prolinmutasjonen ved posisjon 241 (nummerering i henhold til Kabat, svarende til rest 228 ved EU nummereringssystemet (Edelman G. M. et AL., Proc. Natl. Acad. USA 63, 78-8 (1969)) ble innført i IgG4 hengselregionen for å eliminere dannelse av monomeriske antistoffragmenter, dvs. "halv-antistoffer" omfattende én LC og én HC. VH-fragmentet ble skåret ut fra GENEART kloningsvektor og klonet inn i en linearisert ptt-basert vektor inneholdende sekvensen for et humant IgG4(S241P) CH-domene senere transformert inn i E. coli for seleksjon. Sekvensen av den endelige konstruksjonen ble bekreftet ved DNA-sekvensering. VL-fragmentet ble skåret ut fra GENEART kloningsvektoren og klonet inn i en linearisert ptt-basert vektor inneholdende sekvensen for et humant kappa CL-domene og deretter transformert inn i E. coli for seleksjon. Sekvensen av den endelige konstruksjonen ble bekreftet ved DNA-sekvensering. Nukleinsyre- og aminosyresekvenser for VL, VH, LC og HC av CDR-graftet HzTFPI4F36 monoklonalt antistoff (signalpeptidsekvens utelatt) er gitt i sekvenslisten (SEQ ID NO: 26-). 3 Fremstilling av ekspresjonsvektorer for mus/ human kimær TFPI4F36 For å muliggjøre best mulig evaluering av de humaniserte TFPI4F36- variantene, ble en mus/ human kimær versjon av anti-tfpi4f36 antistoffet (ChimTFPI4F36) konstruert for å eliminere enhver forskjell relatert til konstant region origo og isotype. ptt-baserte ekspresjonsvektorer ble fremstilt for transient ekspresjon av kimært anti-tfpi4f36 antistoff med murine variable domener på human kappa/igg4(s241p) isotype skaffold.
83 81 1 Regionen svarende til VH-domenet ble PCR-amplifisert fra et anti- TFPI4F36A1B2 HC ekspresjonsplasmid ved anvendelse av en generisk ptt spesifikk primer og en primer spesifikk for den C-terminale enden av VH-domenet. Senseprimeren er spesifikk for en sekvenslengde oppstrøms for HindIII restriksjonssetet og ATG startkodonet. Anti-sense primeren inneholdt et Nhel restriksjonssete i leseramme i VH/CH transisjon sekvensen. Det fremstilte PCR-fragmentet ble underlagt restriksjonskløyving, klonet inn i en linearisert ptt-basert vektor inneholdende sekvensen for et humant IgG4(S241P) CH-domene og deretter transformert inn i E. coli for seleksjon. Sekvensen av den endelige konstruksjonen ble bekreftet ved DNA-sekvensering. Regionen svarende til VL-domenet ble PCR-amplifisert fra en TFPI4F36A1B2 LC ekspresjonsplasmid ved anvendelse av en generisk ptt-spesifikk primer og en primer spesifikk for den C-terminale enden av VL-domenet. Sense-primeren er spesifikk for en sekvens strekning oppstrøms for HindIII restriksjonssetet og startkodonet ATG. Anti-sense primeren inneholdt et BsiWI restriksjonssete i leseramme i VL/CL transisjon sekvensen. Det fremstilte PCR-fragmentet ble underlagt restriksjonskløyving, klonet inn i en linearisert ptt-basert vektor inneholdende sekvensen for et humant kappa CL domene og deretter transformert inn i E. coli for seleksjon. Sekvensen av den endelige konstruksjonen ble bekreftet ved DNA-sekvensering. 2 Rekombinant ekspresjon av mab-varianter De murine anti-tfpi4f36a1b2, kimære anti-tfpi4f36 og humaniserte TFPI4F36 antistoffvariantene ble uttrykt transient i HEK293-6E-celler ifølge en generisk antistoff ekspresjonsmetode. Den følgende fremgangsmåten beskriver den generiske transfeksjonsmetoden anvendt for suspensjon-tilpassede HEK293-6Eceller. 3 Opprettholdelse av celler HEK293-6E celler ble dyrket i suspensjon i Freestyle TM 293 ekspresjonsmedium (Gibco) supplert med 2 μg/ml Geneticin (Gibco), 0,1 % volum/volum av surfaktanten Pluronic F-68 (Gibco) og 1 % volum/volum Penicillin- Streptomycin (Gibco). Celler ble dyrket i Erlenmeyer rystekolber i rysteinkubatorer ved 37 C, 8 % CO 2 og 12 rpm og opprettholdt ved celletettheter på mellom 0,1-1, x 6 celler/ml. DNA transfeksjon
 μl media/ μg DNA, blandet og inkubert ved romtemperatur (23-2 C) i min.")
84 82 1 Celletettheten av kulturer anvendt for transfeksjon var 0,9-2,0 x 6 celler/ml. En blanding av 0, μg LC vektor-dna + 0, μg HC vektor DNA ble anvendt per ml cellekultur. DNA ble fortynnet i Opti-MEM media (Gibco) μl media/ μg DNA, blandet og inkubert ved romtemperatur (23-2 C) i min. 293Fectin TM (Invitrogen) ble anvendt som transfeksjonsreagens ved en konsentrasjon på 1 μl per μg DNA. 293Fectin TM ble fortynnet X i Opti-MEM media (Gibco), blandet og inkubert ved romtemperatur (23-2 C) i min. DNA og 293Fectin-løsningene ble blandet og hensatt for inkubasjon ved romtemperatur (23-2 C) i 2 min. DNA-293Fectin-blandingen ble deretter tilsatt direkte til cellekulturen. Den transfekterte cellekulturen ble overført til en rysteinkubator ved 37 C, 8 % CO 2 og 12 rpm. 3-6 dager etter transfeksjon ble cellekultursupernatanter høstet ved sentrifugering, fulgt av filtrering gjennom et 0,22 μm PES filter (Corning). Kvantitativ analyse av antistoffproduksjon ble utført ved Biolayer Interferometry direkte på rensede cellekultursupernatanter ved anvendelse av ForteBio Octet-systemet og protein A biosensorer eller kvantitativ protein A HPLC. 2 Aktivitetsanalyses av den CDR-graftede varianten av humanisert anti- TFPI4F36 Humanisering ved minimal CDR grafting resulterte i et dramatisk tap av affinitet forårsaket av effekt på både on- og off-rate. TFPI bindingsaffiniteten av den initielt graftede versjonen av det humaniserte TFPI4F36 antistoffet (HzTFPI4F36- CDRgraftet, i tabell 1 listet opp som Humanisert TFPI4F36) var minst 0 ganger lavere enn affiniteten på ~ pm av det opprinnelige mus TFPI4F36 antistoffet (se tabell 1). Retensjon av affinitet i det kimære antistoffet bekreftet at human kappa/igg4(s241p) FC ikke hadde noen effekt på antistoffaffinitet. Affinitetsanalysene ble utført ved anvendelse av SRP som beskrevet nedenfor. Tabell 1 3
85 Overflate plasmonresonans (Biacore) analyse av hztfpi4f36-tfpi interaksjon De kinetiske parametrene for interaksjonen mellom rekombinant human TFPI og opprinnelig murin anti-tfpi4f36a1b2, kimært anti-tfpi4f36, og ulike varianter av det humaniserte TFPI4F36 antistoffet ble bestemt ved SPR-analyse i Biacore, ved anvendelse av to forskjellige metoder. Initiell kinetikk rangerings-studier var basert på en metode for oppfanging ("capture") av rensede mabs som beskrevet i Eksempel 1. Disse ble fulgt av en direkte bindingskinetikk-metode på utvalgte mabkonstruksjoner, med det monoklonale antistoffet kovalent bundet via frie amingrupper til den karboksymetylerte dekstranmembranen (CM) på sensor chipoverflaten. Rekombinant human TFPI ble injisert i forskjellige konsentrasjoner, fulgt av en dissosiasjonsperiode med konstant bufferstrøm over sensor chip-overflaten som beskrevet i Eksempel 8. Seterettet mutagenese for å innføre tilbakemutasjoner i humanisert mab Basert på den lave affiniteten av den CDR-graftede versjonen av humanisert anti- TFPI4F36, ble en serie på 27 human-til mus reverse mutasjoner (referert til som tilbakemutasjoner) generert i den lette kjeden (LC) og tunge kjeden (HC) av HzTFPI4F36-CDRgraftet. Disse mutantene ble uttrykt, renset og analysert ved Biacore, enten som separate mutanter eller som LC/HC kombinasjonsmutanter. Listene over frembrakte mutasjoner er vist i tabellene 2 og 3, henholdsvis. Seterettet mutagenese ble utført for å innføre human-til-mus reverse mutasjoner (fra nå av referert til som tilbakemutasjoner) ved de spesifikk restene i de HzTFPI4F36-CDRgraftede LC/HC konstruksjonene som fremhevet i humaniserings-designet. Mutasjoner ble innført ved to forskjellige metoder: 1) QuickChange Seterettet eller Multi Seterettet mutagenese-sett fra Stratagene ble anvendt for å innføre punktmutasjoner og kombinasjonsmutasjoner. Settene ble anvendt i henhold til fabrikantens protokoll. 2) Standard 2-trinns overlappende PCR-metoder ble også anvendt for å innføre punktmutasjoner og for å frembringe kombinasjonsmutasjoner. LC og HC-ekspresjonsplasmidene for HzTFPI4F36-CDRgraftet ble anvendt som templater for de første rundene av mutagenese. I etterfølgende runder ble mutasjoner også innført ved anvendelse av tidligere muterte plasmider som templat. Sekvensene av alle endelige konstruksjoner ble bekreftet ved DNA-sekvensering. Tabell 2: Muterte varianter av HzTFPI4F36-CDRgraftet lett kjede

86 84 Tabell 3: Muterte varianter av HzTFPI4F36-CDRgraftet tung kjede
87 8 NO/EP
88 Mutasjonene i både LC og HC som listet opp i tabellene 2 og 3 er konsekvent nummerert i henhold til Kabat nummereringsskjemaet som vist i figur 1. LC-mutantene listet opp i Tabell 2 ble uttrykt som LC-mutanter kun sammen med villtype HC HzTFPI4F36 CDRgraftet. HC-mutantene listet opp i tabell 3 ble uttrykt som HC-mutanter kun sammen med villtype LC HzTFPI4F36 CDRgraftet. LC- HC kombinasjonsmutantene ble også uttrykt ved å kombinere forskjellige LC- og HCmutanter. Mutanter er konsekvent navngitt etter den muterte kjeden, dvs. den endelige humaniserte mab-varianten er uttrykt med villtype HzTFPI4F36-CDRgraftet LC og den muterte HzTFPI4F36 FR2, S49A, CDR2 HC. Transient HEK293-6Eekspresjon ble utført som beskrevet ovenfor. Det initielle settet av 9 punktmutasjoner (HzTFPI4F36 LC-S63T & HzTFPI4F36 HC- Q3E; G44R; S49A; Y9F; A60P; K64Q; S77T; A93T) var basert på et primært sett av tilbakemutasjoner fremhevet i humaniserings designet. Ingen av punktmutasjonenen reddet affiniteten av antistoffet, imidlertid ble mutasjoner i den andre human tung kjede struktur ("framework")-regionen (FR2, mellom CDR H1 og CDR H2) og i den C-terminale regionen av CDR H2 (utelatt i minimal CDR graftingsskjema) fremhevet som viktig for TFPI-binding. Affinitetsmålinger ved Biacore analyser ble utført som beskrevet ovenfor. De følgende rundene med mutagenese inkuderte flere patch mutanter i hvilke alle restene i individuelle regioner ble mutert samlet. Mutanten HzTFPI4F36 HC- Kabat CDR2, har 3 mutasjoner Y9F, A60P, K64Q i den C-terminale regionen av CDR H2, hvilket svarer til grafting CDR H2 i henhold til Kabat-definisjonen og ikke i henhold til minimal CDR grafting-skjemaet anvendt for den initielle HzTFPI4F36- CDRgraftede varianten. Denne patchede mutanten forbedret sammen med patch mutanter i LC FR2 og HC FR2 affiniteten av det humaniserte TFPI4F36 antistoffet betydelig, men ingen av de tre patch mutantene gjenopprettet individuelt den høye TFPI4F36-affiniteten. HC-mutanten med kombinerte mutasjoner i HC FR2 og CDR2 (A40T, G42E, G44R, S49A, Y9F, A60P, K64Q) gjenopprettet affinitet fullstendig. Denne mutanten innførte 7 ytterligere murine rester i antistoffsekvensen. Kombinasjon av LC FR2- mutanter (både FR2 mutasjoner og Y36L) og HC FR2 og/eller CDR2-mutanter førte også til mutanter med høy affinitet. Imidlertid førte kombinasjonen av disse LC/HC-mutantene konsekvent til lavere ekspresjonsutbytter sammenlignet med HCmutantene alene. Disse resultatene indikerte at inkludering av LC-mutantene hadde en negativ innvirkning på stabiliteten av disse antistoffvariantene, hvilket følgelig antyder at det finnes et ømtålig interaksjons-mønster mellom det humaniserte TFPI4F36 LC og HC.
89 I den siste serien av mutanter, ble de 7 mutasjonene i HC FR2 og CDR2 (A40T, G42E, G44R, S49A, Y9F, A60P, K64Q) analysert for å eliminere potensielt ikke-medvirkende tilbakemutasjoner. En serie på mutanter ble generert for å ta tak i dette formålet. I 3 mutanter ble tilbakemutasjoner utelukket i FR2: HzTFPI4F36 HC-G42E, G44R, CDR2 HzTFPI4F36 HC-FR2, CDR2 HzTFPI4F36 HC-G42E, G44R, S49A CDR2 I 2 mutanter ble ytterligere mutasjoner i CDR2 også eliminert : HzTFPI4F36 HC-G44R, A60P, K64Q HzTFPI4F36 HC-G42E, G44R, A60P, K64Q Ingen av mutantene var imidlertid on par med den kombinerte HC FR2 CDR2- mutanten. Enten ble affinitet eller ekspresjonsnivåer (eller begge) påvirket ved hvilken som helst reduksjon i HC FR2 CDR2 mutant-undergruppen. De to mutantene HzTFPI4F36 HC G42E, G44R, Y9F, A60P, K64Q med gjenværende tilbakemutasjoner og HzTFPI4F36 HC G42E, G44R, A60P, K64Q med 4 gjenværende tilbakemutasjoner ble valgt for grundig sammenligning med HzTFPI4F36 HC-FR2, S49A, CDR2. o HzTFPI4F36 HC-G42E, G44R, A60P, K64Q og HzTFPI4F36 HC-FR2, S49A, CDR2 ble uttrykt ved sammenlignbare nivåer, mens HzTFPI4F36 HC-G42E, G44R, CDR2 hadde et litt lavere ekspresjonsnivå. Akselererte biofysiske stabilitetsstudier viste ikke noen forskjeller i stabilitet i de tre variantene. o Affinitetene målt ved Biacore er listet opp nedenfor. o In vivo effekt målt i dpt assay (som beskrevet nedenfor) var sammenlignbare for alle de tre variantene. Tabell 4: Kinetiske parametere for interaksjonen mellom TFPI og humaniserte TFPI4F36-varianter.
90 88 1 Basert på dataene beskrevet ovenfor, testet den opprinnelige HC FR2, og CDR2-mutanten med 7 HC tilbakemutasjoner (A40T, G42E, G44R, S49A, Y9F, A60P, K64Q) bedre enn andre varianter; denne varianten er heri referert til som HzTFPI4F36 eller som mabtfpi21. Det er sannsynlig at CDR2-mutasjonene Y9F, A60P, K64Q påvirker antistoffaffinitet ved å interagere direkte med antigen. Mutasjonene A40T, G42E, G44R befinner seg i en FR2-dreining som forbinder CDRH1 og CDR H2, langt unna den antigenbindende overflaten og kunne bli balansert for stabilisering av LC-HC interaksjoner. Mutasjonen S49A er begravet midt i et svært hydrofobt cluster av sidekjeder hvilket kunne forklare hvorfor alanin er foretrukket fremfor serin ved denne posisjonen. Interessant nok blir derfor den høye affiniteten av HzTFPI4F36 oppnådd som en kombinasjon av mutasjoner som forbedrer den direkte antigeninteraksjonen og mutasjoner langt unna den antigenbindende regionen som stabiliserer antistoffet. Til slutt har HzTFPI4F36 en affinitet (K D ) på ~2 pm og inneholder 3 aminosyrerester avledet fra mus antistoffsekvensen, som tilsvarer,2 % av det totale antall rester i antistoffet. Aminosyresekvensene for variabel lett (VL) region, variabel tung (VH) region, lett kjede og tung kjede av en valgt humanisert konstruksjon, HzTFPI4F36 (mabtfpi 21), er vist i SEQ ID NO: 1, 18, 21 og 24, henholdsvis. 2 3 In-vitro effekt-assays Anti-TFPI4F36 antistoffet er i stand til å nøytralisere TFPI-mediert inhibering av koagulasjonsfaktor Xa (FXa) og komplekset av vevsfaktor (TF) og faktor VIIa (FVIIa). Aktivitetene av murine og humaniserte TFPI4F36 antistoff-varianter ble målt i en dilute protrombintid (dpt) test. dpt assay ble anvendt for å måle prokoagulant aktiviteten av anti-tfpi antistoffer. Økende plasmakonsentrasjoner av anti-tfpi antistoff forkorter dpt koagulasjonstiden. Eksempel 4: Rensing, krystallisering og struktur av Fab-fragmentet av MuTFPI4F36 (Fab) og det andre Kunitz-domenet (K2) av human Vevsfaktorsignalveisinhibitor (TFPI) Et fragment av TFPI omfattende dens andre Kunitz-domene (K2) og et C- terminalt His 6 -merke (SEQ ID NO: 2) ble ko-krystallisert med MuTFPI4F36 Fabfragmentet (Fab). Strukturen av komplekset ble oppløst ved røntgenkrystallografi. Den K2-bindende epitopen ble funnet å være bestående av restene E, E11, P13, R17, Y19, T21, Y23, Q28, Q31, E33, R34, F3, K36 og L0, Paratopen i Fab ble
91 89 funnet å omfatte restene E31, S32, D33, Y37, A96, T97, H98 og F99 av MuTFPI4F36 lett kjede (SEQ ID NO: 4) og restene N31, R3, S4, S6, Y7, Y9, F60, P61, D62, Q6, Y2, D3 og D6 av MuTFPI4F36 tung kjede (SEQ ID NO: 8). Materialer og metoder Analytisk størrelseseksklusjonskromatografi. Analytisk størrelseseksklusjonskromatografi (SEC) ble utført ved anvendelse av en Biosep S-00 (0 x 7,80 mm) kolonne (Phenomenex) eluert med PBS-buffer ( mm fosfat, mm NaCl, 3mM KCl, ph 7,) ved en strømningshastighet på 0,8 ml/min. 1 Fremstilling og rensing av Fab/K2-komplekset. Fab/K2-komplekset ble fremstilt ved å blande Fab (0,27mg/ml i PBS-buffer, ph 7,4) og K2 (0,29 mg/ml i PBS, ph 7,4) i et molart forhold på 1:1, (,4 mg Fab og 1,4 mg K2). Komplekset ble konsentrert i en sentrifugalfilter anordning (Amicon, kd mw cut-off) til en konsentrasjon på ~6,7 mg/ml. For å fjerne overskytende K2 ble den konsentrerte prøven applisert på en Superdex 7 (CV0) gelfiltreringskolonne eluert med PBS-buffer, ph 7,4 ved en strømningshastighet på 1 ml/min. Fraksjoner inneholdende Fab/K2-komplekset ble samlet og konsentrert til en proteinkonsentrasjon på 9,2 mg/ml. Denne løsningen ble anvendt for krystallisering. 2 Krystallisering av Fab/K2-komplekset. Fab/K2-komplekset ble krystallisert som staver ved "hengende dråpe" ("hanging drop")-metoden ved anvendelse av en presipitant løsning inneholdende 0,2 M tribasisk kaliumsitrat (ph 8,0) og % vekt/volum PEG 3,. Bestemmelse av krystallstruktur. Strukturen av Fab/K2-komplekset ble oppklart ved "molecular replacement"- metoden ved anvendelse av PDB-strukturene 1F8T og ITFX som templater for Fab og K2-molekylene, henholdsvis. 3 Resultater Komplekset mellom Fab og K2 ble fremstilt ved å tilsette overskudd av K2 til en løsning av Fab. Figur 6 viser kompleksdannelsen overvåket ved analytisk størrelseseksklusjonskromatografi (SEC). Ved denne metoden separeres molekyler i henhold til deres molekylstørrelse hvor de større speciene elueres tidligere enn mindre. Toppene svarende til K2 og Fab ble godt separert på grunn av den store forskjellen i molekylvekt (mw ~8 kda og 48 kda, henholdsvis). Tilsetning av K2 til
92 Fab-løsningen førte til den forventede mindre forskyvningen i topp-posisjonen mot kortere retensjonstider. Komplekset ble enkelt separert og oppnådd i ren form ved å skille ut overskytende K2 ved anvendelse av preparativ SEC. Betingelser for krystallisering av Fab/K2-komplekset ble screenet ved anvendelse av flere kommersielle krystalliserings-screens. Hengende dråpe-metoden ga stavformede krystaller egnet for enkeltkrystall røntgenanalyse og strukturen ble oppklart ved molecular replacement-metoden ved anvendelse av strukturer deponert i PDB som templater. Figur 7 viser den totale strukturen av Fab/K2-komplekset. Fremvist er de lette og tunge kjedene, som utgjør Fab-molekylet, og som fremviser den forventede immunglobulin ß-sandwich-folden karakteristisk for antistoffmolekyler. Også vist er CDR-løkkene som kommer i kontakt med antigenet og definerer spesifisiteten og affiniteten av antistoffet. Antigenet, K2, fremviser det karakteristiske enkelt anti-parallelle ß-sheet (ß1 (I-N26) og ß2 (Q31-Y37)) og den N-terminale α-heliksen (α1, L0-I6) som definerer Kunitz-folden (Figur 8). Til stede er også den eventuelle 3 -heliksen nær den N-terminale enden (α0, D-F8) fulgt av løkke 1 (L1, L9-Y19) som fører til ß1, som er forbundet med ß2 via en kort løkke (Lß, N27-K). I det C-terminale segmentet forbinder løkke 2 (L2, G38-T49) ß2 med αl. De karakteristiske tre disulfidbindingene (C7-C7, C16-C40 og C33-C3) forbinder α0 med α1, L1 med L2 og ß2 med α1, henholdsvis. To strukturer av K2 har blitt deponert i Worldwide Protein DataBank (PDB). En struktur, IADZ, er bestemt ved NMR-spektroskopi og representerer strukturen i fri løsning, mens den andre, ITFX, er bestemt ved røntgenkrystallografi og representerer K2 kompleksert med trypsin fra svin. Figur 9 viser den strukturelle superponeringen av K2 representert ved IADZ, ITFX og K2-molekylet i kompleks med Fab. Ryggrad linjene synes å være svært lik hos alle de tre strukturene, hvilket antyder at Kunitz-folden med dens tre stabiliserende disulfidbindinger er nokså rigid. 3 Beskrivelse av MuTFPI4F36 K2 bindingsepitopen Bindingsepitopen på antigenet K2, definert som rester i K2 inneholdende minst ett sidekjede tungt atom beliggende innenfor en avstand på 4 Å eller mindre fra et tungt atom i Fab, omfatter restene E, E11, P13, R17, Y19, T21, Y23, Q28, Q31, E33, R34, F3, K36 og L0 (Figur ). Kontaktrestene i K2 er lokalisert i Ll (E, E11, P13, R17, Y19), i ß-sheet-strukturen (T21, Y23, Q31, E33, R34, F3, K36) og den sammenbindende løkken, Lß (Q28) og, til slutt, en enkelt en i α1 (L0). Figur viser bindingsepitopen avbildet på både 3D-strukturene av K2 og den primære aminosyresekvensen.
93 Beskrivelse av MuTFPI4F36 paratopen Paratopen i MuTFPI4F36 Fab-fragmentet ble bestemt fra den samme røntgenstrukturen av komplekset mellom MuTFPI4F36 Fab og TFPI K2-domenet. Paratopen ble definert som de restene i MuTFPI4F36 Fab som har et tungt atom innenfor en avstand på mindre enn 4 Å fra et tungt atom i K2-domenet. Kontaktrestene i den lette kjeden er lokalisert ved restene E31, S32, D33, Y37, A96, T97, H98 og F99 fra SEQ ID NO: 4. Kontaktrestene i den tunge kjeden er lokalisert ved restene N31, R3, S4, S6, Y7, Y9, F60, P61, D62, Q6, Y2, D3 og D6 fra SEQ ID NO: 8. Lokaliseringen av paratopen er illustrert i Figur 3. Eksempel : Struktur av K2/HzTFPI4F36 Fab-komplekset Ved anvendelse av metodologi lik den beskrevet for bestemmelse av den tredimensjonale strukturen av MuTFPI4F36 Fab bundet til K2, ble strukturen av komplekset mellom Fab-fragmentet fra det humaniserte antistoffet, HzTFPI4F36, og K2 bestemt. Fab av HzTFPI4F36 ble som forventet funnet å binde til samme region på K2 som det murine Fab fra hvilket det er avledet. Den totale likheten mellom strukturene av de to kompleksene er tydelig i Figur 11, hvor ryggrad båndlinjer er overlagret for K2/TFPI4F36 Fab og K2/HzTFPI4F36 Fab-kompleksene. Epitopen (definert ved anvendelse av en 4 Å cut-off) på K2 ble, for HzTFPI4F36, funnet å omfatte restene E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, K36 og L0. Sammenlignet med strukturen av K2/MuTFPI4F36 Fab av murin opprinnelse, er D12, F24, N26 og C32 i det humaniserte K2/HzTFPI4F36- komplekset innenfor 4 Å cut-off, mens F3 er utenfor. Dette avspeiler mindre forskjeller i orientering av sidekjedene innenfor bindingsflatene av K2/MuTFPI4F36 Fab og K2/HzTFPI4F36 Fab-kompleksene, til tross for det faktum at CDR-regionene i MuTFPI4F36 og HzTFPI4F36 er identiske. Eksemplene 6 til 8 Funksjonen av HzTFPI4F36 (mabtfpi 21) ble sammenlignet med funksjonen av alle (fire) kommersielt tilgjengelige monoklonale antistoffer, av hvilke noen er tidligere nevnt å binde til K2-domenet av TFPI; av hvilke noen ikke har blitt beskrevet med hensyn til binding. 3 Eksempel 6: TFPI-nøytraliseringsassay: FXa-inhibering Materialer anvendt var BSA-buffer i assay (0 mm Hepes; 0,1 M NaCl, mm CaCl 2, 0,1 mg/ml BSA, ph 7,4) og reagensene vist i tabell. Tabell : Materialer anvendt
94 92 Metode : Rekombinant fullengde human TFPI (endelig konsentrasjon 6 nm)) ble blandet i BSA-buffer med økende konsentrasjoner av mab av interesse (sluttkonsentrasjon: - nm) i min. FXa ble tilsatt og inkubert min med blandingen i ytterligere min. Kromogent substrat S276 ble tilsatt og absorpsjonen ved 40 nm ble målt i 1 min i et Spectramax. 0 % aktivitet representerer aktiviteten av FXa uten tilsetning av TFPI. Tabell 6: Nøytralisering av TFPI-inhibering av FXa Konklusjon: Ved nm nøytraliserte HzTFPI4F36 (mabtfpi 21) TFPI-inhibering av FXa
95 93 fullstendig. Nesten ingen aktivitet ble detektert for mab0281, mab4904 og mab Eksempel 7: TFPI nøytralisingsassay: FVIIa /TF/ FXa inhibering Materialer anvendt var BSA-buffer (0 mm Hepes; 0,1 M NaCl, mm CaCl 2, 0,1 mg/ml BSA, ph 7,4) EDTA: 0 mm og reagensene listet opp i tabell 7. Tabell 7 1 Metode : Tilsett alle komponentene i sluttkonsentrasjonene angitt i tabellen. Tilsett 2 μl FX, 2 μl TFPI mab i varierende konsentrasjoner, 2 μl human TFPI, 2 μl FVIIa- TF (innovin) i mikrotiterbrønner. Inkubering i 40 min ved romtemperatur. Tilsett 0 μl EDTA fulgt av 0 μl S-276. Bland og les av platen i 1 min ved 40 nm i Spectramax. 0 % aktivitet er aktiviteten av FVIIa/TF/FX oppnådd uten TFPI til stede. Tabell 8: Nøytralisering av TFPI-inhibering av FVIIa/TF/FX
96 94 Konklusjon: Ved en mab-konsentrasjon på nm blir TFPI fullstendig nøytralisert av mabtfpi21. mab2974 når også metning men nøytraliserer ikke TFPI fullstendig (3 % nøytralisering). Eksempel 8: Bindingsinteraksjonsanalyse Tabell 9 Materialer anvendt var som listet opp i tabell 9. 1 Metode : Bindingsinteraksjonsanalyse ble oppnådd ved overflate plasmonresonans i et Biacore T-0 instrument. Oppfanging av det relevante monoklonale antistoffet ved en fast konsentrasjon ble oppnådd ved direkte immobilisering til en CM-chip av mab til et nivå på RU i mm natriumacetat ph 4,-,0. Fire-ganger


97 9 fortynninger av rekombinant human fullengde TFPI eller human TFPI kort form (1-161 aminosyrerester) fra 0 nm til 0,2 nm ble testet for binding til den immobiliserte mab. Kjøring og fortynningsbuffer: mm HEPES, mm, 0,00 % p, ph 7,4. Regenerering ble oppnådd med mm Glysin, ph 1,7. Bestemmelse av kinetikk og bindingskonstanter (k on, k off, K D ) ble oppnådd ved å forutsette en 1:1- interaksjon av TFPI og antistoffet av interesse ved anvendelse av Biacore T0 evaluering programvare. Resultater er vist i tabell. Konkurranse mellom de forskjellige mab om binding til TFPI når bundet til mabtfpi21 ("mab21", HzTFPI4F36) ble oppnådd ved immobilisering av mabtfpi21 til 000 RU ved en CM-chip fulgt av binding av 0 nm TFPI fulgt av varierende konsentrasjoner av mabs (2974, 0281, 4904, 29741) som skal testes med hensyn til konkurranse. Resultater er vist i tabell 11. Regenerering av chipen ble oppnådd ved mm Glysin, ph 1,7. 1 Tabell : Overflate plasmonresonans (SPR)-analyse. Binding til fullengde human TFPI. Kinetikk og bindingskonstanter. Tabell 11: SPR-analyse. Bindingskonstant for binding til fullengde human TFPI og TFPI 161 (K1 og K2-domener). Konkurranse med mabtfpi 21. Konklusjon
98 96 mabtfpi21 binder til TFPI med en høyere affinitet enn hvilke som helst av de andre mabs testet (KD 1 pm). Kun mab2974 konkurrerer om binding til samme sete som mab TFPI4F Eksempel 9: Nøytralisering av TFPI på humane vaskulære endotelceller fra navlestreng (HUVEC). Endotelceller uttrykker TFPI konstitutivt i en form som er bundet til celleoverflaten gjennom en glykosylfosfatidylinositol (GPI)-forankring. GPI-forankret TFPI inhiberer spesifikt TF-mediert aktivitet når TF blir uttrykt på samme celle som TFPI. For å vise at HzTFPI4F36 (mabtfpi21) nøytraliserer inhiberingen ved cellebundet TFPI mye mer effektivt enn mab 2974 anvendte vi humane vaskulære endotelceller fra navlestreng HUVEC; og for å indusere TF-ekspresjon ble disse cellene stimulert med TNFα (Sigma RBI) og IL1ß (Roche) før testing av FVIIa/TFkatalysert aktivering av FX. HUVEC-celler ble dyrket til konfluens i 96-brønners plater i EBM-2 medium (Clonetics) og stimulert med ng/ml TNFα og ng/ml IL1ß i 2 timer før testing. Testing ble utført i 2 mm HEPES, 137 mm NaCl, 3, mm KCl, mm CaCl, 1 mg/ml BSA (0,1 %) ph 7,4, og FX-aktivering ble fulgt i nærvær av antistoff (0 - nm) og med tilsetning av 0 pm FVIIa og 0 nm FX. Dannelse av FXa ble målt med 0,6 mm av et kromogent substrat, S-276 (Chromogenix) og kalibrert mot en FXa standardkurve. Figur 12 viser resultatene når inhiberingen ved cellebundet TFPI ble opphevet ved 0- nm HzTFPI4F36 eller mab TF/FVIIa-mediert aktivering av FX ble stimulert by HzTFPI4F36 med en halv maksimal effekt konsentrasjon, (EC 0 ~ nm) mens nesten ingen stimulering av FXa-generering ble observert med 2974 mab ved nm. Følgelig viser dette eksemplet at HzTFPI4F36, i motsetning til mab 2974, effektivt nøytraliserer inhibering av TF/FVIIa-mediert FX aktivering ved cellebundet TFPI. Eksempel : Nøytralisering av TFPI-inhibering av TF/ FVIIa-aktivitet om MDA- MB 231 humane brystkarsinomceller. MDA-MB 231-celler uttrykker konstitutivt høye nivåer av TF og ubetydelige mengder av TFPI på overflaten. Celleoverflate TF/FVIIa-mediert aktivering av FX kan inhiberes ved eksogent tilsatt TFPI. For å vise at HzTFPI4F36 nøytraliserer denne type TFPI-inhibering mye mer effektivt enn mab 2974 anvendte vi MDA-MB 231 celler og testet evnen av forskjellige konsentrasjoner av antistoff til å oppheve TFPIinhiberingen av FVIIa/TF-katalysert aktivering av FX.
99 97 MDA-MB 231 celler ble dyrket til konfluens i 96-brønners plater i DMEM Gibco cat# supplert med % FCS og 1 % P/S. Testing ble utført i 2 mm HEPES, 137 mm NaCl, 3. mm KCl, mm CaCl, 1 mg/ml BSA (0,1 %) ph 7,4, og FXaktivering ble fulgt i nærvær av antistoff (0 - nm) og med tilsetning av 2, nm fullengde human rekombinant TFPI, 0 pm FVIIa og 0 nm FX. Dannelse av FXa ble målt med 0,6 mm av et kromogent substrat, S-276 (Chromogenix). Absorbansen ved 40 nm ble målt kontinuerlig og FXa-aktivitet ble bestemt ved å måle stigningen av progresskurven ved 1 min etter initiering av reaksjonen. Figur 13 viser resultatene når inhibering ved TFPI ble opphevet ved 0- nm HzTFPI4F36 eller mab TF/FVIIa-mediert aktivering av FX ble stimulert av HzTFPI4F36 med en halv maksimal effekt konsentrasjon, (EC 0 ~ 2 nm) mens stimulering av FXa-generering ble oppnådd ved en vesentlig høyere konsentrasjon av 2974 mab (EC 0 > nm). 1 2 Eksempel 11: Kartlegging av bindingsepitopene av de anti-tfpi monoklonale antistoffene, HzTFPI4F36 og mab2974, ved anvendelse av ELISA. Bindingsepitopen for HzTFPI4F36 på TFPI Kunitz-domene 2 (K2) har blitt kartlagt ved oppløsning av krystallstrukturen av TFPI-K2/ HzTFPI4F36-komplekset. Effekten av å mutere enkelt aminosyrerester i TFPI inne i (E, R17 og Y19) og utenfor (D) bindingsepitopen for HzTFPI4F36 på bindingsaffiniteten til HzTFPI4F36 og mab2974 (R&Dsystems) ble analysert ved ELISA. TFPI variantene ble uttrykt i HEK293-F celler og ELISA ble utført ved anvendelse av det kondisjonerte mediet fra cellekulturene. Konsentrasjonene av TFPI-WT og TFPI-mutanter ble anslått ved en ELISA, som binder TFPI K1 (MAb4903, American Diagnostica) og K3 (MAb4F1, egenprodusert) og er følgelig ikke påvirket av mutasjonene. Effekten av mutasjonene på binding til HzTFPI4F36 ble analysert ved anvendelse av MAb4903 og HzTFPI4F36 i ELISA. Effekten på MAb2974-binding ble bestemt ved anvendelse av en ELISA med MAb2974 og MAb4F1. Effektene av mutasjonene i TFPI- Kunitz 2 på binding til HzTFPI4F36 og MAb2974 henholdsvis, ble beregnet relativt til TFPI-WT (0 % binding) og illustrert i figur 14. Tallene har blitt korrigert for forskjeller i ekspresjonsnivåer. 3 Konklusjon: Alaninmutasjon av de tre aminosyrerestene innenfor bindingsepitopen for HzTFPI4F36 resulterte i redusert binding til HzTFPI4F36, mens alaninsubstitusjon av
100 resten lokalisert utenfor epitopen (TFPI-DA) ikke hadde noen effekt. Kun én av de fire alaninmutantene, TFPI-Y19A, hadde redusert binding til MAb2974. Til slutt har HzTFPI4F36 og MAb2974 atskilte men overlappende bindingsepitoper lokalisert på TFPI-Kunitz 2. Eksempel 12: In vivo-studier Kaniner ble gjort transient hemofile ved intravenøs administrering av 00 RBU/kg av monoklonale anti-fviii-antistoffer. Etter minutter mottok kaninene 100 U/kg av anti-tfpi-antistoff (4F36; 1,93 mg/kg). Blødning i huden ble indusert 4 minutter etter administrering av anti-fviii-antistoff. 4F36-antistoffet forårsaket en signifikant reduksjon i blødningstiden i huden (Figur 1). Administrering av 4F36 antistoffet førte ikke til noen signifikant reduksjon i blodplatetallet (Figur 18). Et lignende forsøk ble gjentatt i hvilket tre grupper på åtte transient hemofile kaniner mottok enten isotype kontrollantistoff (negativ kontrollgruppe), 2 mg/kg anti-tfpi (mab 4F36) eller 9 mg/kg NovoSeven (positiv kontrollgruppe) minutter etter at blødning i huden ble indusert. Resultater er illustrert i figur 16: administrering av mab 4F36 resulterte i en betydelig reduksjon av blodtap (omtrent 8 %) hos alle recipienter, hvilket viser at mab4f36 kan anvendes "på forespørsel". Eksempel 13: Vurdering av dose effekt-forhold i kanin Dose-effekt forholdet for det humaniserte mab HzTFPI4F36 ble undersøkt i en kanin hemofili-modell. Kaniner ble gjort transient hemofile ved iv administrering av et monoklonalt anti-fviii-antistoff. Etter minutter mottok kaninene 0,, 1, 2 mg/kg HzTFPI4F36 eller et isotype kontrollantistoff. Etter ytterligere 3 minutter ble blødning i huden indusert, fulgt av en 60 minutters observasjonperiode. HzTFPI4F36 reduserte, betydelig og doseavhengig, blødningstid så vel som blodtap ved økning av dosen fra 0, til 2 mg/kg (figur 17). Følgelig ble en signifikant reduksjon av både blødningstid og blodtap oppnådd med 1 mg/kg HzTFPI4F36, svarende til en plasmakonsentrasjon på ng/ml HzTFPI4F36. Normalisering av blødningen ble oppnådd ved 2 mg/kg, svarende til en plasmakonsentrasjon på 980 ng/ml HzTFPI4F36. Disse data indikerer at den "effektive konsentrasjonen", f.eks. plasmakonsentrasjonen nødvendig for normalisering i foreliggende modell - av HzTFPI4F36 er i området og 980 ng/ml. Eksempel 14: PK/PD i kaniner - "Virkningsvarighet"
101 99 1 En farmakokinetisk studie av HzTFPI4F36 i kaniner dosert med mg/kg ble utført. Ved på forhånd bestemte tidspunkter i løpet av studien ble blodprøver tatt fra kaninene for farmakokinetisk profilering ved en ELISA som måler fri HzTFPI4F36 (vist i fig 3 nedenfor). Effektstudier ble utført ved 4 dager (96 timer), 7 dager (168 timer) og dager (240 timer) etter administrering, ved anvendelse av blødning i hud-modellen i transient hemofile kaniner, effekt tidspunktene er angitt i figur 19. Den farmakokinetiske profilen er bifasisk indikativ for mål-mediert clearance. Følgelig, over bøyningen av kurven er overskytende fri mab til stede (mab fri >TFPI total ), under bøyningen: mab fri < TFPI total. I god overensstemmelse med den farmakokinetiske profilen, var både blødningstid og blodtap betydelig redusert både ved 4 og 7 dager etter administrering av mg/kg HzTFPI4F36 intravenøst, mens ingen signifikant effekt ble observert etter dager (figur ). Disse data bekrefter at den effektive plasmakonsentrasjonen av HzTFPI4F36 i en blødning i hud-modell i hemofile kaniner er mellom og 980 ng/ml hvilket er nær TFPI metningsgrensen (bøyning av kurven). En enkelt iv. dose på mg/kg HzTFPI4F36 reduserte følgelig blødning i huden i minst 7 dager, hvilket svarer til den tidsperioden plasmakonsentrasjonen var over den "effektive konsentrasjonen". 2 Eksempel 1: Farmakokinetisk modell basert på ape PK-data En farmakokinetisk evaluering ble foretatt basert på en farmakokinetisk studie i aper, hvor både enkle og multiple doser ble administrert (figur 21). Dosenivåer var i området fra 2 til 0 mg/kg. PK-profilen i ape ( mg/kg, øvre panel) ligner den for kanin hvilket indikerer tilstedeværelse av lignende distribusjon av løselig og endotelbundet TFPI. Følgelig indikerer disse data at kanin effekt-data kan anvendes for å predikere effektområdet i ape. Videre er affiniteten av HzTFPI4F36 for human, ape og kanin TFPI lik (samme epitop) og lignede TFPI vevsdistribusjon i de tre artene muliggjør dosepredikeringer i ape og menneske. 3 Eksempel 16: Simuleringer Basert på modellen presentert ovenfor var det mulig å foreta en rekke simuleringer. Det viktigste formålet med simuleringene var å beskrive optimale doseringsregimer i en multippel dose-setting. Mål (TFPI)-konsentrasjonen var ikke kjent, men kanin effekt-dataene ovenfor muliggjør den antagelsen om at dersom målet er nær metning ved et nivå på 000 ng/ml så er full effekt i en blødningsmodell oppnådd. Derfor var det viktigste formålet med simuleringene å utrede hvilke dosenivåer over en tidsperiode som ville føre til fullstendig metning. Figur 22 fremviser en simulering av 1 mg/kg administrert subkutant. Figur 23 viser
 administrert IV annenhver uke.")
102 0 en simulering av 1 mg/kg HzTFPI4F36 (mabtfpi 21) administrert IV hver tredje uke. Figur 24 viser en simulering av mg/kg HzTFPI4F36 (mabtfpi 21) administrert IV annenhver uke. I korthet kan, basert på simuleringene ovenfor, følgende predikering av doseregime foretas for mennesker: Tabell 12: Doseregime
103 1 NO/EP
104 2 NO/EP
105 3 NO/EP
106 4 NO/EP
107 NO/EP
108 6 NO/EP
109 7 NO/EP
110 8 NO/EP
111 9 NO/EP
112 1 NO/EP
113 111
114 112 NO/EP
115 113 NO/EP
116 114 NO/EP
117 11 NO/EP
118 116 NO/EP
119 117 NO/EP
120 118 NO/EP
121 119 NO/EP
122 1 NO/EP
123 121 NO/EP
124 122 NO/EP
125 123 NO/EP
126 124 NO/EP
127 12 NO/EP
128 126 Patentkrav 1. Monoklonalt antistoff som er i stand til å binde spesifikt til K2-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori nevnte antistoff er i stand å binde spesifikt til en epitop omfattende rest R17 fra SEQ ID NO: 2 og K D for nevnte antistoff er lavere enn 0,8 nm, som bestemt ved anvendelse av overflate plasmonresonans. 2. Monoklonalt antistoff som er i stand til å binde spesifikt til K2-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori nevnte antistoff er i stand å binde til en epitop omfattende E, E11, D12, P13, R17, Y19, T21, Y23, F24, N26, Q28, Q31, C32, E33, R34, F3, K36 and L0 fra SEQ ID NO: Monoklonalt antistoff ifølge krav 1 eller 2, hvori K D for nevnte antistoff er lavere enn 0,7 nm, slik som lavere enn 0,6 nm, slik som lavere enn 0, nm, slik som lavere enn 0,4 nm, slik som lavere enn 0,3 nm, slik som lavere enn 0,2 nm, slik som lavere enn 0,1 nm, slik som lavere enn 0,0 nm, slik som lavere enn 0,02 nm som bestemt ved anvendelse av overflate plasmonresonans Monoklonalt antistoff ifølge hvilke som helst av kravene 1-3, hvori den lette kjeden av nevnte antistoff omfatter aminosyrerestene E, i posisjonen svarende til posisjon 31, S, i posisjonen svarende til posisjon 32, D, i posisjonen svarende til posisjon 33, Y, i posisjonen svarende til posisjon 37, A, i posisjonen svarende til posisjon 96, T, i posisjonen svarende til posisjon 97 og F, i posisjonen svarende til posisjon 99 av SEQ ID NO: 1; og hvori den tunge kjeden av nevnte antistoff omfatter aminosyrerestene N, i posisjonen svarende til posisjon 31, R, i posisjonen svarende til posisjon 3, S, i posisjonen svarende til posisjon 4, Y, i posisjonen svarende til posisjon 7, Y, i posisjonen svarende til posisjon 9, F, i posisjonen svarende til posisjon 60,
129 127 P, i posisjonen svarende til posisjon 61, D, i posisjonen svarende til posisjon 62, Q, i posisjonen svarende til posisjon 6, Y, i posisjonen svarende til posisjon 2, D, i posisjonen svarende til posisjon 3 og D, i posisjonen svarende til posisjon 6 av SEQ ID NO 18.. Monoklonalt antistoff ifølge krav 4, hvori nevnte tunge kjede ytterligere omfatter en S i posisjonen svarende til posisjon 2 av SEQ ID NO: 18 og/eller hvori nevnte lette kjede ytterligere omfatter en H i posisjonen svarende til posisjon 98 fra SEQ ID NO: 1 og nevnte tunge kjede ytterligere omfatter en S i posisjonen svarende til posisjon 6 fra SEQ ID NO: Monoklonalt antistoff ifølge hvilke som helst av kravene 1 til, hvori den tunge kjeden av nevnte antistoff omfatter en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være erstattet av en ulik aminosyre. 7. Monoklonalt antistoff ifølge hvilke som helst av kravene 1-, hvori den tunge kjeden av nevnte antistoff videre omfatter: 2 en CDR1-sekvens med aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18, hvori én av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR2-sekvens med aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og hvori den lette kjeden av nevnte antistoff omfatter: 3 en CDR1-sekvens med aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1, hvori én, to eller tre av disse aminosyrene kan være substituert med en annen aminosyre; og/eller en CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre; og/eller
130 128 en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1, hvori én eller to av disse aminosyrene kan være substituert med en annen aminosyre. 8. Monoklonalt antistoff ifølge hvilke som helst av kravene 6-7, hvori nevnte aminosyresubstitusjon er en konservativ substitusjon. 9. Monoklonalt antistoff som er i stand til å binde til Kunitz 2 (K2)-domenet av vevsfaktorsignalveisinhibitor (TFPI), hvori den tunge kjeden av nevnte antistoff omfatter: 1 en CDR1-sekvens av aminosyrene 31 til 3 (NYAMS) fra SEQ ID NO: 18; og en CDR2-sekvens av aminosyrene 0 til 66 (TISRSGSYSYFPDSVQG) fra SEQ ID NO: 18; og en CDR3-sekvens av aminosyrene 99 til 1 (LGGYDEGDAMDS) fra SEQ ID NO: 18. og hvori den lette kjeden av nevnte antistoff omfatter: 2 en CDR1-sekvens av aminosyrene 24 til 39 (KSSQSLLESDGKTYLN) fra SEQ ID NO: 1; og en CDR2-sekvens av aminosyrene til 61 (LVSILDS) fra SEQ ID NO: 1; og en CDR3-sekvens av aminosyrene 94 til 2 (LQATHFPQT) fra SEQ ID NO: 1.. Monoklonalt antistoff ifølge hvilke som helst av kravene 1-9, hvori den lette kjeden av nevnte antistoff omfatter SEQ ID NO: 1 og den tunge kjeden av nevnte antistoff omfatter SEQ ID NO: Monoklonalt antistoff ifølge hvilke som helst av kravene 1-, hvori den lette kjeden av nevnte antistoff omfatter SEQ ID NO: 21 og den tunge kjeden av nevnte antistoff omfatter SEQ ID NO: Monoklonalt antistoff ifølge hvilke som helst av kravene 1-11, som er i stand til å nøytralisere TFPI-inhiberingen av membranbundet FVIIa/TF/FXa med minst %, slik som minst 60 %, slik som minst 6 %, slik som minst 70 %, slik som minst 7 %, slik som minst 80 %, slik som minst 8 %, slik som
131 129 minst 90 %, slik som minst 9 %, slik som opp til 0 %, slik som 0 %, som målt i en FVIIa/TF/FXa inhibitor assay, når TFPI er mettet med nevnte antistoff. 13. Eukaryot celle, som uttrykker det monoklonale antistoffet, eller fragment derav, ifølge hvilke som helst av kravene Farmasøytisk preparat, hvori det omfatter et antistoff ifølge hvilke som helst av kravene 1-12 og en farmasøytisk akseptabel bærer eller fortynningsmiddel. 1. Monoklonalt antistoff ifølge hvilke som helst av kravene 1-12 for anvendelse ved behandling av et subjekt med en koagulopati Monoklonalt antistoff for anvendelse ifølge krav 1, hvori nevnte subjekt har hvilken som helst medfødt, ervervet og/eller iatrogen koagulopati, som for eksempel hemofili A, med eller uten inhibitorer, og hemofili B, med eller uten inhibitorer. 2
132 124 NO/EP

133 12 NO/EP
134 126 Figur 3
135 127 Figur 4
136 128 NO/EP
137 129 NO/EP
138 1 NO/EP

139 131 Figur 8
140 132 Figur 9
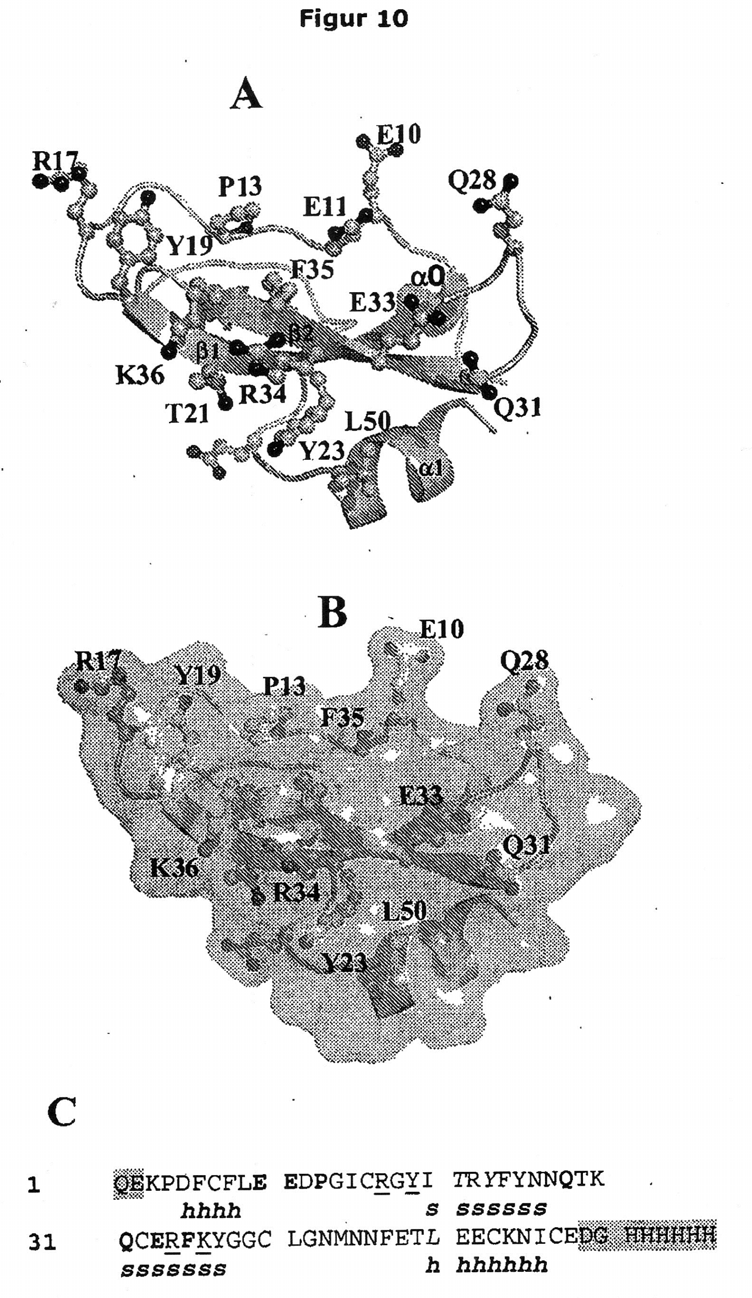
141 133 NO/EP

142 134 Figur 11
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 240726 B1 (19) NO NORGE (1) Int Cl. H0K 3/36 (2006.01) H0K 3/42 (2006.01) H0K 3/46 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.17 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 240726 B1 (19) NO NORGE (1) Int Cl. H0K 3/36 (2006.01) H0K 3/42 (2006.01) H0K 3/46 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.17 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2011486 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.09.17 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2011486 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.09.17 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2148670 B1 (19) NO NORGE (51) Int Cl. A61K 31/137 (2006.01) A61P 25/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.02 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2148670 B1 (19) NO NORGE (51) Int Cl. A61K 31/137 (2006.01) A61P 25/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.02 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2310382 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/4412 (2006.01) A61P 35/00 (2006.01) C07D 401/14 (2006.01) C07D 403/12 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2310382 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/4412 (2006.01) A61P 35/00 (2006.01) C07D 401/14 (2006.01) C07D 403/12 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2129377 B1 (19) NO NORGE (51) Int Cl. A61K 31/451 (2006.01) A61K 9/08 (2006.01) A61P 25/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.23
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2129377 B1 (19) NO NORGE (51) Int Cl. A61K 31/451 (2006.01) A61K 9/08 (2006.01) A61P 25/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.23
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2272978 B1 (19) NO NORGE (51) Int Cl. C12Q 1/68 (2006.01) Patentstyret (21) Oversettelse publisert 2012.08.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2272978 B1 (19) NO NORGE (51) Int Cl. C12Q 1/68 (2006.01) Patentstyret (21) Oversettelse publisert 2012.08.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 270722 B1 (19) NO NORGE (1) Int Cl. F21V 23/02 (06.01) F21S 8/02 (06.01) F21V 23/00 (06.01) Patentstyret (21) Oversettelse publisert 14.03. (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 270722 B1 (19) NO NORGE (1) Int Cl. F21V 23/02 (06.01) F21S 8/02 (06.01) F21V 23/00 (06.01) Patentstyret (21) Oversettelse publisert 14.03. (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2178851 B1 (19) NO NORGE (51) Int Cl. C07D 261/08 (2006.01) A61K 31/42 (2006.01) A61P 3/06 (2006.01) C07D 413/12 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2178851 B1 (19) NO NORGE (51) Int Cl. C07D 261/08 (2006.01) A61K 31/42 (2006.01) A61P 3/06 (2006.01) C07D 413/12 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2252286 B1 (19) NO NORGE (51) Int Cl. A61K 31/357 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.16 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2252286 B1 (19) NO NORGE (51) Int Cl. A61K 31/357 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.16 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2285808 B1 (19) NO NORGE (51) Int Cl. C07D 471/20 (2006.01) A61K 31/407 (2006.01) A61K 31/424 (2006.01) A61K 31/437 (2006.01) A61K 31/438 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2285808 B1 (19) NO NORGE (51) Int Cl. C07D 471/20 (2006.01) A61K 31/407 (2006.01) A61K 31/424 (2006.01) A61K 31/437 (2006.01) A61K 31/438 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2114970 B1 (19) NO NORGE (51) Int Cl. C07F 9/58 (2006.01) A61K 31/44 (2006.01) A61P 1/00 (2006.01) A61P 11/06 (2006.01) A61P 19/02 (2006.01) A61P
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2114970 B1 (19) NO NORGE (51) Int Cl. C07F 9/58 (2006.01) A61K 31/44 (2006.01) A61P 1/00 (2006.01) A61P 11/06 (2006.01) A61P 19/02 (2006.01) A61P
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2125711 B1 (19) NO NORGE (51) Int Cl. C07C 321/20 (2006.01) A61K 31/216 (2006.01) A61K 31/421 (2006.01) A61K 31/4402 (2006.01) A61K 31/495 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2125711 B1 (19) NO NORGE (51) Int Cl. C07C 321/20 (2006.01) A61K 31/216 (2006.01) A61K 31/421 (2006.01) A61K 31/4402 (2006.01) A61K 31/495 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2274977 B1 (19) NO NORGE (1) Int Cl. A01K 83/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2274977 B1 (19) NO NORGE (1) Int Cl. A01K 83/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 237066 B1 (19) NO NORGE (1) Int Cl. E06C 1/12 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 237066 B1 (19) NO NORGE (1) Int Cl. E06C 1/12 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2173868 B1 (19) NO NORGE (51) Int Cl. C12N 9/50 (2006.01) C07K 14/415 (2006.01) C12N 15/29 (2006.01) C12N 15/57 (2006.01) C12N 15/81 (2006.01) A23J
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2173868 B1 (19) NO NORGE (51) Int Cl. C12N 9/50 (2006.01) C07K 14/415 (2006.01) C12N 15/29 (2006.01) C12N 15/57 (2006.01) C12N 15/81 (2006.01) A23J
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2384729 B1 (19) NO NORGE (1) Int Cl. A61G /12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2384729 B1 (19) NO NORGE (1) Int Cl. A61G /12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 218466 B1 (19) NO NORGE (1) Int Cl. B67C 3/26 (06.01) B6D 47/ (06.01) B67C 7/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02. (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 218466 B1 (19) NO NORGE (1) Int Cl. B67C 3/26 (06.01) B6D 47/ (06.01) B67C 7/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02. (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2636033 B1 (19) NO NORGE (51) Int Cl. Patentstyret G09B 23/28 (2006.01) G09B 23/30 (2006.01) (21) Oversettelse publisert 2015.11.09 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2636033 B1 (19) NO NORGE (51) Int Cl. Patentstyret G09B 23/28 (2006.01) G09B 23/30 (2006.01) (21) Oversettelse publisert 2015.11.09 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 213696 B1 (19) NO NORGE (1) Int Cl. B23K 9/32 (2006.01) B23K 9/28 (2006.01) Patentstyret (21) Oversettelse publisert 2014.04.07 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 213696 B1 (19) NO NORGE (1) Int Cl. B23K 9/32 (2006.01) B23K 9/28 (2006.01) Patentstyret (21) Oversettelse publisert 2014.04.07 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2170890 B1 (19) NO NORGE (51) Int Cl. C07D 487/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.03.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2170890 B1 (19) NO NORGE (51) Int Cl. C07D 487/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.03.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2246321 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/135 (2006.01) C07C 211/42 (2006.01) Patentstyret (21) Oversettelse publisert 2011.12.12
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2246321 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/135 (2006.01) C07C 211/42 (2006.01) Patentstyret (21) Oversettelse publisert 2011.12.12
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 224294 B1 (19) NO NORGE (1) Int Cl. F16K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.10 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 224294 B1 (19) NO NORGE (1) Int Cl. F16K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.10 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP22342 (12) Oversettelse av europeisk patentskrift (11) NO/EP 22342 B1 (19) NO NORGE (1) Int Cl. F2D 23/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske Patentmyndighets
NO/EP22342 (12) Oversettelse av europeisk patentskrift (11) NO/EP 22342 B1 (19) NO NORGE (1) Int Cl. F2D 23/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 242166 B1 (19) NO NORGE (1) Int Cl. G06K 19/077 (06.01) G06K 19/06 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 242166 B1 (19) NO NORGE (1) Int Cl. G06K 19/077 (06.01) G06K 19/06 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2317621 B1 (19) NO NORGE (1) Int Cl. H02G 3/12 (06.01) Patentstyret (21) Oversettelse publisert 1.02.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2317621 B1 (19) NO NORGE (1) Int Cl. H02G 3/12 (06.01) Patentstyret (21) Oversettelse publisert 1.02.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22799 B1 (19) NO NORGE (1) Int Cl. A61K 31/23 (06.01) A61K 31/047 (06.01) A61K 31/231 (06.01) A61K 31/232 (06.01) A61K 31/3 (06.01) A61K 31/93 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22799 B1 (19) NO NORGE (1) Int Cl. A61K 31/23 (06.01) A61K 31/047 (06.01) A61K 31/231 (06.01) A61K 31/232 (06.01) A61K 31/3 (06.01) A61K 31/93 (06.01)
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2184425 B1 (19) NO NORGE (51) Int Cl. E05B 17/20 (2006.01) E05B 63/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.02.06 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2184425 B1 (19) NO NORGE (51) Int Cl. E05B 17/20 (2006.01) E05B 63/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.02.06 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 24012 B1 (19) NO NORGE (1) Int Cl. B2C 1/00 (2006.01) B2C 1/06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.12.22 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 24012 B1 (19) NO NORGE (1) Int Cl. B2C 1/00 (2006.01) B2C 1/06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.12.22 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 223094 B1 (19) NO NORGE (1) Int Cl. A43B 7/32 (06.01) A43B 7/12 (06.01) A43B 7/34 (06.01) A43B 13/12 (06.01) A43B 13/41 (06.01) B29D 3/14 (.01) Patentstyret
(12) Oversettelse av europeisk patentskrift (11) NO/EP 223094 B1 (19) NO NORGE (1) Int Cl. A43B 7/32 (06.01) A43B 7/12 (06.01) A43B 7/34 (06.01) A43B 13/12 (06.01) A43B 13/41 (06.01) B29D 3/14 (.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2128505 B1 (19) NO NORGE (51) Int Cl. F16L 9/12 (2006.01) F16L 3/14 (2006.01) F16L 11/127 (2006.01) F24F 13/02 (2006.01) H05F 3/02 (2006.01) Patentstyret
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2128505 B1 (19) NO NORGE (51) Int Cl. F16L 9/12 (2006.01) F16L 3/14 (2006.01) F16L 11/127 (2006.01) F24F 13/02 (2006.01) H05F 3/02 (2006.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2672278 B1 (19) NO NORGE (1) Int Cl. G01R 1/067 (2006.01) G01R 1/04 (2006.01) G01R 19/1 (2006.01) Patentstyret (21) Oversettelse publisert 201.04.20
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2672278 B1 (19) NO NORGE (1) Int Cl. G01R 1/067 (2006.01) G01R 1/04 (2006.01) G01R 19/1 (2006.01) Patentstyret (21) Oversettelse publisert 201.04.20
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 11438 B1 (19) NO NORGE (1) Int Cl. E04B 1/343 (06.01) B63B 29/02 (06.01) Patentstyret (21) Oversettelse publisert.02.23 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 11438 B1 (19) NO NORGE (1) Int Cl. E04B 1/343 (06.01) B63B 29/02 (06.01) Patentstyret (21) Oversettelse publisert.02.23 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 88493 B1 (19) NO NORGE (1) Int Cl. G06F 1/00 (06.01) H01L 23/34 (06.01) G06F 1/ (06.01) Patentstyret (21) Oversettelse publisert 13.04.22 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 88493 B1 (19) NO NORGE (1) Int Cl. G06F 1/00 (06.01) H01L 23/34 (06.01) G06F 1/ (06.01) Patentstyret (21) Oversettelse publisert 13.04.22 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2445326 B1 (19) NO NORGE (51) Int Cl. H05K 5/02 (2006.01) B43K 23/12 (2006.01) B43K 24/06 (2006.01) H01R 13/60 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2445326 B1 (19) NO NORGE (51) Int Cl. H05K 5/02 (2006.01) B43K 23/12 (2006.01) B43K 24/06 (2006.01) H01R 13/60 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2216387 B1 (19) NO NORGE (51) Int Cl. C10L 5/44 (2006.01) C10L 5/14 (2006.01) C10L 5/36 (2006.01) Patentstyret (21) Oversettelse publisert 2013.05.06
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2216387 B1 (19) NO NORGE (51) Int Cl. C10L 5/44 (2006.01) C10L 5/14 (2006.01) C10L 5/36 (2006.01) Patentstyret (21) Oversettelse publisert 2013.05.06
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2311023 B1 (19) NO NORGE (51) Int Cl. G09F 17/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2311023 B1 (19) NO NORGE (51) Int Cl. G09F 17/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 7044 B1 (19) NO NORGE (1) Int Cl. A61K 36/18 (06.01) A61K 33/04 (06.01) A61K 33/18 (06.01) A61K 33/ (06.01) A61K 36/22 (06.01) A61K 36/28 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 7044 B1 (19) NO NORGE (1) Int Cl. A61K 36/18 (06.01) A61K 33/04 (06.01) A61K 33/18 (06.01) A61K 33/ (06.01) A61K 36/22 (06.01) A61K 36/28 (06.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 19724 B1 (19) NO NORGE (1) Int Cl. B63H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 12.12. (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 19724 B1 (19) NO NORGE (1) Int Cl. B63H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 12.12. (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2613860 B1 (19) NO NORGE (51) Int Cl. B01D 15/18 (2006.01) C11B 3/10 (2006.01) C11C 1/00 (2006.01) C11C 1/08 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2613860 B1 (19) NO NORGE (51) Int Cl. B01D 15/18 (2006.01) C11B 3/10 (2006.01) C11C 1/00 (2006.01) C11C 1/08 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2097141 B1 (19) NO NORGE (51) Int Cl. A62B 35/00 (2006.01) Patentstyret (21) Oversettelse publisert 2013.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2097141 B1 (19) NO NORGE (51) Int Cl. A62B 35/00 (2006.01) Patentstyret (21) Oversettelse publisert 2013.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2231428 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) Patentstyret (21) Oversettelse publisert 12.11.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2231428 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) Patentstyret (21) Oversettelse publisert 12.11.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2491293 B1 (19) NO NORGE (1) Int Cl. F17C 3/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.2 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2491293 B1 (19) NO NORGE (1) Int Cl. F17C 3/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.2 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse
 Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse") (12) Oversettelse av europeisk patentskrift (11) NO/EP 217368 B2 (19) NO NORGE (1) Int Cl. B42D / (06.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse publisert.04. (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 217368 B2 (19) NO NORGE (1) Int Cl. B42D / (06.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse publisert.04. (80) Dato for
(86) Europeisk innleveringsdag
 Europeisk innleveringsdag") (12) Oversettelse av europeisk patentskrift (11) NO/EP 297978 B1 (19) NO NORGE (1) Int Cl. A41B 9/02 (06.01) Patentstyret (21) Oversettelse publisert 14.03.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 297978 B1 (19) NO NORGE (1) Int Cl. A41B 9/02 (06.01) Patentstyret (21) Oversettelse publisert 14.03.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP2770 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2770 B1 (19) NO NORGE (1) Int Cl. B23K 3/00 (06.01) C21D 6/00 (06.01) C21D 9/04 (06.01) C22C 38/00 (06.01) C22C 38/44 (06.01) Patentstyret
NO/EP2770 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2770 B1 (19) NO NORGE (1) Int Cl. B23K 3/00 (06.01) C21D 6/00 (06.01) C21D 9/04 (06.01) C22C 38/00 (06.01) C22C 38/44 (06.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2082973 B1 (19) NO NORGE (1) Int Cl. B6D 81/34 (2006.01) Patentstyret (21) Oversettelse publisert 2014.06.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2082973 B1 (19) NO NORGE (1) Int Cl. B6D 81/34 (2006.01) Patentstyret (21) Oversettelse publisert 2014.06.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") 1 3 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2207775 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/5377 (2006.01) A61P 3/06 (2006.01) C07D 401/14 (2006.01) C07D 413/14 (2006.01)
1 3 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2207775 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/5377 (2006.01) A61P 3/06 (2006.01) C07D 401/14 (2006.01) C07D 413/14 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2146022 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.11.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2146022 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.11.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2708433 B1 (19) NO NORGE (1) Int Cl. B61B 1/02 (2006.01) B61B 12/02 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.12 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2708433 B1 (19) NO NORGE (1) Int Cl. B61B 1/02 (2006.01) B61B 12/02 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.12 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22619 B1 (19) NO NORGE (1) Int Cl. B21D 1/4 (2006.01) B21K 21/04 (2006.01) F42B /02 (2006.01) F42B /188 (2006.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22619 B1 (19) NO NORGE (1) Int Cl. B21D 1/4 (2006.01) B21K 21/04 (2006.01) F42B /02 (2006.01) F42B /188 (2006.01) Patentstyret (21) Oversettelse publisert
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 23470 B1 (19) NO NORWAY (1) Int Cl. G01N 33/86 (2006.01) C12Q 1/6 (2006.01) G01N 33/49 (2006.01) G01N 33/43 (2006.01) Norwegian Industrial Property
(12) Translation of european patent specification (11) NO/EP 23470 B1 (19) NO NORWAY (1) Int Cl. G01N 33/86 (2006.01) C12Q 1/6 (2006.01) G01N 33/49 (2006.01) G01N 33/43 (2006.01) Norwegian Industrial Property
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2238877 B1 (19) NO NORGE (1) Int Cl. A47J 31/08 (06.01) Patentstyret (21) Oversettelse publisert 13.03.11 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2238877 B1 (19) NO NORGE (1) Int Cl. A47J 31/08 (06.01) Patentstyret (21) Oversettelse publisert 13.03.11 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2096736 B1 (19) NO NORGE (1) Int Cl. H02K 1/32 (2006.01) H02K 3/24 (2006.01) H02K 9/00 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2096736 B1 (19) NO NORGE (1) Int Cl. H02K 1/32 (2006.01) H02K 3/24 (2006.01) H02K 9/00 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0
(12) Translation of european patent specification
 Translation of european patent specification") NO/EP20 (12) Translation of european patent specification (11) NO/EP 20 B1 (19) NO NORWAY (1) Int Cl. C07K 16/28 (2006.01) A61P 3/00 (2006.01) A61P 37/00 (2006.01) Norwegian Industrial Property Office
NO/EP20 (12) Translation of european patent specification (11) NO/EP 20 B1 (19) NO NORWAY (1) Int Cl. C07K 16/28 (2006.01) A61P 3/00 (2006.01) A61P 37/00 (2006.01) Norwegian Industrial Property Office
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP28769 (12) Oversettelse av europeisk patentskrift (11) NO/EP 28769 B1 (19) NO NORGE (1) Int Cl. F17D 1/18 (06.01) F16L 3/00 (06.01) Patentstyret (21) Oversettelse publisert 1.04. (80) Dato for Den
NO/EP28769 (12) Oversettelse av europeisk patentskrift (11) NO/EP 28769 B1 (19) NO NORGE (1) Int Cl. F17D 1/18 (06.01) F16L 3/00 (06.01) Patentstyret (21) Oversettelse publisert 1.04. (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 20789 B1 (19) NO NORGE (1) Int Cl. B61D 1/00 (06.01) B61D 17/ (06.01) B61D 23/00 (06.01) Patentstyret (21) Oversettelse publisert 12.06.04 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 20789 B1 (19) NO NORGE (1) Int Cl. B61D 1/00 (06.01) B61D 17/ (06.01) B61D 23/00 (06.01) Patentstyret (21) Oversettelse publisert 12.06.04 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2213923 B1 (19) NO NORGE (1) Int Cl. F16L 19/02 (06.01) F16L 19/028 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2213923 B1 (19) NO NORGE (1) Int Cl. F16L 19/02 (06.01) F16L 19/028 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 26098 B1 (19) NO NORWAY (1) Int Cl. C07K 14/08 (06.01) C12Q 1/70 (06.01) Norwegian Industrial Property Office (21) Translation Published 16..24
(12) Translation of european patent specification (11) NO/EP 26098 B1 (19) NO NORWAY (1) Int Cl. C07K 14/08 (06.01) C12Q 1/70 (06.01) Norwegian Industrial Property Office (21) Translation Published 16..24
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 21181 B1 (19) NO NORGE (1) Int Cl. F16L 2/00 (2006.01) F16L 33/26 (2006.01) H01P 1/04 (2006.01) Patentstyret (21) Oversettelse publisert 2013.10.28
(12) Oversettelse av europeisk patentskrift (11) NO/EP 21181 B1 (19) NO NORGE (1) Int Cl. F16L 2/00 (2006.01) F16L 33/26 (2006.01) H01P 1/04 (2006.01) Patentstyret (21) Oversettelse publisert 2013.10.28
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2261144 B1 (19) NO NORGE (1) Int Cl. B6G 21/00 (06.01) B6G 21/08 (06.01) Patentstyret (21) Oversettelse publisert 13.07.08 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2261144 B1 (19) NO NORGE (1) Int Cl. B6G 21/00 (06.01) B6G 21/08 (06.01) Patentstyret (21) Oversettelse publisert 13.07.08 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2146836 B1 (19) NO NORGE (1) Int Cl. A47G 9/ (06.01) B26D 3/00 (06.01) B26D 3/28 (06.01) B29C 44/6 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2146836 B1 (19) NO NORGE (1) Int Cl. A47G 9/ (06.01) B26D 3/00 (06.01) B26D 3/28 (06.01) B29C 44/6 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2477830 B1 (19) NO NORGE (1) Int Cl. B60K 1/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2477830 B1 (19) NO NORGE (1) Int Cl. B60K 1/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse
 Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2175588 B2 (19) NO NORGE (51) Int Cl. H04L 12/14 (2006.01) H04L 29/08 (2006.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2175588 B2 (19) NO NORGE (51) Int Cl. H04L 12/14 (2006.01) H04L 29/08 (2006.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22442 B1 (19) NO NORGE (1) Int Cl. G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13..28 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22442 B1 (19) NO NORGE (1) Int Cl. G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13..28 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2243894 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2243894 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 222267 B1 (19) NO NORWAY (1) Int Cl. A01N 1/02 (06.01) A23G 9/38 (06.01) C07K 14/43 (06.01) Norwegian Industrial Property Office (21) Translation
(12) Translation of european patent specification (11) NO/EP 222267 B1 (19) NO NORWAY (1) Int Cl. A01N 1/02 (06.01) A23G 9/38 (06.01) C07K 14/43 (06.01) Norwegian Industrial Property Office (21) Translation
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 261673 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) B60H 1/00 (06.01) Patentstyret (21) Oversettelse publisert 1.01.12 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 261673 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) B60H 1/00 (06.01) Patentstyret (21) Oversettelse publisert 1.01.12 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2236434 B1 (19) NO NORGE (1) Int Cl. B6D 77/04 (06.01) B6D 77/06 (06.01) Patentstyret (21) Oversettelse publisert 11.12.19 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2236434 B1 (19) NO NORGE (1) Int Cl. B6D 77/04 (06.01) B6D 77/06 (06.01) Patentstyret (21) Oversettelse publisert 11.12.19 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2264391 B1 (19) NO NORGE (1) Int Cl. F27D 3/1 (2006.01) C21B 7/12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.11.18 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2264391 B1 (19) NO NORGE (1) Int Cl. F27D 3/1 (2006.01) C21B 7/12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.11.18 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 240126 B1 (19) NO NORGE (1) Int Cl. C07D 211/62 (06.01) A61K 31/16 (06.01) A61K 31/44 (06.01) A61K 31/0 (06.01) A61K 31/06 (06.01) C07D 7/277 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 240126 B1 (19) NO NORGE (1) Int Cl. C07D 211/62 (06.01) A61K 31/16 (06.01) A61K 31/44 (06.01) A61K 31/0 (06.01) A61K 31/06 (06.01) C07D 7/277 (06.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2244923 B1 (19) NO NORGE (1) Int Cl. B61K 9/ (2006.01) Patentstyret (21) Oversettelse publisert 2013.09.30 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2244923 B1 (19) NO NORGE (1) Int Cl. B61K 9/ (2006.01) Patentstyret (21) Oversettelse publisert 2013.09.30 (80) Dato for Den Europeiske Patentmyndighets
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 21847 B1 (19) NO NORGE (1) Int Cl. F24F 7/08 (06.01) F24F 11/04 (06.01) F24F 12/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 21847 B1 (19) NO NORGE (1) Int Cl. F24F 7/08 (06.01) F24F 11/04 (06.01) F24F 12/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2216871 B1 (19) NO NORGE (51) Int Cl. H02J 7/00 (2006.01) H01R 13/22 (2006.01) H01R 13/62 (2006.01) Patentstyret (21) Oversettelse publisert 2014.09.08
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2216871 B1 (19) NO NORGE (51) Int Cl. H02J 7/00 (2006.01) H01R 13/22 (2006.01) H01R 13/62 (2006.01) Patentstyret (21) Oversettelse publisert 2014.09.08
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2292031 B1 (19) NO NORGE (51) Int Cl. H04W 8/26 (2009.01) Patentstyret (21) Oversettelse publisert 2013.03.25 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2292031 B1 (19) NO NORGE (51) Int Cl. H04W 8/26 (2009.01) Patentstyret (21) Oversettelse publisert 2013.03.25 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2231500 B1 (19) NO NORGE (51) Int Cl. B66F 9/00 (2006.01) B60P 1/02 (2006.01) B60P 3/022 (2006.01) B62B 3/065 (2006.01) B66D 1/00 (2006.01) B66F 9/06
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2231500 B1 (19) NO NORGE (51) Int Cl. B66F 9/00 (2006.01) B60P 1/02 (2006.01) B60P 3/022 (2006.01) B62B 3/065 (2006.01) B66D 1/00 (2006.01) B66F 9/06
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 08940 B1 (19) NO NORGE (1) Int Cl. B6D 2/2 (06.01) A47G 19/34 (06.01) B6D 83/06 (06.01) G01F 11/26 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 08940 B1 (19) NO NORGE (1) Int Cl. B6D 2/2 (06.01) A47G 19/34 (06.01) B6D 83/06 (06.01) G01F 11/26 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2246634 B1 (19) NO NORGE (1) Int Cl. F24F 11/02 (2006.01) F24F 3/044 (2006.01) F24F 11/00 (2006.01) F24F 13/04 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2246634 B1 (19) NO NORGE (1) Int Cl. F24F 11/02 (2006.01) F24F 3/044 (2006.01) F24F 11/00 (2006.01) F24F 13/04 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 203638 B1 (19) NO NORGE (1) Int Cl. A61K 9/00 (2006.01) A61K 31/382 (2006.01) A61K 31/498 (2006.01) A61K 31/3 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 203638 B1 (19) NO NORGE (1) Int Cl. A61K 9/00 (2006.01) A61K 31/382 (2006.01) A61K 31/498 (2006.01) A61K 31/3 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 1974881 B1 (19) NO NORGE (1) Int Cl. B27B 19/00 (06.01) A61B 17/14 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 1974881 B1 (19) NO NORGE (1) Int Cl. B27B 19/00 (06.01) A61B 17/14 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 17118 B1 (19) NO NORGE (1) Int Cl. B60M 1/06 (06.01) B60M 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.09.29 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 17118 B1 (19) NO NORGE (1) Int Cl. B60M 1/06 (06.01) B60M 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.09.29 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP918 (12) Oversettelse av europeisk patentskrift (11) NO/EP 918 B1 (19) NO NORGE (1) Int Cl. H02J 7/00 (06.01) Patentstyret (21) Oversettelse publisert 14.02.03 (80) Dato for Den Europeiske Patentmyndighets
NO/EP918 (12) Oversettelse av europeisk patentskrift (11) NO/EP 918 B1 (19) NO NORGE (1) Int Cl. H02J 7/00 (06.01) Patentstyret (21) Oversettelse publisert 14.02.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2217383 B1 (19) NO NORGE (1) Int Cl. B0B 12/00 (06.01) B0B 11/00 (06.01) G01F 11/02 (06.01) G01F 1/07 (06.01) G07C 3/04 (06.01) Patentstyret (21)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2217383 B1 (19) NO NORGE (1) Int Cl. B0B 12/00 (06.01) B0B 11/00 (06.01) G01F 11/02 (06.01) G01F 1/07 (06.01) G07C 3/04 (06.01) Patentstyret (21)
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2616248 B1 (19) NO NORWAY (51) Int Cl. G03C 11/08 (2006.01) B41M 7/00 (2006.01) G02B 1/11 (2015.01) G03C 11/14 (2006.01) Norwegian Industrial
(12) Translation of european patent specification (11) NO/EP 2616248 B1 (19) NO NORWAY (51) Int Cl. G03C 11/08 (2006.01) B41M 7/00 (2006.01) G02B 1/11 (2015.01) G03C 11/14 (2006.01) Norwegian Industrial
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 263813 B1 (19) NO NORWAY (1) Int Cl. C07K 16/18 (2006.01) A61K 39/39 (2006.01) A61P 37/06 (2006.01) Norwegian Industrial Property Office (21)
(12) Translation of european patent specification (11) NO/EP 263813 B1 (19) NO NORWAY (1) Int Cl. C07K 16/18 (2006.01) A61K 39/39 (2006.01) A61P 37/06 (2006.01) Norwegian Industrial Property Office (21)
PATENTKRAV. og en første lett kjede og en andre lett kjede, hvor første og andre lette kjeder er forskjellige.
 1 PATENTKRAV 1. Isolert monoklonalt antistoff som bærer en forskjellig spesifisitet i hvert kombinasjonssete og som består av to kopier av et enkelt tung kjede polypeptid og en første lett kjede og en
1 PATENTKRAV 1. Isolert monoklonalt antistoff som bærer en forskjellig spesifisitet i hvert kombinasjonssete og som består av to kopier av et enkelt tung kjede polypeptid og en første lett kjede og en
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2190 B1 (19) NO NORGE (1) Int Cl. C23C 14/28 (06.01) C23C 14/0 (06.01) Patentstyret (21) Oversettelse publisert 12.04. (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2190 B1 (19) NO NORGE (1) Int Cl. C23C 14/28 (06.01) C23C 14/0 (06.01) Patentstyret (21) Oversettelse publisert 12.04. (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2391650 B1 (19) NO NORGE (51) Int Cl. C07K 16/24 (2006.01) A61K 39/00 (2006.01) A61K 39/395 (2006.01) Patentstyret (21) Oversettelse publisert 2015.03.09
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2391650 B1 (19) NO NORGE (51) Int Cl. C07K 16/24 (2006.01) A61K 39/00 (2006.01) A61K 39/395 (2006.01) Patentstyret (21) Oversettelse publisert 2015.03.09
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 9863 B1 (19) NO NORGE (1) Int Cl. E04B 2/96 (06.01) Patentstyret (21) Oversettelse publisert 13.09.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 9863 B1 (19) NO NORGE (1) Int Cl. E04B 2/96 (06.01) Patentstyret (21) Oversettelse publisert 13.09.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 216340 B1 (19) NO NORGE (1) Int Cl. B60C 11/11 (06.01) B60C 11/03 (06.01) B60C 11/12 (06.01) Patentstyret (21) Oversettelse publisert 12.12.03 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 216340 B1 (19) NO NORGE (1) Int Cl. B60C 11/11 (06.01) B60C 11/03 (06.01) B60C 11/12 (06.01) Patentstyret (21) Oversettelse publisert 12.12.03 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 196721 B1 (19) NO NORGE (1) Int Cl. A61L 9/04 (06.01) A61B 19/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02.06 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 196721 B1 (19) NO NORGE (1) Int Cl. A61L 9/04 (06.01) A61B 19/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02.06 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2289870 B1 (19) NO NORGE (51) Int Cl. C07C 231/24 (2006.01) C07C 237/32 (2006.01) Patentstyret (21) Oversettelse publisert 2013.03.11 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2289870 B1 (19) NO NORGE (51) Int Cl. C07C 231/24 (2006.01) C07C 237/32 (2006.01) Patentstyret (21) Oversettelse publisert 2013.03.11 (80) Dato for
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 237122 B1 (19) NO NORWAY (1) Int Cl. G06K 7/08 (06.01) G06F 3/044 (06.01) G06F 3/0488 (13.01) G06K 19/067 (06.01) Norwegian Industrial Property
(12) Translation of european patent specification (11) NO/EP 237122 B1 (19) NO NORWAY (1) Int Cl. G06K 7/08 (06.01) G06F 3/044 (06.01) G06F 3/0488 (13.01) G06K 19/067 (06.01) Norwegian Industrial Property
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 238426 B1 (19) NO NORGE (1) Int Cl. G01S 1/68 (06.01) B63C 9/32 (06.01) F41B 13/00 (06.01) F41B 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 238426 B1 (19) NO NORGE (1) Int Cl. G01S 1/68 (06.01) B63C 9/32 (06.01) F41B 13/00 (06.01) F41B 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2451844 B1 (19) NO NORWAY (51) Int Cl. C07K 16/28 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2015.09.21 (80)
(12) Translation of european patent specification (11) NO/EP 2451844 B1 (19) NO NORWAY (51) Int Cl. C07K 16/28 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2015.09.21 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2372667 B1 (19) NO NORGE (1) Int Cl. G08G 1/01 (06.01) G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13.01.21 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2372667 B1 (19) NO NORGE (1) Int Cl. G08G 1/01 (06.01) G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13.01.21 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2148223 B1 (19) NO NORGE (1) Int Cl. G01V 3/ (06.01) G01V 3/24 (06.01) Patentstyret (21) Oversettelse publisert 13.03.04 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2148223 B1 (19) NO NORGE (1) Int Cl. G01V 3/ (06.01) G01V 3/24 (06.01) Patentstyret (21) Oversettelse publisert 13.03.04 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 264739 B1 (19) NO NORGE (1) Int Cl. A61F 13/00 (06.01) A61B 17/132 (06.01) A61F 13/64 (06.01) A61F 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 264739 B1 (19) NO NORGE (1) Int Cl. A61F 13/00 (06.01) A61B 17/132 (06.01) A61F 13/64 (06.01) A61F 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2383703 B1 (19) NO NORGE (1) Int Cl. G07B 1/06 (11.01) G08G 1/017 (06.01) Patentstyret (21) Oversettelse publisert 13.01.21 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2383703 B1 (19) NO NORGE (1) Int Cl. G07B 1/06 (11.01) G08G 1/017 (06.01) Patentstyret (21) Oversettelse publisert 13.01.21 (80) Dato for Den Europeiske
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 211778 B1 (19) NO NORWAY (1) Int Cl. A61K 38/16 (06.01) C07K 7/08 (06.01) Norwegian Industrial Property Office (21) Translation Published 1.09.07
(12) Translation of european patent specification (11) NO/EP 211778 B1 (19) NO NORWAY (1) Int Cl. A61K 38/16 (06.01) C07K 7/08 (06.01) Norwegian Industrial Property Office (21) Translation Published 1.09.07
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2003466 B1 (19) NO NORGE (1) Int Cl. G01S /02 (2010.01) Patentstyret (21) Oversettelse publisert 2014.07.14 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2003466 B1 (19) NO NORGE (1) Int Cl. G01S /02 (2010.01) Patentstyret (21) Oversettelse publisert 2014.07.14 (80) Dato for Den Europeiske Patentmyndighets