(12) Oversettelse av europeisk patentskrift
|
|
|
- Lennart Markussen
- 8 år siden
- Visninger:
Transkript
1 (12) Oversettelse av europeisk patentskrift (11) NO/EP B1 (19) NO NORGE (1) Int Cl. A61K 39/39 (06.01) A61P 3/00 (06.01) Patentstyret (21) Oversettelse publisert (80) Dato for Den Europeiske Patentmyndighets publisering av det meddelte patentet (86) Europeisk søknadsnr (86) Europeisk innleveringsdag (87) Den europeiske søknadens Publiseringsdato () Prioritet , US, P , US, 9796 P (84) Utpekte stater AT BE BG CH CY CZ DE DK EE ES FI FR GB GR HR HU IE IS IT LI LT LU LV MC MT NL NO PL PT RO SE SI SK TR Utpekte samarbeidende stater AL BA MK RS (73) Innehaver Merrimack Pharmaceuticals, Inc., One Kendall Square Bldg. 700, 2nd Floor, Cambridge, MA 02139, USA (72) Oppfinner SCHOEBERL, Birgit, 324 Windsor Street, Cambridge, MA 02141, USA NIELSEN, Ulrik, 18 Thomas Street, Quincy, MA 02169, USA FELDHAUS, Michael, P.O. Box 1888, Grantham, NH 0373, USA BUCKLER, David, 6 Erick Court, Chester, NJ 079, USA MURUGANANDAM, Arumugam, 364 Rainbow DriveOpposite to Wipro Corporate OfficeSarjapur Rd., Doddakannahalli,Bangalore 6003, India (74) Fullmektig Hjerrild & Levin A/S, Vedbæk Strandvej 341, DK-290 VEDBÆK, Danmark (4) Benevnelse Antistoffer med ERBB3 og anvendelse derav (6) Anførte publikasjoner EP-A , US-A , WO-A-06/01738, WO-A-06/0706 WO-A-06/0919, WO-A-07/077028, WO-A-97/388 BALINT R F ET AL: "ANTIBODY ENGINEERING BY PARSIMONIOUS MUTAGENESIS" GENE, ELSEVIER, AMSTERDAM, NL, vol. 137, no. 1, 27 December 1993 ( ), pages 9-118, XP ISSN: , DAVIES J ET AL: "Affinity improvement of single antibody VH domains: residues in all three hypervariable regions affect antigen binding" IMMUNOTECHNOLOGY, ELSEVIER SCIENCE PUBLISHERS BV, NL, vol. 2, no. 3, 1 September 1996 ( ), pages , XP ISSN: HOLT L J ET AL: "Domain antibodies: proteins for therapy" TRENDS IN BIOTECHNOLOGY, ELSEVIER PUBLICATIONS, CAMBRIDGE, GB, vol. 21, no. 11, 1 November 03 ( ), pages , XP ISSN: , LITTLE M ET AL: "Of mice and men: hybridoma and recombinant antibodies" IMMUNOLOGY TODAY, ELSEVIER PUBLICATIONS, CAMBRIDGE, GB, vol. 21, no. 8, 1 August 00 ( ), pages , XP ISSN: LOEE H ET AL: "ISOLATION AND CHARACTERIZATION OF FOUR ALTERNATE C-ERBB3 TRANSCRIPTS EXPRESSED IN OVARIAN CARCINOMA-DERIVED CELL LINES AND NORMAL HUMAN TISSUES" ONCOGENE, NATURE PUBLISHING GROUP, GB BASINGSTOKE, HANTS, vol. 16, no. 2, 2 June 1998 ( ), pages , XP00877 ISSN: , PINKAS-KRAMARSKI, R. ET AL: "The oncogenic ErbB- 2/ErbB-3 heterodimer is a surrogate receptor of the epidermal growth factor and betacellulin.", ONCOGENE, vol. 16, 1998, pages , XP006823,
 Utpekte stater AT BE BG CH CY CZ DE DK EE ES FI FR GB GR HR HU IE IS IT LI LT LU LV MC MT NL NO PL PT RO SE SI SK TR Utpekte samarbeidende stater AL BA MK RS (73) Innehaver")
2 1 Beskrivelse Oppfinnelsens bakgrunn [0001] ErbB/HER underfamilien av polypeptid-vekstfaktor-reseptorer inkluderer epidermal vekstfaktor (EGF)- reseptor (EGFR, ErbB1/HER1), det neu onkogene produktet (ErbB2/HER2), og de nyligere identifiserte ErbB3/HER3 og ErbB4/HER4 reseptorproteinene (se, f.eks. Hynes et. al. (1994) Biochim. Biophys. Acta Rev Cancer, ). Hver av disse reseptorene er forutsagt å bestå av et ekstracellulært ligand-bindende domene, et membranspennende domene, et cytosolisk protein tyrosin-kinase (PTK) domene og et C-terminalt fosforyleringsdomene (se, f.eks. Kim et al. (1998) Biochem. 334 J., ). [0002] Eksperimenter in vitro har indikert at proteintyrosinkinase-aktiviteten til ErbB3 proteinet dempes betydelig 1 2 i forhold til den av andre ErbB/HER familiemedlemmer og denne dempingen har blitt tilskrevet, delvis, til forekomsten av ikke-konservative aminosyresubstitusjoner i det forutsagte katalytiske domenet av ErbB3 (se, f.eks. Guy et al. (1994) Proc. Natl. Acad. Sci. USA. 91, ; Sierke et al. (1997) Biochem. 322 J., ). Imidlertid har ErbB3 proteinet blitt vist å bli fosforylert i en rekke cellulære sammenhenger. For eksempel blir ErbB3 konstitutivt fosforylert på tyrosin-rester i et undersett av humane brystkreftcellelinjer som overuttrykker dette proteinet (se, f.eks. Kraus et al. (1993) Proc. Natl. Acad. Sci. USA. 90, , og Kim et al. Supra, se, også, Schaefer et al. (06) Neoplasia 8 (7): og Schaefer et al. Cancer Res (04) 64 (): ). [0003] Selv om rollen for ErbB3 i kreft har blitt utforsket (se, f.eks. Horst et al.(0) 11, 19-27; Xue et al. (06) Cancer Res. 66, ), forblir ErbB3 i stor grad ikke verdsatt som et mål for klinisk intervensjon. Dagens immunoterapier fokuserer primært på å inhibere virkningen av ErbB2 og, spesielt, heterodimerisering av ErbB2/ErbB3 komplekser (se, f.eks. Sliwkowski et al. (1994) J. Biol. Chem. 269 (): (1994)). Følgelig er det et formål med foreliggende oppfinnelse å tilveiebringe forbedrede immunoterapier som effektivt inhiberer ErbB3 signalisering, og kan brukes for å behandle og diagnostisere en rekke krefttyper. [0004] EP-A-1283 beskriver anvendelsen av en inhibitor av ErbB3 aktivitet, særlig et anti-erbb3-antistoff for diagnosen, forhindringen eller behandlingen av hyperproliferative sykdommer. [000] WO 06/0919 beskriver fremgangsmåter for å forbedre evnen til spesifikk binding av bispesifikke bindende sammensetninger. I én utførelsesform er målmolekylet ErbB3. 3 [0006] WO 97/388 beskriver antistoffer som binder til ErbB3 protein og videre har en evne til å redusere heregulin-indusert dannelse av et ErbB2-ErbB3 proteinkompleks, for å øke bindingsaffiniteten av heregulin for ErbB3 proteinet og/eller for å redusere heregulin-indusert ErbB2-aktivering i en celle som uttrykker ErbB2 og ErbB3.
 Biochem. 334 J., 189-19).")
3 2 [0007] Lee og Maihle (Oncogene (1998) 16: ) beskriver isoleringen og karakteriseringen av fire alternative c-erbb3 transkripter uttrykt i ovariekarsinom-avledede cellelinjer og normale humane vev. [0008] Pinkas-Kramarski et al (Oncogene (1998) 16: ) viser at de onkogene ErbB2/ErbB3 heterodimerene er en surrogatreseptor av den epidermale vekstfaktoren og betacellulin. Oppsummering av oppfinnelsen 1 [0009] Foreliggende oppfinnelse tilveiebringer en ny klasse av monoklonale antistoffer som binder seg til ErbB3 reseptoren og inhiberer ulike ErbB3 funksjoner. Oppfinnelsen tilveiebringer et antistoff eller antigenbindende del derav som binder til human ErbB3 og omfatter: (a) en tung kjede variabel region, hvori CDR1en omfatter aminosyresekvensen ifølge SEKV ID NR: 7, CDR2en omfatter aminosyresekvensen ifølge SEKV ID NR: 8 og CDR3en omfatter aminosyresekvensen ifølge SEKV ID NR: 9, og en lett kjede variabel region, hvori CDR1en omfatter aminosyresekvensen ifølge SEKV ID NR:, CDR2en omfatter aminosyresekvensen ifølge SEKV ID NR: 11, og CDR3en omfatter aminosyresekvensen ifølge SEKV ID NR: 12; eller (b) en tung kjede variabel region, hvori CDR1en, CDR2en og CDR3en omfatter aminosyresekvenser som er minst 90 % identiske med CDR1, CDR2 og CDR3 aminosyresekvensene definert i (a), og en lett kjede variabel region, hvori CDR1en, CDR2en og CDR3en omfatter aminosyresekvenser som er minst 90 % identiske med CDR1, CDR2 og CDR3 aminosyresekvensene definert i (a) og hvori antistoffet eller den antigenbindende delen derav, er i stand til å inhibere EGF-lignende ligandmediert fosforylering av ErbB3. 2 [00] Som beskrevet heri, inkluderer EGF-lignende ligander EGF, TGF-α, betacellulin, heparin-bindende epidermal vekstfaktor, biregulin og amfiregulin, som binder til EGFR og induserer dimerisering av EGFR med ErbB3. Denne dimeriseringen fører i sin tur til fosforylering av ErbB3, og aktiverer signalisering gjennom reseptoren. Monoklonale antistoffer ifølge foreliggende oppfinnelse er således nyttige for å behandle og diagnostisere en rekke krefttyper forbundet med ErbB3-mediert cellulær signalisering. 3 [0011] I en annen utførelsesform er antistoffene videre kjennetegnet ved én eller flere av de følgende egenskapene: (i) inhibering av ErbB3-ligandmediert signalisering, inkludert signalisering mediert ved binding av ErbB3-ligander, så som heregulin, epiregulin, epigen og BIR, til ErbB3, (ii) inhibering av proliferasjon av celler som uttrykker ErbB3, (iii) evnen til å redusere nivåer av ErbB3 på celleoverflaten (f.eks. ved å indusere internalisering av ErbB3), (iv) inhibering av VEGF-sekresjon fra celler som uttrykker ErbB3, (v) inhibering av migreringen av celler som uttrykker ErbB3, (vi) inhibering av sfæroidevekst av celler som uttrykker ErbB3, og/eller (vii) binding til en epitope lokalisert på domene I (rester -9) av ErbB3, for eksempel, en epitope som involverer eller som spenner over restene -2 av aminosyresekvensen av ErbB3.
![Oppsummering av oppfinnelsen 1 [0009] Foreliggende oppfinnelse tilveiebringer en ny klasse av monoklonale antistoffer som binder seg til ErbB3 reseptoren og inhiberer ulike ErbB3 funksjoner.](/docs-images/46/1534177/images/page_3.jpg "Oppfinnelsen tilveiebringer et antistoff eller antigenbindende del derav som binder til human ErbB3 og omfatter: (a) en tung kjede variabel region, hvori CDR1en omfatter aminosyresekvensen ifølge")
4 3 [0012] Spesielle monoklonale antistoffer og antigenbindende deler derav ifølge foreliggende oppfinnelse utviser en K D på 0 nm eller mindre, som målt ved en overflate-plasmonresonansanalyse eller en cellebindingsanalyse [0013] Spesielle monoklonale antistoffer og antigenbindende deler derav ifølge foreliggende oppfinnelse inkluderer en tung kjede variabel region som omfatter en aminosyresekvens som er minst 90 % (f.eks. 9 %, 96 %, 97 %, 98 % eller 99 %) identisk med tung kjede variabel region aminosyresekvensen fremlagt i SEKV ID NR: 1, SEKV ID NR: 3, SEKV ID NR:, SEKV ID NR: 3 eller SEKV ID NR: 37. Andre spesielle monoklonale antistoffer og antigenbindende deler derav ifølge foreliggende oppfinnelse inkluderer en lett kjede variabel region som omfatter en aminosyresekvens som er minst 90 % (f.eks. 9 %, 96 %, 97 %, 98 % eller 99 %) identisk med lett kjede variabel region aminosyresekvensen fremlagt i SEKV ID NR: 2, SEKV ID NR: 4, SEKV ID NR: 6, SEKV ID NR: 36 eller SEKV ID NR: 38. Antistoffene kan også inkludere begge de tidligere nevnte tung kjede og lett kjede variable regionene. 1 [0014] Søknaden beskriver også antistoffer og antigenbindende deler derav som inkluderer én eller flere CDRsekvenser valgt fra en tung kjede variabel region CDR1 som omfatter SEKV ID NR: 13; en tung kjede variabel CDR2 region som omfatter SEKV ID NR: 14, en tung kjede variabel region CDR3 som omfatter SEKV ID NR: 1, en lett kjede variabel region CDR1 som omfatter SEKV ID NR: 16, en lett kjede variabel region CDR2 som omfatter SEKV ID NR: 17, en lett kjede variabel region CDR3 som omfatter SEKV ID NR: 18, og kombinasjoner derav. [001] Enda andre beskrevne antistoffer og antigenbindende deler derav inkluderer: én eller flere CDR-sekvenser valgt fra en tung kjede variabel region CDR1 som omfatter SEKV ID NR: 19; en tung kjede variabel region CDR2 som omfatter SEKV ID NR: ; en tung kjede variabel region CDR3 som omfatter SEKV ID NR: 21, en lett kjede variabel region CDR1 som omfatter SEKV ID NR: 22, en lett kjede variabel region CDR2 som omfatter SEKV ID NR: 23, en lett kjede variabel region CDR3 som omfatter SEKV ID NR: 24, og kombinasjoner derav. 2 3 [0016] Enda andre beskrevne antistoffer og antigenbindende deler derav inkluderer, eller én eller flere CDRsekvenser valgt fra en tung kjede variabel region CDR1 som omfatter SEKV ID NR: 39; en tung kjede variabel region CDR2 som omfatter SEKV ID NR: 40, en tung kjede variabel region CDR3 som omfatter SEKV ID NR: 41, en lett kjede variabel region CDR1 som omfatter SEKV ID NR: 42, en lett kjede variabel region CDR2 som omfatter SEKV ID NR: 43, en lett kjede variabel region CDR3 som omfatter SEKV ID NR: 44, og kombinasjoner derav. [0017] Enda andre antistoffer og antigenbindende deler derav som er beskrevet inkluderer, eller én eller flere CDR-sekvenser valgt fra en tung kjede variabel region CDR1 som omfatter SEKV ID NR: 4; en tung kjede variabel region CDR2 som omfatter SEKV ID NR: 46, en tung kjede variabel region CDR3 som omfatter SEKV ID NR: 47, en lett kjede variabel region CDR1 som omfatter SEKV ID NR: 48, en lett kjede variabel region CDR2 som omfatter SEKV ID NR: 49; en lett kjede variabel region CDR3 som omfatter SEKV ID NR: 0, og kombinasjoner derav. [0018] I én utførelsesform er antistoffene og antistoffdelene derav fullstendig humane (dvs. inneholder human CDR og rammeverksekvenser). Spesielle humane antistoffer ifølge oppfinnelsen inkluderer de som har en tung
 identisk med tung kjede variabel region aminosyresekvensen fremlagt i SEKV ID NR: 1, SEKV ID NR: 3, SEKV ID NR:, SEKV ID NR: 3 eller SEKV ID NR: 37.")
5 4 kjede variabel region som er fra et humant VH3 kimlinje-gen, og/eller en lett kjede variabel region fra humant VL2 kimlinje-gen. [0019] Også omfattet ved foreliggende oppfinnelse er monoklonale antistoffer og deler derav som binder til de samme eller overlappende epitoper bundet ved et hvilket som helst av antistoffene eller deler derav beskrevet heri (for eksempel en epitope lokalisert på domene I av ErbB3, så som en epitope som involverer eller spenner, restene -2 av aminosyresekvensen av ErbB3). Antistoffer som har den samme aktiviteten som antistoffene beskrevet heri, for eksempel antistoffer, som har den samme sekvensen som Ab #6, er også omfattet ved foreliggende oppfinnelse. Spesielt tilveiebringer oppfinnelsen antistoffet eller antigenbindende del derav ifølge oppfinnelsen, som omfatter en tung kjede variabel region som omfatter aminosyresekvensen ifølge SEKV ID NR: 1 og en lett kjede variabel region som omfatter aminosyresekvensen ifølge SEKV ID NR: 2. 1 [00] Antistoffer ifølge foreliggende oppfinnelse inkluderer alle kjente former av antistoffer og andre proteinstativer med antistoff-lignende egenskaper. For eksempel kan antistoffet være et humant antistoff, et humanisert antistoff, et bispesifikt antistoff, et kimært antistoff eller et proteinstativ med antistoff-lignende egenskaper, så som fibronektin eller Ankyrin repeterende enheter. Antistoffet kan også være et Fab, Fab'2, scfv eller et domene-antistoff. Antistoffet også kan ha en hvilken som helst av de følgende isotyper: IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgAsec, IgD og IgE. 2 [0021] I enda en annen utførelsesform tilveiebringer foreliggende oppfinnelse videre sammensetninger som omfatter kombinasjoner av antistoffer eller antigenbindende deler beskrevet heri, formulert med en akseptabel bærer og/eller adjuvans. I en spesiell utførelsesform omfatter sammensetningen to eller flere antistoffer som binder forskjellige epitoper på ErbB3 eller antistoffer beskrevet heri kombinert med anti-kreft antistoffer som ikke binder ErbB3. [0022] I enda en annen utførelsesform tilveiebringer foreliggende oppfinnelse en isolert nukleinsyresammensetning som koder for et humant antistoff som binder human ErbB3, denne sammensetningen omfatter en nukleinsyre som omfatter en sekvens som koder for en tung kjede variabel region, denne sekvensen er minst 90 % identisk med, eller er en sekvens som hybridiserer under høy stringens til SEKV ID NR: 2 og en nukleinsyre som omfatter en sekvens som koder for en lett kjede variabel region, denne sekvensen er minst 90 % identisk med, eller er en sekvens som hybridiser under høy stringens til SEKV ID NR: 26. Nukleinsyrene kan være omfattet i ekspresjonsvektorer. 3 [0023] Foreliggende oppfinnelse tilveiebringer videre vertscelletransgene ikke-humane pattedyr, hybridomer og transgene planter som uttrykker og/eller produserer antistoffene og antigenbindende deler som er beskrevet heri. [0024] Tilveiebrakt ved foreliggende oppfinnelse er også kits som omfatter én eller flere isolerte, monoklonale antistoffer eller antigenbindende deler derav beskrevet heri og instruksjoner for anvendelse ved behandling eller

6 1 diagnostisering av en sykdom assosiert med ErbB3 avhengig signalisering, så som kreft. Antistoffer og antigenbindende deler derav ifølge foreliggende oppfinnelse kan anvendes i en lang rekke terapeutiske og diagnostiske applikasjoner, særlig onkologiske applikasjoner. I et annet aspekt tilveiebringer oppfinnelsen følgelig et antistoff eller antigenbindende del derav, ifølge oppfinnelsen for anvendelse i en fremgangsmåte for inhibering av EGF-lignende ligandmediert fosforylering av ErbB3 eller behandling av kreft i et menneske, eventuelt hvori kreftformen er melanom, brystkreft, eggstokk-kreft, nyrekarsinom, gastrointestinal/colonkreft, lungekreft, klarcellekarsinom eller prostatakreft. Antistoffene eller antigenbindende deler derav kan administreres alene eller i kombinasjon med andre terapeutiske midler, som for eksempel anti-kreft midler, f.eks. andre antistoffer, kjemoterapeutiske midler og/eller stråling. [002] I én utførelse tilveiebringer oppfinnelsen et antistoff eller antigenbindende del derav ifølge oppfinnelsen for anvendelse ifølge oppfinnelsen, hvori antistoffet eller antigenbindende del derav, blir administrert intravenøst, intramuskulært eller subkutant til subjektet, og/eller blir administrert i kombinasjon med et andre anti-kreftantistoff, så som anti-igf1r antistoff, et anti-egfr-antistoff eller et anti-cmet antistoff, eller et anti-kreft small molecule, så som en antimetabolitt, et alkyleringsmiddel, en topoisomeraseinhibitor, et mikrotubuli-rettet middel, en kinase-inhibitor, en proteinsynteseinhibitor, et immunoterapeutikum, et hormon eller analog derav, en somatostatinanalog, et glukokortikoid, en aromataseinhibitor, en mtor-inhibitor, et small molecule målsøkende IGF1R eller et small molecule målsøkende EGFR. 2 [0026] I enda andre utførelsesformer tilveiebringer oppfinnelsen et antistoff eller antigenbindende del ifølge oppfinnelsen for anvendelse i en fremgangsmåte for å diagnostisere en kreft i en forbindelse med ErbB3 i et subjekt, som omfatter (a) å bringe ex vivo eller in vivo celler fra subjektet i kontakt med nevnte antistoff eller antigenbindende del derav, og (b) måle nivået av binding av nevnte antistoff eller antigenbindende del til ErbB3 på cellene, hvori unormalt høye nivåer av binding av nevnte antistoff eller antigenbindende del til ErbB3 indikerer at subjektet har en kreft assosiert med ErbB3. [0027] Andre trekk og fordeler ved oppfinnelsen vil fremgå fra den følgende detaljerte beskrivelsen og fra kravene. Kort beskrivelse av tegningene [0028] 3 Figurene 1A og 1B er søylediagrammer som viser bindingen av ulike anti-erbb3 antistoffkandidater (Fabs, referert til heri som Abs) til ErbB3 uttrykt på MALME-3M melanomceller ved anvendelse av et geite antimenneske Alexa 647 sekundært antistoff Figurene 2A-2D er grafer som viser K D -verdiene av ulike anti-erbb3 antistoffkandidater. Figurene 2A og 2B er grafer som viser K D -verdien av henholdsvis Antistoff #6 (referert til som Ab #6) og Antistoff #3 (også referert til som Ab #3), som målt ved anvendelse av overflate-plasmonresonans (SPR) teknologi. Figur 2C

7 og 2D er grafer som viser K D -verdiene av henholdsvis Ab #6 og Ab #3, som målt ved anvendelse av en celle-bindingsanalyse ved anvendelse av MALME-3M melanomceller. Fig. 3 er en graf som viser bindingsspesifisiteten av et anti-erbb3-antistoff (Ab #6) til ErbB3 ved anvendelse av ELISA. EGFR, BSA og TGF-α ble anvendt som kontroller. Figur 4 er en graf som viser evnen et anti-erbb3-antistoff (Ab #6) har til å redusere totale ErbB3 nivåer i MALME-3M melanomceller in vitro, som målt ved anvendelse av ELISA. Figur A og B er grafer som viser evnen et anti-erbb3-antistoff (Ab #6) har til å nedregulere ErbB3 reseptorer på MALME-3M-celler, målt ved anvendelse av FACS-analyse. Figur A viser resultatene ved anvendelse av en IgG1-isotype av antistoffet. Figur B viser resultatene ved anvendelse av en IgG2-isotype av antistoffet. Figurene 6A-6D er grafer som viser tidsforløpet av antistoff-mediert ErbB3 nedregulering (Ab #6), som målt ved anvendelse av FACS analyse. Figur 7 er et søylediagram som viser evnen forskjellige anti-erbb3-antistoffer har til å nedregulere ErbB3 i melanomceller in vivo. Figur 8 er et søylediagram som viser evnen et anti-erbb3-antistoff (Ab #6) har til å nedregulere ErbB3 i ADRr xenografter in vivo. Figur 9 er en graf som viser evnen et anti-erbb3-antistoff (Ab #6) har til å inhibere proliferasjon av MALME-3M-celler i en celletiter glød-analyse. Figur er en graf som viser evnen et anti-erbb3-antistoff (Ab #6) har til å inhibere celleproliferasjon i en ovarial cellelinje, ADRr. Figur 11 er en graf som viser evnen et anti-erbb3-antistoff (Ab #6) har til å inhibere proliferasjon av ACHNceller. Figur 12 er et søylediagram som viser evnen et anti-erbb3-antistoff (Ab #6) har til å inhibere ErbB3- fosforylering i ADRr xenografter in vivo. Figurene 13A-13C er grafer som viser evnen et anti-erbb3-antistoff (Ab #6) har til å inhibere betacellulin og heregulin-mediert fosforylering av ErbB3 i ADRr-celler. Figurene 14A-14B er grafer som viser evnen et anti-erbb3-antistoff (Ab #6 IgG2-isotypen) har til å inhibere ErbB3-fosforylering i ovariale tumorcellelinjer OVCAR og OVCAR 8. Figurene 1A-1C er grafer som viser evnen betacellulin (BTC) har til å binde ErbB1 som vist ved mangel på binding til ErbB1 negative MALME-3M celler (Figur 17A), binding til ErbB1 positive ADRr-celler ved konsentrasjoner på henholdsvis nm (figur 17B) og 0 nm (figur 17B), og inhiberingen av slik binding ved erbitux. Figurene 16A-16B er grafer som viser evnen et anti-erbb3-antistoff (Ab #6 IgG2-isotypen) har til å inhibere heregulin-mediert signalisering i MALME-3M celler. Figur 16A viser evnen Ab #6 har til å inhibere heregulin-mediert fosforylering av ErbB3 i MALME-3M-celler og 16B viser evnen Ab #6 har til å inhibere fosforylering av AKT i MALME-3M celler.
 har til å nedregulere ErbB3 reseptorer på MALME-3M-celler, målt ved anvendelse av FACS-analyse.")
8 7 1 2 Figurene 17A-D er grafer som viser evnen et anti-erbb3-antistoff (Ab #6) har til å inhibere (A) ovarial (ADRr-celler), (B) prostata Du14 celler), (C) ovarial (OvCAR8 celler) og (D) pankreatisk (Colo37 celler) tumorvekst via xenograftstudier. Figurene 18A og 18B er grafer som viser evnen Ab #6 (Figur 18A) og Fab for Ab #3 (Figur 18B) har til å inhibere heregulinbinding til ErbB3 på MALME-3M celler, som målt ved anvendelse av FACS-analyse. Figurene 19A og 19B er grafer som viser evnen Ab #6 har til å inhibere bindingen av epiregulin til ErbB3 på ADRr-celler. Figur 19A viser bindingen av epiregulin til ADRr-celler, og figur 19B viser evnen både erbitux og Ab #6 har til å inhibere epiregulinbinding til ADRr-celler. Figurene A og B er grafer som viser evnen heparinbindende epidermal vekstfaktor (HB-EGF) har til å binde ErbB på ADRr-celler (Figur A) og den manglende evnen et anti-erbb3-antistoff (Ab #6) har til å inhibere slik binding (figur B). Figurene 21A-21C viser aminosyresekvensene av de variable tung og lett kjede regionene av antistoffer: Ab #6, Ab #3, Ab #14, Ab #17 og Ab #19. Figurene 22A-22B viser nukleotidsekvensene av de variable tung og lett kjede regionene av antistoffer: Ab #6, Ab #3 og Ab #14. Figur 23 viser aminosyresekvensene av de variable lett kjede regionene av antistoffer: Ab #6, Ab #17 og Ab #19, som har blitt revertert til den tilsvarende kimlinje aminosyresekvensen. Aminosyrerestforandringer er understreket. Figurene 24A-24C er grafer som viser evnen Ab #6 har til å inhibere VEGF-sekresjon i tumorceller. Figur 2 er en graf som viser virkningen av Ab #6 på cellemigrering. Figurene 26A-C er grafer som viser (A) inhibering av sfæroidevekst i AdrR-celler, (B) inhibering av HRGindusert sfæroidevekst i AdrR og (C) inhibering av HRG-indusert sfæroidevekst i Du14 celler. Figurene 27 A og B er grafer som viser virkningen av Ab #6 på (A) HRG og (B) BTC-binding til AdrR-celler. Figur 28 er en graf som viser virkningen av Ab #6 på HGF-indusert ErbB3-fosforylering. Figurene 29 A og B viser virkningen av Ab #6 på fosforylering av (A) perbb1 og perbb3 og (B) HRG-indusert ErbB2/3 kompleksdannelse. Figur er en graf som viser at Ab #6 binder aminosyrerester -2 av ErbB3. Detaljert beskrivelse av oppfinnelsen [0029] For at foreliggende oppfinnelse kan bli bedre forstått, blir visse begreper først definert. Ytterligere definisjoner er fremlagt gjennom hele den detaljerte beskrivelsen. 3 I. Definisjoner [00] Begrepene "ErbB3", "HER3," "ErbB3-reseptor" og "HER3-reseptor", som anvendt om hverandre heri, refererer til humant ErbB3 protein, som beskrevet i U.S. Pat. nr.,480,968 og Plowman et al. Proc. Natl. Acad Sci.

9 8 USA, 87: (1990), se, også Kani et al. Biochemistry 44: (0), Cho og Leahy, Science 297: (02)). [0031] Begrepet "EGF-lignende ligand," som anvendt heri, refererer til ligander av epidermal vekstfaktorreseptor (EGFR), inkludert epidermal vekstfaktor (EGF) og nært beslektede proteiner, så som transformerende vekstfaktor-α (TGF-α), betacellulin (BTC), heparin-bindende epidermal vekstfaktor (HB-EGF), biregulin (BIR) og amfiregulin (AR), som binder til EGFR på overflaten av celler og stimulerer reseptorens iboende protein-tyrosin-kinaseaktivitet. Spesifikt induserer EGF-lignende ligander dannelsen av EGFR (også referert til som ErbB1) og ErbB3 proteinkompleks (se f.eks. Kim et al. (1998) Biochem J., 334:189-19), som resulterer i fosforylering av tyrosinrester i komplekset. [0032] Antistoffene og antigenbindende deler derav ifølge foreliggende oppfinnelse inhiberer EGF-lignende 1 ligandmediert fosforylering av ErbB3 og utviser, i visse utførelsesformer, én eller flere av de følgende ytterligere egenskaper: (i) inhibering av én eller flere av heregulin, epiregulin, epigen og biregulin (BIR)-mediert signalisering gjennom ErbB3, (ii) inhibering av proliferasjon av celler som uttrykker ErbB3, (iii) evnen til å redusere nivåer av ErbB3 på celleoverflater, (iv) inhibering av VEGF-sekresjon fra celler som uttrykker ErbB3, (v) inhibering av migreringen av celler som uttrykker ErbB3, (vi) inhibering av sfæroidevekst av celler som uttrykker ErbB3, og/eller (vii) binding til en epitope lokalisert på domene I av ErbB3, f.eks. en epitope som involverer eller spenner over restene - 2 av aminosyresekvensen av ErbB3. [0033] Begrepet "inhibering" som anvendt heri, refererer til en hvilken som helst statistisk signifikant reduksjon i biologisk aktivitet, inkludert fullstendig blokkering av aktiviteten. For eksempel kan "inhibering" referere til en reduksjon på omtrent %, %, %, 40 %, 0 %, 60 %, 70 %, 80 %, 90 % eller 0 % i biologisk aktivitet. 2 [0034] Følgelig refererer begrepet "inhibering av EGF-lignende ligandmediert fosforylering av ErbB3," som anvendt heri, til evnen et antistoff eller antigenbindende del har til statistisk signifikant å redusere fosforyleringen av ErbB3 indusert ved en EGF-lignende ligand, i forhold til fosforyleringen i en ubehandlet (kontroll) celle. Cellen som uttrykker ErbB3 kan være en naturlig forekommende celle eller cellelinje eller kan bli rekombinant produsert ved å introdusere nukleinsyre som koder for ErbB3 inn i en vertscelle. I én utførelsesform inhiberer antistoffet eller den antigenbindende delen derav EGF-lignende ligandmediert fosforylering av ErbB3 med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 %, som bestemt, for eksempel ved Western-blott fulgt av utforsking med et anti-fosfotyrosinantistoff som beskrevet i Kim et al. (1998) Biochem J., 334: og eksemplene infra. 3 [003] Frasen "inhibering av heregulin, epiregulin, epigen eller biregulin-mediert signalisering ved ErbB3," som anvendt heri, refererer til evnen et antistoff eller en antigenbindende del derav har til statistisk signifikant å redusere signalisering mediert ved en ErbB3-ligand (f.eks. heregulin, epiregullin, epigen og biregulin) ved ErbB3, i forhold til signaliseringen i fravær av antistoffet (kontroll). ErbB3-ligander blir også referert til heri som "heregulin-
 og ErbB3 proteinkompleks (se f.eks. Kim et al.")
10 9 lignende ligander." Dette betyr at i nærvær av antistoffet eller antigenbindende del derav, blir et signal mediert i en celle som uttrykker ErbB3 ved én eller flere av heregulin, epiregulin, epigen og biregulin, i forhold til en kontroll (uten antistoff), statistisk signifikant redusert. Et ErbB3-ligandmediert signal kan måles ved analyse av nivået eller aktiviteten av et ErbB3 substrat, og/eller et protein som foreligger i en cellulær kaskade som involverer ErbB3. I én utførelsesform, reduserer antistoffet eller antigenbindende del derav nivået eller aktiviteten av et ErbB3 substrat og/eller det/den av et protein i en cellulær kaskade som involverer ErbB3, med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % i forhold til nivået eller aktiviteten i fravær av slikt antistoff eller antigenbindende del derav (kontroll). Slik ErbB3-ligandmediert signalisering kan måles ved anvendelse av faglig anerkjente teknikker som måler nivået eller aktiviteten av et substrat av ErbB3 (f.eks. SHC eller PI3K) eller et protein i en cellulær kaskade som involverer ErbB3 (f.eks. AKT) ved anvendelse av kinaseanalyser for slike proteiner (se, f.eks. Horst et al. supra, Sudo et al. (00) Methods Enzymol, 322:388-92; og Morgan et al. (1990) Eur. J. Biochem. 191: ). 1 [0036] I en spesiell utførelsesform, inhiberer antistoffet eller antigenbindende del derav ErbB3-ligand (f.eks. heregulin, epiregulin, epigen eller biregulin) mediert signalisering ved ErbB3 ved å inhibere bindingen av ErbB3- liganden (f.eks. én eller flere av heregulin, epiregulin, epigen eller biregulin) til ErbB3. Noen ligander (f.eks. biregulin eller BIR) fungerer både som EGF-lignende ligander (dvs. binder til EGFR/ErbB1) så vel som ErbB3- lignende ligander (dvs. binder til ErbB3). 2 [0037] Frasen "inhibering av heregulin, epiregulin, epigen eller biregulin-binding til ErbB3," som anvendt heri, refererer til evnen et antistoff eller en antigenbindende del derav har til statistisk signifikant å redusere bindingen av en ErbB3-ligand (f.eks. én eller flere av heregulin, epiregulin, epigen eller biregulin) til ErbB3, i forhold til bindingen i fravær av antistoff (kontroll). Dette betyr at, i nærværet av antistoffet eller antigenbindende del derav, blir mengden av ErbB3-liganden (f.eks. heregulin, epiregulin, epigen eller biregulin) som binder til ErbB3 i forhold til en kontroll (uten antistoff), statistisk signifikant redusert. Mengden av en ErbB3-ligand som binder ErbB3 kan bli redusert i nærværet av et antistoff eller antigenbindende del derav ifølge oppfinnelsen med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % i forhold til mengden i fraværet av antistoffet eller antigenbindende del derav (kontroll). En reduksjon i ErbB3-ligandbinding kan måles ved hjelp av faglig anerkjente teknikker som måler nivået av binding av merket ErbB3-ligand (f.eks. radiomerket heregulin, epiregulin, epigen eller biregulin) til celler som uttrykker ErbB3 i nærvær eller fravær (kontroll) av antistoffet eller antigenbindende del derav 3 [0038] Frasen "inhibering av proliferasjon av en celle som uttrykker ErbB3," som anvendt heri, refererer til evnen et antistoff eller en antigenbindende del derav har til statistisk signifikant å redusere proliferasjon av en celle som uttrykker ErbB3 i forhold til proliferasjonen i fraværet av antistoffet. I én utførelsesform kan proliferasjonen av en celle som uttrykker ErbB3 (f.eks. en kreftcelle) reduseres med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % når cellene bringes i kontakt med antistoffet, eller antigenbindende del derav ifølge foreliggende oppfinnelse, i forhold

11 til proliferasjonen målt i fraværet av antistoffet eller antigenbindende del derav (kontroll). Cellulær proliferasjon kan bli analysert ved anvendelse av faglig anerkjente teknikker som måler raten av celledeling, fraksjonen av celler innen en cellepopulasjon som gjennomgår celledeling, og/eller rate av celletap fra en cellepopulasjon på grunn av terminal differensiering eller celledød (f.eks. ved anvendelse av en celletiter glød analyse eller tymidininkorporering). [0039] Frasen "evnen til å redusere nivået av ErbB3 på celleoverflaten," som anvendt heri, refererer til evnen et antistoff eller antigenbindende del derav har til statistisk signifikant å redusere mengden ErbB3 funnet på 1 2 overflaten av en celle som er blitt eksponert for antistoffet i forhold til en ubehandlet (kontroll) celle. For eksempel kan en reduksjon i nivåer av ErbB3 på celleoverflater resultere fra øket internalisering av ErbB3 (eller øket ErbB3 endocytose). I én utførelsesform, avtar antistoffet eller antigenbindende del derav celleoverflateekspresjonen av ErbB3 med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % og/eller øker internalisering av ErbB3-reseptoren med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % i forhold til celleoverflateekspresjonen eller internaliseringen i fraværet av antistoffet eller antigenbindende del derav (kontroll). Nivåene av ErbB3 på overflater av celler og/eller internalisering av ErbB3-reseptoren i fraværet og nærværet av et antistoff eller antigenbindende del derav, kan lett måles ved anvendelse av faglig anerkjente teknikker, så som de beskrevet i Horst et al. supra og i eksemplene heri. [0040] Frasen "inhibering av VEGF-sekresjon fra celler som uttrykker ErbB3," som anvendt heri, refererer til evnen et antistoff eller en antigenbindende del derav har til statistisk signifikant å redusere VEGF-sekresjon fra en celle som uttrykker ErbB3 i forhold til VEGF-sekresjon i fraværet av antistoffet. I én utførelsesform kan VEGF sekresjonen av en celle som uttrykker ErbB3 (f.eks. en kreftcelle) bli redusert med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % når cellene bringes i kontakt med antistoffet eller antigenbindende del derav ifølge foreliggende oppfinnelse, i forhold til VEGF-sekresjonen målt i fraværet av antistoffet eller antigenbindende del derav (kontroll). VEGF-sekresjon kan bli analysert ved anvendelse av faglig anerkjente teknikker, så som de beskrevet heri. 3 [0041] Frasen "inhibering av migreringen av celler som uttrykker ErbB3," som anvendt heri, refererer til evnen et antistoff eller en antigenbindende del derav har til statistisk signifikant å redusere migreringen av en celle som uttrykker ErbB3 i forhold til migreringen av cellen i fraværet av antistoffet. I én utførelsesform kan migreringen av en celle som uttrykker ErbB3 (f.eks. en kreftcelle) reduseres med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % når cellene bringes i kontakt med antistoffet eller antigenbindende del derav ifølge foreliggende oppfinnelse, i forhold til cellemigrering målt i fraværet av antistoffet eller antigenbindende del derav (kontroll). Cellemigrering kan analyseres ved anvendelse av faglig anerkjente teknikker, så som de beskrevet heri.

12 11 1 [0042] Frasen "inhibering av sfæroidevekst av celler som uttrykker ErbB3," som anvendt heri, refererer til evnen et antistoff eller en antigenbindende del derav har til statistisk signifikant å redusere migreringen av en celle som uttrykker ErbB3 i forhold til migreringen av cellen i fraværet av antistoffet. I én utførelsesform kan migreringen av en celle som uttrykker ErbB3 (f.eks. en kreftcelle) reduseres med minst %, eller minst %, eller minst %, eller minst 40 %, eller minst 0 %, eller minst 60 %, eller minst 70 %, eller minst 80 %, eller minst 90 %, eller 0 % når cellene bringes i kontakt med antistoffet eller antigenbindende del derav ifølge foreliggende oppfinnelse, i forhold til cellemigrering målt i fraværet av antistoffet eller antigenbindende del derav (kontroll). Cellemigrering kan analyseres ved anvendelse av faglig anerkjente teknikker, så som de beskrevet heri. Begrepet "antistoff" eller "immunoglobulin", som anvendt om hverandre heri, inkluderer hele antistoffer og ethvert antigenbindende fragment (dvs. "antigenbindende del") eller enkle kjeder derav. Et "antistoff" omfatter minst to tunge (H) kjeder og to lette (L) kjeder innbyrdes forbundet med disulfid-bindinger. Hver tunge kjede er omfattet av en tung kjede variabel region (forkortet heri som V H ) og en tung kjede konstant region. Tung kjede konstant regionen er omfattet av tre domener, CH1, CH2 og CH3. Hver lette kjede er omfattet av en lett kjede variabel region (forkortet heri som V L ) og en lett kjede konstant region. Lett kjede konstant regionen er omfattet av ett domene, CL. V H og V L - regionene kan videre deles inn i regioner av hypervariabilitet, betegnet komplementaritetsbestemmende regioner (CDR), ispedd regioner som er mer bevart, betegnet rammeverkregioner (FR). Hver V H og V L er sammensatt av tre CDRer og fire FRer, arrangert fra aminoterminus til karboksyterminus i den følgende rekkefølgen: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. De variable regionene i de tunge og lette kjedene inneholder et bindende domene som vekselvirker med et antigen. De konstante regionene i antistoffene kan mediere bindingen av immunoglobulinet til vertsvev eller faktorer, inkludert ulike celler av immunsystemet (f.eks. effektorceller) og den første komponenten (Clq) av det klassiske komplementsystemet. Eksempelvise antistoffer ifølge oppfinnelsen inkluderer antistoffer #1, 3 og 14, og antigenbindende deler derav. 2 3 [0043] Begrepet "antigenbindende del" av et antistoff (eller ganske enkelt "antistoffdel"), som anvendt heri, refererer til ett eller flere fragmenter av et antistoff som beholder evnen til å spesifikt binde til et antigen (f.eks. ErbB3). Det har blitt vist at den antigenbindende funksjonen for et antistoff kan bli utført ved fragmenter av et fulllengde antistoff. Eksempler på bindende fragmenter omfavnet innen begrepet "antigenbindende del" av et antistoff inkluderer (i) et Fab-fragment, et monovalent fragment bestående av V L, V H, CL og CH1 domenene, (ii) et F(ab') 2 -fragment, et bivalent fragment omfattende to Fab-fragmenter knyttet sammen ved en disulfidbro ved hengselregionen, (iii) et Fd-fragment bestående av V H og CH1 domenene, (iv) et Fv fragment bestående av V L og V H -domenene av en enkelt arm av et antistoff, (v) et dab inkludert VH- og VL-domener, (vi) et dab fragment (Ward et al. (1989) Nature 341, 44-46), som består av et V H -domene, (vii) et dab som består av et VH eller et VLdomene, og (viii) en isolert komplementaritetsbestemmende region (CDR) eller (ix) en kombinasjon av to eller flere isolerte CDRer som eventuelt kan være sammenføyd ved en syntetisk linker. Videre, selv om de to domenene av Fv-fragmentet, V L og V H, er kodet for ved separate gener, kan de bli sammenføyd, ved anvendelse av rekombinante metoder, ved en syntetisk linker som gjør dem i stand til å bli dannet som en enkelt proteinkjede hvori V L og V H - regionene pares for å danne monovalente molekyler (kjent som enkeltkjede Fv (scfv), se f.eks. Bird et al. (1988) Science 242, , og Huston et al. (1988) Proc. Natl. Acad Sci. 8 USA, ). Slike enkeltkjede antistoffer

13 12 1 er også ment å være omfavnet innen begrepet "antigenbindende del" av et antistoff. Disse antistoff-fragmentene oppnås ved anvendelse av konvensjonelle teknikker kjent for fagpersonene, og fragmentene blir screenet for nytte på samme måte som intakte antistoffer blir det. Antigenbindende deler kan produseres ved rekombinante DNAteknikker, eller ved enzymatisk eller kjemisk spalting av intakte immunoglobuliner. [0044] Begrepet "monoklonalt antistoff" som anvendt heri refererer til et antistoff oppnådd fra en populasjon av hovedsakelig homogene antistoffer, dvs. de individuelle antistoffene som omfatter populasjonen er identiske med unntak av mulige naturlig forekommende mutasjoner som kan foreligge i mindre mengder. Monoklonale antistoffer er svært spesifikke, ved å være rettet mot et enkelt antigent sete. Videre, i motsetning til konvensjonelle (polyklonale) antistoffpreparater som typisk inkluderer forskjellige antistoffer rettet mot forskjellige determinanter (epitoper), blir hvert monoklonale antistoff rettet mot en enkelt determinant på antigenet. Monoklonale antistoffer kan fremstilles ved anvendelse av hvilken som helst faglig anerkjent teknikk og de beskrevet heri, så som for eksempel en hybridoma-metode, som beskrevet ved Kohler et al. (197) Nature, 26:49, et transgent dyr, som beskrevet ved, for eksempel, (se f.eks. Lonberg, et al. (1994) Nature 368 (6474): 86-89), rekombinante DNA-metoder (se, f.eks. U.S. Pat. nr. 4,816,67), eller ved anvendelse av fag-antistoffbiblioteker ved anvendelse av teknikkene beskrevet i f.eks. Clackson et al. Nature, 32: (1991) og Marks et al. J. Mol. Biol. 222:81-97 (1991). Monoklonale antistoffer inkluderer kimære antistoffer, humane antistoffer og humaniserte antistoffer, og kan forekomme naturlig eller kan produseres rekombinant. 2 [004] Begrepet "rekombinant antistoff" refererer til antistoffer som blir fremstilt, uttrykt, skapt eller isolert ved rekombinante metoder, så som (a) antistoffer isolert fra et dyr (f.eks. en mus) som er transgen eller transkromosomal for immunoglobulingener (f.eks. humane immunoglobulingener) eller et hybridom fremstilt derfra, (b) antistoffer isolert fra en vertscelle transformert for å uttrykke antistoffet, f.eks. fra et transfektom, (c) antistoffer isolert fra et rekombinant, kombinatorisk antistoffbibliotek (f.eks. inneholdende humane antistoffsekvenser) ved anvendelse av fag-display, og (d) antistoffer fremstilt, uttrykt, skapt eller isolert ved en hvilken som helst annen metode som involverer spleising av immunglobulingensekvenser (f.eks. humane immunoglobulingener) til andre DNA-sekvenser. Slike rekombinante antistoffer kan ha variable og konstante regioner avledet fra humane kimlinje immunoglobulinsekvenser. I visse utførelsesformer, kan imidlertid slike rekombinante humane antistoffer underkastes in vitro mutagenese og således er aminosyresekvensene av V H og V L -regionene av de rekombinante antistoffene sekvenser som, selv om de er avledet fra og relatert til humane kimlinje V H og V L -sekvenser, ikke nødvendigvis kan eksistere naturlig innen det humane antistoff kimlinjerepertoaret in vivo. 3 [0046] Begrepet "kimært immunoglobulin" eller antistoff refererer til et immunoglobulin eller antistoff som har variable regioner avledet fra en første art og som har konstante regioner avledet fra en andre art. Kimære immunglobuliner eller antistoffer kan konstrueres, for eksempel ved genteknologi, fra immunglobulingensegmenter som tilhører ulike arter.

14 13 [0047] Begrepet "humant antistoff", som anvendt heri, er ment å inkludere antistoffer som har variable regioner hvori både rammeverk og CDR-regionene er avledet fra humane kimlinje immunoglobulinsekvenser som beskrevet, for eksempel ved Kabat et al. (se Kabat et al. (1991) Sequences of proteins of Immunological Interest, femte utgave, US Department of Health and Human Services, NIH publikasjon nr ). Videre, hvis antistoffet inneholder en konstant region, er den konstante regionen også avledet fra humane kimlinje immunoglobulinsekvenser. De humane antistoffene kan inkludere aminosyrerester som ikke kodes for ved humane kimlinje immunoglobulinsekvenser (f.eks. mutasjoner introdusert ved tilfeldig eller setespesifikk mutagenese in vitro eller ved somatisk mutasjon in vivo). Imidlertid er begrepet "humant antistoff", som anvendt her, ikke tenkt å inkludere antistoffer hvori CDR-sekvenser avledet fra kimlinjen av en annen pattedyrart, så som en mus, har blitt podet på humane rammeverk-sekvenser. [0048] Det humane antistoffet kan ha minst én eller flere aminosyrer erstattet med en aminosyrerest, f.eks. en 1 aktivitetsstyrkende aminosyrerest som ikke kodes for ved den humane kimlinje immunoglobulinsekvensen. Typisk kan det humane antistoffet ha opptil tyve posisjoner erstattet med aminosyrerester som ikke er en del av den humane kimlinje immunoglobulinsekvensen. I en spesiell utførelsesform er disse erstatningene innen CDRregionene som beskrevet i detalj nedenfor. 2 [0049] Begrepet "humanisert immunoglobulin" eller "humanisert antistoff" refererer til et immunoglobulin eller antistoff som inkluderer minst ett humanisert immunoglobulin eller antistoffkjede (dvs. minst én humanisert lett eller tung kjede). Begrepet "humanisert immunoglobulinkjede" eller "humanisert antistoffkjede" (dvs. en "humanisert immunglobulin lett kjede" eller "humanisert immunglobulin tung kjede") refererer til en immunoglobulin- eller antistoffkjede (dvs. henholdsvis en lett eller tung kjede) som har en variabel region som inkluderer en variabel rammeverkregion hovedsakelig fra et humant immunoglobulin eller antistoff og komplementaritetsbestemmende regioner (CDRer) (f.eks. minst én CDR, fortrinnsvis to CDRer, mer foretrukket tre CDRer), hovedsaken fra et ikke-humant immunoglobulin eller antistoff, og inkluderer videre konstante regioner (f.eks. minst én konstant region eller del derav, i tilfellet av en lett kjede, og fortrinnsvis tre konstante regioner i tilfellet av en tung kjede). Begrepet "humanisert variabel region" (f.eks. "humanisert lett kjede variabel region" eller "humanisert tung kjede variabel region") refererer til en variabel region som inkluderer en variabel rammeverkregion hovedsakelig fra et humant immunoglobulin eller antistoff og komplementaritetsbestemmende regioner (CDRer), hovedsakelig fra et ikke-humant immunglobulin eller antistoff. [000] Et "bispesifikt" eller "bifunksjonelt antistoff" er et kunstig hybrid antistoff som har to forskjellige tung/lett 3 kjede par og to forskjellige bindingsseter. Bispesifikke antistoffer kan produseres ved en rekke metoder inkludert fusjon av hybridomer eller tilknytning av Fab'-fragmenter. Se, f.eks. Songsivilai & Lachmann, (1990) Clin. Exp. Immunol. 79, ; Kostelny et al. (1992) J. Immunol. 148, I en spesiell utførelsesform inkluderer et bispesifikt antistoff i henhold til foreliggende oppfinnelse bindingsseter for både ErbB3 og IGF1-R (dvs. insulinlignende vekstfaktor-1-reseptor). I en annen utførelsesform inkluderer et bispesifikt antistoff i henhold til foreliggende oppfinnelse bindingsseter for både ErbB3-og C-MET. I andre utførelsesformer inkluderer et bispesifikt

15 14 antistoff et bindingssete for ErbB3 og et bindingssete for ErbB2, ERbB3, ErbB4, EGFR, Lewis Y, MUC-1, EpCAM, CA12, prostataspesifikt membranantigen, PDGFR-α, PDGFR-β, C-KIT eller en hvilken som helst av FGFreseptorene. [001] Som anvendt heri, er et "heterologt antistoff" definert i forhold til den transgene ikke-humane organismen eller planten som produserer et slikt antistoff. [002] Et "isolert antistoff", som anvendt heri, er ment å referere til et antistoff som er hovedsakelig uten andre antistoffer som har forskjellige antigene spesifisiteter (f.eks. er et isolert antistoff som spesifikt binder til ErbB3 i det vesentlige uten antistoffer som spesifikt binder antigener andre enn ErbB3). I tillegg er et isolert antistoff typisk hovedsakelig uten annet cellulært materiale og/eller kjemikalier. I én utførelsesform av oppfinnelsen blir en kombinasjon av "isolerte" monoklonale antistoffer som har forskjellige ErbB3 bindingsspesifisiteter kombinert i en veldefinert sammensetning. 1 [003] Som anvendt heri refererer "isotype" til antistoffklassen (f.eks. IgM eller IgG1) som blir kodet for ved tung kjede konstant region gener. I én utførelsesform er et antistoff eller antigenbindende del derav av en isotype valgt fra en IgG1, en IgG2, en IgG3, en IgG4, en IgM, en IgA1 en IgA2, en IgAsec, en IgD eller en IgE-antistoff-isotype. I noen utførelsesformer er et monoklonalt antistoff ifølge foreliggende oppfinnelse av IgG1 isotypen. I andre utførelsesformer er et monoklonalt antistoff ifølge oppfinnelsen av IgG2-isotypen. [004] Som anvendt heri refererer "isotypeskifte" til fenomenet ved hvilket klassen eller isotypen, av et antistoff forandres fra én Ig-klasse til én av de andre Ig-klassene. 2 [00] Som anvendt heri refererer "ikke-skiftet isotype" til den isotypiske klassen av tung kjede som produseres når intet isotypeskifte har funnet sted; CH-genet som koder for den ikke-skiftede isotypen er typisk det første CHgenet umiddelbart nedstrøms for det funksjonelt omarrangerte VDJ-genet. Isotypeskifte har blitt klassifisert som klassisk eller ikke-klassisk isotypeskifte. Klassisk isotypeskifte forekommer ved rekombineringshendelser som involverer minst én skiftesekvensregion i et gen som koder for et antistoff. Ikke-klassisk isotypeskifte kan forekomme ved for eksempel homolog rekombinering mellom human σ μ og human Σ μ ( -assosiert delesjon). Alternative ikke-klassiske skiftemekanismer, så som intertransgen og/eller interkromosomal rekombinering, blant andre, kan forekomme og effektuere isotypeskifte. [006] Som anvendt heri refererer begrepet "skiftesekvens" til de DNA-sekvensene som er ansvarlige for 3 skifterekombinering. En "skiftedonor" sekvens, typisk en μ skifteregion, vil være ' (dvs. oppstrøms) for konstruktregionen som skal slettes i løpet av skifterekombineringen. "Skifteakseptor" regionen vil være mellom konstruktregionen som skal slettes og den erstattende konstante region (f.eks. γ, ε, osv.). Siden det ikke er noe spesifikt sted hvor rekombinering alltid forekommer, vil den endelige gensekvensen typisk ikke være forutsigbar fra konstrukten.
![[002] Et "isolert antistoff", som anvendt heri, er ment å referere til et antistoff som er hovedsakelig uten andre antistoffer som har forskjellige antigene spesifisiteter (f.eks.](/docs-images/46/1534177/images/page_15.jpg "er et isolert antistoff som spesifikt binder til ErbB3 i det vesentlige uten antistoffer som spesifikt binder antigener andre enn ErbB3).")
16 1 [007] Et "antigen" er en enhet (f.eks. en proteinholdig enhet eller peptid) som et antistoff eller antigenbindende del derav binder til. I ulike utførelsesformer av foreliggende oppfinnelse, er et antigen et ErbB3 eller et ErbB lignende molekyl. I en spesiell utførelsesform ifølge oppfinnelsen, er et antigen human ErbB3. [008] Begrepet "epitope" eller "antigendeterminant" refererer til et sete på et antigen som et immunoglobulin eller antistoff spesifikt binder til. Epitoper kan bli dannet både fra tilgrensende aminosyrer eller ikke-tilgrensende aminosyrer sidestilt ved tertiær folding av et protein. Epitoper dannet fra tilgrensende aminosyrer blir typisk beholdt ved eksponering for denaturerende løsemidler, mens epitoper dannet ved tertiær folding vanligvis blir tapt ved behandling med denaturerende løsemidler. En epitope inkluderer typisk minst 3, 4,, 6, 7, 8, 9,, 11, 12, 13, 14 eller 1 aminosyrer i en unik romlig konformasjon. Fremgangsmåter for å bestemme romlig konformasjon av epitoper inkluderer teknikker innen faget og de beskrevet heri, for eksempel, røntgenkrystallografi og 2- dimensjonal kjernemagnetisk resonans. Se, f.eks. Epitope Mapping Protocols in Methods in Molecular Biology, bind 66, GE Morris, Ed. (1996). [009] Også omfavnet ved foreliggende oppfinnelse er antistoffer som binder den samme eller en overlappende epitope som antistoffene ifølge foreliggende oppfinnelse, dvs. antistoffer som konkurrerer om binding til ErbB3, eller binder epitoper som overlapper med epitoper bundet ved antistoffene beskrevet heri, dvs. en epitope lokalisert på domene I av ErbB3. Antistoffer som gjenkjenner den samme epitopen kan bli identifisert ved anvendelse av rutineteknikker så som en immunoanalyse, for eksempel, ved å vise evnen ett antistoff har til å blokkere bindingen av et annet antistoff til et målantigen, dvs. en konkurrerende bindingsanalyse. Konkurrerende binding bestemmes i en analyse hvori immunoglobulin under test inhiberer spesifikk binding av et referanseantistoff til et felles antigen, så som ErbB3. Tallrike typer konkurrerende bindingsanalyser er kjent, for eksempel: fastfase direkte eller indirekte radioimmunoanalyse (RIA), fastfase direkte eller indirekte enzymimmunoanalyse (EIA), sandwich-konkurranse analyse (se Stahli et al. (1983) Methods in Enzymology 9:242), fastfase direkte biotin-avidin EIA (se Kirkland et al. (1986) J. Immunol. 137:3614); fastfase direkte merket analyse, fastfase direkte merket sandwichanalyse (se Harlow og Lane (1988) Antibodies: A Laboratory Manual, Cold Spring Harbor Press); fastfase direkte merke RIA ved anvendelse av I-12 merke (se Morel et al. (1988) Mol. Immunol. 2 (1): 7), fastfase direkte biotin-avidin EIA (Cheung et al. (1990) Virology 176:46); og direkte merket RIA. (Moldenhauer et al. (1990) Scand. J. Immunol. 32:77). Typisk involverer en slik analyse anvendelsen av renset antigen (f.eks. ErbB3) bundet til en fast overflate eller celler som bærer den ene eller andre av disse, et umerket test-immunoglobulin og et merket referanse-immunglobulin. Konkurrerende inhibering måles ved å bestemme mengden av merke bundet til den faste overflaten eller celler i nærværet av test-immunglobulin. Vanligvis er testimmunglobulinet til stede i overskudd. Når et konkurrerende antistoff er til stede i overskudd, vil det vanligvis inhibere spesifikk binding av et referanse-antistoff til et felles antigen ved minst 0- %, -60 %, 60-6 %, 6-70 % 70-7 % eller mer.
![[008] Begrepet "epitope" eller "antigendeterminant" refererer til et sete på et antigen som et immunoglobulin eller antistoff spesifikt binder til.](/docs-images/46/1534177/images/page_16.jpg "Epitoper kan bli dannet både fra tilgrensende aminosyrer eller ikke-tilgrensende aminosyrer sidestilt ved tertiær folding av et protein.")
17 16 [0060] Som anvendt heri, betyr begrepene "spesifikk binding", "binder spesifikt", "selektiv binding" og "binder selektivt," at et antistoff eller antigenbindende del derav, utviser merkbar affinitet for et spesielt antigen eller epitope og, generelt ikke utviser signifikant kryssreaktivitet med andre antigener og epitoper. "Merkbar" eller foretrukket binding inkluderer binding med en affinitet på minst 6, 7, 8, 9 M -1 eller M -1. Affiniteter større enn 7 M -1, foretrukket større enn 8 M -1 er mer foretrukket. Verdier som ligger mellom de som er angitt heri er også ment å være innen omfanget av den foreliggende oppfinnelse og en foretrukket bindingsaffinitet kan bli indikert som et område av affiniteter, for eksempel, 6 til M -1, fortrinnsvis 7 til M -1, mer foretrukket 8 til M -1. Et antistoff som "ikke utviser signifikant kryssreaktivitet" er ett som ikke i merkbar grad binder til en uønsket enhet (f.eks. en uønsket proteinaktig enhet). For eksempel, i en utførelsesform, vil et antistoff eller antigenbindende del derav som spesifikt binder til ErbB3 merkbart binde det ErbB3 molekylet, men vil ikke signifikant reagere med andre ErbB-molekyler og ikke-erbb proteiner eller peptider. Spesifikk eller selektiv binding kan bestemmes i henhold til hvilken som helst faglig anerkjent metode for å bestemme slik binding, inkludert, for eksempel, i henhold til Scatchard-analyse og/eller konkurrerende bindingsanalyser. 1 2 [0061] Begrepet "K D," som anvendt heri, er ment å referere til dissosiasjons-likevektkonstanten for en spesiell antistoff-antigen-vekselvirkning eller affiniteten av et antistoff for et antigen. I én utførelsesform binder antistoffet eller antigenbindende del derav ifølge foreliggende oppfinnelse et antigen (f.eks. ErbB3) med en affinitet (K D ) på 0 nm eller bedre (dvs. eller mindre) (f.eks. 40 nm og nm eller nm eller nm eller mindre), som målt ved anvendelse av en overflate-plasmonresonansanalyse eller en cellebindingsanalyse. I en spesiell utførelsesform, binder et antistoff eller antigenbindende del derav ifølge foreliggende oppfinnelse ErbB3 med en affinitet (K D ) på 8 nm eller bedre (f.eks. 7 nm, 6 nm, nm, 4, nm, 2 nm, 1, nm, 1,4 nm, 1,3 nm, 1 nm eller mindre), som målt ved overflate-plasmonresonansanalyse eller en cellebindingsanalyse. I andre utførelsesformer binder et antistoff eller antigenbindende del derav et antigen (f.eks. ErbB3) med en affinitet (K D ) på omtrent mindre enn -7 M, så som omtrent mindre enn -8 M, -9 M eller - M eller enda lavere når bestemt ved overflate-plasmonresonans (SPR)-teknologi i et Biacore 00 instrument ved anvendelse av rekombinant ErbB3 som analytten og antistoffet som liganden, og binder til det forhåndsbestemte antigenet med en affinitet som er minst to ganger større enn dets affinitet for binding til et ikke-spesifikt antigen (f.eks. BSA, kasein) annet enn det forhåndsbestemte antigenet eller et nært beslektet antigen. [0062] Begrepet "K off," som anvendt heri, er ment å referere til "off-rate" konstanten for dissosiasjonen av et antistoff fra antistoff/antigen-komplekset. 3 [0063] Begrepet "EC0", som anvendt heri, refererer til konsentrasjonen av et antistoff eller en antigenbindende del derav, som induserer en respons, enten i en in vitro eller en in vivo-analyse, som er 0 % av den maksimale responsen, dvs. halvveis mellom den maksimale responsen og grunnlinjen. [0064] Som anvendt heri er "glykosyleringsmønster" definert som mønsteret av karbohydrat-enheter som er kovalent knyttet til et protein, mer spesifikt til et immunglobulinprotein.

18 17 [006] Begrepet "naturlig forekommende" som brukt heri som anvendt på et objekt refererer til det faktum at et objekt kan bli funnet i naturen. For eksempel et polypeptid eller polynukleotid-sekvens som er til stede i en organisme (inkludert virus) som kan isoleres fra en kilde i naturen, og som ikke har blitt modifisert med hensikt av menneske i laboratoriet naturlig forekommende. [0066] Begrepet "omarrangert" som anvendt heri refererer til en konfigurasjon av en tung kjede eller lett kjede immunglobulin-locus, hvori et V-segment er posisjonert umiddelbart tilstøtende til et D-J- eller J-segment i en konformasjon som koder for grunnleggende henholdsvis et fullstendig V H eller V L domene. En omarrangert immunoglobulin genlocus kan identifiseres ved sammenligning med kimlinje DNA; en omarrangert locus vil ha minst ett rekombinert heptamer/nonamer homologielement. 1 [0067] Begrepet "u-omarrangert" eller "kimlinje-konfigurasjon" som anvendt heri med referanse til et V-segment refererer til konfigurasjonen hvori V-segmentet ikke er rekombinert for å være umiddelbart tilstøtende et D- eller J-segment. [0068] Begrepet "nukleinsyremolekyl", som anvendt heri, er ment å inkludere DNA-molekyler og RNA-molekyler. Et nukleinsyremolekyl kan være enkelttrådet eller dobbelttrådet, men er fortrinnsvis dobbelttrådet DNA. 2 [0069] Begrepet "isolert nukleinsyremolekyl", som anvendt heri, med referanse til nukleinsyrer som koder for antistoffer eller antistoffdeler (f.eks. V H, V L, CDR3) som binder til ErbB3, er ment å referere til et nukleinsyremolekyl hvori nukleotidsekvensene som koder for antistoffet eller antistoffdelen er uten andre nukleotidsekvenser som koder for antistoffer som binder andre antigener enn ErbB3, disse andre sekvensene kan naturlig flankere nukleinsyren i humant genomisk DNA. [0070] Begrepet "modifisering" eller "modifikasjon", som anvendt heri, er ment å referere til endring av én eller flere aminosyrer i antistoffene eller antigenbindende deler derav. Endringen kan produseres ved å tilsette, substituere eller slette en aminosyre ved én eller flere posisjoner. Endringen kan produseres ved anvendelse av kjente teknikker, så som PCR-mutagenese. For eksempel kan i noen utførelsesformer, et antistoff eller en antigenbindende del derav identifisert ved anvendelse av fremgangsmåtene ifølge oppfinnelsen bli modifisert, for derved å modifisere bindingsaffiniteten av antistoffet eller antigenbindende del derav til ErbB3. 3 [0071] Foreliggende oppfinnelse omfavner også "konservative aminosyresubstitusjoner" i sekvensene av antistoffene ifølge oppfinnelsen, dvs. nukleotid- og aminosyresekvensmodifikasjoner som ikke opphever bindingen av antistoffet kodet for ved nukleotidsekvensen eller som inneholder aminosyresekvensen, til antigenet, dvs. ErbB3. Konservative aminosyresubstitusjoner inkluderer substitusjonen av en aminosyre i én klasse ved en aminosyre av den samme klassen, hvor en klasse er definert ved felles fysisk-kjemiske aminosyre-sidekjedeegenskaper og høye substitusjonsfrekvenser i homologe proteiner funnet i naturen, som bestemt, for eksempel

19 18 ved en standard Dayhoff frekvensutvekslingsmatrise eller BLOSUM matrise. Seks generelle klasser av aminosyresidekjeder er blitt kategorisert og inkluderer: Klasse I (Cys); Klasse II (Ser, Thr, Pro, Ala, Gly); Klasse III (Asn, Asp, Gin, Glu); klasse IV (His, Arg, Lys), Klasse V (Ile, Leu, Val, Met) og klasse VI (Phe, Tyr, Trp). For eksempel er en substitusjon av Asp for en annen klasse III rest så som Asn, Gln eller Glu, en konservativ substitusjon. Således blir en forutsagt ikke-essensiell aminosyrerest i et anti-erbb3-antistoff fortrinnsvis erstattet med en annen aminosyrerest fra den samme klassen. Fremgangsmåter for å identifisere nukleotid- og aminosyre-konservative substitusjoner som ikke eliminerer antigenbinding er velkjent innen faget (se, f.eks. Brummell et al. Biochem. 32: (1993); Kobayashi et al. Protein Eng. 12 (): (1999), og Burks et al. Proc. Natl. Acad Sci. USA 94: 412 til 417 (1997)). [0072] Begrepet "ikke-konservativ aminosyresubstitusjon" refererer til substitusjonen av en aminosyre i én klasse med en aminosyre fra en annen klasse, for eksempel substitusjon av en Ala, en klasse II-rest, med en klasse III-rest så som Asp, Asn, Glu eller Gln. 1 [0073] Alternativt, i en annen utførelsesform, kan mutasjoner (konservative eller ikke-konservative) bli introdusert tilfeldig langs hele eller del av en anti-erbb3-antistoff-kodende sekvens, så som ved metningsmutagenese, og de resulterende modifiserte anti-erbb3-antistoffene kan screenes for bindingsaktivitet. 2 [0074] En "konsensussekvens" er en sekvens dannet fra de hyppigst forekommende aminosyrene (eller nukleotider) i en familie av beslektede sekvenser (se f.eks. Winnaker, From Genes to Clones (Verlagsgesellschaft, Weinheim, Tyskland 1987). I en familie av proteiner, er hver posisjon i konsensussekvensen opptatt av aminosyren som forekommer hyppigst i den posisjonen i familien. Hvis to aminosyrer forekommer like ofte, kan den ene eller andre bli inkludert i konsensussekvensen. Et "konsensusrammeverk" av et immunglobulin refererer til en rammeverkregion i konsensus immunoglobulinsekvensen. [007] Likeledes kan konsensussekvensen for CDRene bli utledet ved optimal tilpasning av CDR aminosyresekvensene av ErbB3 antistoffer ifølge foreliggende oppfinnelse. [0076] For nukleinsyrer indikerer begrepet "betydelig homologi" at to nukleinsyrer, eller utpekte sekvenser derav, når optimalt innrettet og sammenlignet, er identiske, med egnede nukleotidinsersjoner eller delesjoner, i minst omkring 80 % av nukleotidene, vanligvis minst omkring 90 % til 9 %, og mer foretrukket minst omkring 98 % til 99, % av nukleotidene. Alternativt eksisterer betydelig homologi når segmentene vil hybridisere under selektive hybridiseringsbetingelser, til komplementet av tråden. 3 [0077] Den prosentvise identiteten mellom to sekvenser er en funksjon av antallet identiske posisjoner delt av sekvensene (dvs. % homologi = # identiske posisjoner/totalt # posisjoner x 0), ved å ta hensyn til antallet gap, og lengden av hvert gap, som trenger å bli innført for optimal innretting av de to sekvenser. Sammenligningen av

20 19 sekvenser og bestemmelse av prosentvis identitet mellom to sekvenser kan bli gjennomført ved anvendelse av en matematisk algoritme, så som beskrevet i de ikke-begrensende eksemplene under. [0078] Den prosentvise identitet mellom to nukleotidsekvenser kan bestemmes ved anvendelse av GAPprogrammet i GCG programvaren, ved anvendelse av en NWSgapdna. CMP matrise og en gapvekt på 40, 0, 60, 70 eller 80 og en lengdevekt på 1, 2, 3, 4, eller 6. Den prosentvise identiteten mellom to nukleotid- eller aminosyresekvenser kan også bli bestemt ved anvendelse av algoritmen ifølge E. Meyers og W. Miller (CABIOS, 4:11-17 (1989)) som har blitt inkorporert i ALIGN-programmet (versjon 2.0), ved anvendelse av en PAM1 vektresttabell, en gaplengdestraff på 12 og en gapstraff på 4. I tillegg kan den prosentvise identiteten mellom to aminosyresekvenser bestemmes ved anvendelse av Needleman og Wunsch (J. Mol. Biol. (48): (1970)) algoritmen som har blitt inkorporert i GAP-programmet i GCG programvarepakken, ved anvendelse av enten en Blossum 62 matrise eller en PAM matrise, og en gapvekt på 16, 14, 12,, 8, 6 eller 4 og en lengdevekt på 1, 2, 3, 4, eller [0079] Nukleinsyre- og proteinsekvensene ifølge foreliggende oppfinnelse kan videre anvendes som en "spørresekvens" for å utføre et søk mot offentlige databaser for å, for eksempel, identifisere beslektede sekvenser. Slike søk kan utføres ved anvendelse av NBLAST og XBLAST programmene (versjon 2.0) av Altschul et al. (1990) J. Mol. Biol. 21:403-. BLAST nukleotidsøk kan utføres med NBLAST-programmet, poeng = 0, ordlengde = 12 for å oppnå nukleotidsekvenser homologe med nukleinsyremolekylene ifølge oppfinnelsen. BLAST proteinsøk kan utføres med XBLAST-programmet, poeng = 0, ordlengde = 3 for å oppnå aminosyresekvenser homologe med proteinmolekylene ifølge oppfinnelsen. For å oppnå innrettinger med gap for sammenligningsformål, kan Gapped BLAST benyttes som beskrevet i Altschul et al. (1997) Nucleic Acids Res. 2 (17): Når en benytter BLAST og Gapped BLAST programmer, kan standardparametere for de respektive programmene (f.eks. XBLAST og NBLAST) anvendes. [0080] Nukleinsyrene kan foreligge i hele celler, i et cellelysat eller i en delvis renset eller hovedsakelig ren form. En nukleinsyre er "isolert" eller "gjort hovedsakelig ren" når renset bort fra andre cellulære komponenter eller andre forurensninger, f.eks. andre cellulære nukleinsyrer eller proteiner, ved standardteknikker, inkludert alkali/sds-behandling, CsCl binding, kolonnekromatografi, agarosegelelektroforese og andre velkjent i faget. Se, F. Ausubels, et al. red. Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987). 3 [0081] Nukleinsyresammensetningene ifølge foreliggende oppfinnelse kan, selv om de ofte er i en naturlig sekvens (med unntak av modifiserte restriksjonsseter og lignende), fra enten cdna, genomisk eller blandinger derav, bli mutert, i samsvar med standardteknikker for å tilveiebringe gensekvenser. For kodende sekvenser kan disse mutasjonene påvirke aminosyresekvens som ønsket. Spesielt er det tenkt DNA-sekvenser hovedsakelig homologe med eller avledet fra naturlig V, D, J, konstant, skifter og andre slike sekvenser beskrevet heri (hvor "avledet" indikerer at en sekvens er identisk eller modifisert fra en annen sekvens).

21 [0082] Begrepet "operativt knyttet" refererer til en nukleinsyresekvens plassert i et funksjonelt forhold med en annen nukleinsyresekvens. For eksempel er DNA for en presekvens eller sekretorisk leder operativt knyttet til DNA 1 for et polypeptid hvis det er uttrykt som et preprotein som deltar i utskillelsen av polypeptidet, en promoter eller enhancer er operativt knyttet til en kodende sekvens hvis den påvirker transkripsjonen av sekvensen, eller et ribosombindingssete er operativt knyttet til en kodende sekvens hvis det er posisjonert slik at det fremmer translasjon. Generelt betyr "operativt knyttet" at DNA-sekvensene som er knyttet sammen er tilgrensende og, i tilfelle av en sekretorisk leder, tilgrensende og i lese-fasen. Enhancere trenger imidlertid ikke å være tilgrensende. Tilknytning oppnås ved ligasjon ved praktiske restriksjonsseter. Hvis slike seter ikke eksisterer, blir de syntetiske oligonukleotidadaptere eller linkere anvendt i samsvar med konvensjonell praksis. En nukleinsyre er "operativt knyttet" når den er plassert i et funksjonelt forhold med en annen nukleinsyresekvens. For eksempel er en promoter eller enhancer operativt knyttet til en kodende sekvens hvis den påvirker transkripsjonen av sekvensen. Med hensyn til transkripsjonsregulatoriske sekvenser, betyr operativt knyttet at DNA-sekvensene som blir tilknyttet er tilgrensende og, hvor nødvendig for å føye sammen to proteinkodende regioner, tilgrensende og i leseramme. For skiftesekvenser, indikerer operativt knyttet at sekvensene er i stand til å bevirke skifterekombinering. 2 [0083] Begrepet "vektor", som anvendt heri, er ment å referere til et nukleinsyremolekyl som er i stand til å transportere en annen nukleinsyre som den har blitt knyttet til. Én type vektor er et "plasmid", som refererer til en sirkulær dobbeltrådet DNA-sløyfe som ytterligere DNA-segmenter kan bli ligert inn i. En annen type vektor er en viral vektor, hvori ytterligere DNA segmenter kan bli ligert inn i det virale genomet. Visse vektorer er i stand til autonom replikering i en vertscelle som de er innført i (f.eks. bakterielle vektorer som har et bakterielt replikasjonsopphav og episomale mammalske vektorer). Andre vektorer (f.eks. ikke-episomale mammalske vektorer) kan integreres i genomet til en vertscelle etter introduksjon i vertscellen, og blir dermed replikert sammen med vertsgenomet. Videre er visse vektorer i stand til å styre ekspresjonen av gener som de er operativt knyttet til. Slike vektorer er referert til her som "rekombinante ekspresjonsvektorer" (eller ganske enkelt, "ekspresjonsvektorer"). Generelt er ekspresjonsvektorer av nytte i rekombinante DNA-teknikker ofte i form av plasmider. Begrepene "plasmid" og "vektor" kan anvendes om hverandre. Oppfinnelsen er imidlertid tenkt å inkludere slike andre former av ekspresjonsvektorer, så som virale vektorer (f.eks. replikasjonsdefekte retrovirus, adenovirus og adenoassosierte virus), som tjener ekvivalente funksjoner. [0084] Begrepet "rekombinant vertscelle" (eller ganske enkelt "vertscelle"), som anvendt heri, er tenkt å referere 3 til en celle som en rekombinant ekspresjonsvektor har blitt introdusert i. Det skal forstås at slike begreper er tenkt å referere ikke bare til den spesielle subjektcellen men til avkommet av en slik celle. Fordi visse modifikasjoner kan forekomme i påfølgende generasjoner på grunn av enten mutasjon eller miljømessige påvirkninger, trenger slik avkom faktisk ikke å være identisk med opphavscellen, men er fremdeles inkludert innen omfanget av begrepet "vertscelle" som anvendt heri.
22 21 [008] Begrepene "behandle", "behandlende" og "behandling", som anvendt heri, refererer til terapeutiske eller forebyggende tiltak som beskrevet heri. Metodene for "behandling" anvender å administrere til et subjekt, et antistoff eller antigenbindende del ifølge foreliggende oppfinnelse, for eksempel, et subjekt som har en sykdom eller lidelse assosiert med ErbB3 avhengig signalisering eller er predisponert for å ha en slik sykdom eller lidelse, for å forebygge, kurere, forsinke, redusere alvorligheten av eller lindre ett eller flere symptomer på sykdommen eller lidelsen eller tilbakevendende sykdom eller lidelse, eller for å forlenge overlevelsen av et subjekt utover det som forventes i fravær av slik behandling. 1 [0086] Begrepet "sykdom assosiert med ErbB3 avhengig signalisering," eller "lidelse assosiert med ErbB3 avhengig signalisering," som anvendt heri, inkluderer sykdomstilstander og/eller symptomer assosiert med en sykdomstilstand, hvor økte nivåer av ErbB3 og/eller aktivering av cellulære kaskader som involverer ErbB3 er funnet. Det blir forstått at ErbB3 heterodimeriserer med andre ErbB-proteiner så som EGFR og ErbB2, når økte nivåer av ErbB3 blir funnet. Følgelig omfatter begrepet "sykdom assosiert med ErbB3-avhengig signalisering," også sykdomstilstander og/eller symptomer assosiert med sykdomstilstander hvor økte nivåer av EGFR/ErbB3 og/eller ErbB2/ErbB3 heterodimerer er funnet. Generelt refererer begrepet "sykdom assosiert med ErbB3 avhengig signalisering," til en hvilken som helst lidelse, begynnelsen, progresjon eller vedvarenheten av symptomene som krever deltakelsen av ErbB3. Eksempelvise ErbB3-medierte lidelser inkluderer, men er ikke begrenset til, for eksempel, kreft. 2 3 [0087] Begrepene "kreft" og "kreftaktig" refererer til eller beskriver den fysiologiske tilstanden hos pattedyr som typisk er kjennetegnet ved uregulert cellevekst. Eksempler på kreft inkluderer, men er ikke begrenset til karsinom, lymfom, blastom, sarkom og leukemi. Mer spesielle eksempler på slike kreftformer inkluderer plateepitelkreft, småcellet lungekreft, ikke-småcellet lungekreft, magekreft, kreft i bukspyttkjertelen, glialcelletumorer så som glioblastom og nevrofibromatose, livmorhalskreft, eggstokk-kreft, leverkreft, blærekreft, hepatom, brystkreft, colonkreft, melanom, kolorektalkreft, endometriekarsinom, spyttkjertelkarsinom, nyrekreft, renalkreft, prostatakreft, vulvalkreft, kreft i skjoldbruskkjertelen, hepatisk karsinom og ulike typer av hode- og halskreft. I en spesiell utførelsesform, blir en kreft behandlet eller diagnostisert ved anvendelse av fremgangsmåtene ifølge foreliggende oppfinnelse valgt fra melanom, brystkreft, eggstokk-kreft, renalkarsinom, gastrointestinal/colonkreft, lungekreft og prostatakreft. [0088] Begrepet "effektiv mengde", som anvendt heri, refererer til den mengden av et antistoff eller en antigenbindende del derav som binder ErbB3, som er tilstrekkelig til å bevirke behandling, prognose eller diagnose av en sykdom assosiert med ErbB3 avhengig signalisering, som beskrevet heri, når administrert til et subjekt. En terapeutisk effektiv mengde vil variere avhengig av subjektet og sykdomstilstanden som blir behandlet, vekten og alderen av subjektet, alvorlighetsgraden av sykdomstilstanden, måten for administrasjon og lignende, som enkelt kan bestemmes av en fagperson. Doseringene for administrering kan variere fra for eksempel omkring 1 ng til omkring.000 mg, omkring ng til omkring 9.00 mg, omkring ng til omkring mg, omkring ng til omkring 8.00 mg, omkring ng til omkring 7.00 mg, omkring 40 ng til omkring mg, omkring 0 ng til
23 22 omkring 6.00 mg, omkring 0 ng til omkring mg, omkring 0 ng til omkring.00 mg, omkring 0 ng til omkring.000 mg, omkring 400 ng til omkring 4.00 mg, omkring 00 ng til omkring mg, omkring 1 g til omkring 3.00 mg, omkring g til omkring mg, omkring g til omkring mg, omkring g til 1 omkring 2.7 mg, omkring g til omkring 2.0 mg, omkring 40 g til omkring 2.00 mg, omkring 0 g til omkring 2.47 mg, omkring 0 g til omkring 2.40 mg, omkring 0 g til omkring 2.42 mg, omkring 0 g til omkring 2.000, omkring 400 g til omkring 1.17 mg, omkring 00 g til omkring 1. mg, omkring 0, mg til omkring 1.12 mg, omkring 1 mg til omkring 1.0 mg, omkring 1,2 mg til omkring 1.07 mg, omkring 1, mg til omkring 1.00 mg, omkring 2,0 mg til omkring 1.02 mg, omkring 2, mg til omkring mg, omkring 3,0 mg til omkring 97 mg, omkring 3, mg til omkring 90 mg, omkring 4,0 mg til omkring 92 mg, omkring 4, mg til omkring 900 mg, omkring mg til omkring 87 mg, omkring mg til omkring 80 mg, omkring mg til omkring 82 mg, omkring mg til omkring 800 mg, omkring 40 mg til omkring 77 mg, omkring 0 mg til omkring 70 mg, omkring 0 mg til omkring 72 mg, omkring 0 mg til omkring 700 mg, omkring 0 mg til omkring 67 mg, omkring 400 mg til omkring 60 mg, omkring 00 mg eller omkring 2 mg til omkring 62 mg, av et antistoff eller antigenbindende del derav, ifølge oppfinnelsen. Doseringsregimer kan bli regulert for å gi optimal terapeutisk respons. En effektiv mengde er også én hvor eventuelle toksiske eller skadelige effekter (dvs. bivirkninger) av et antistoff eller antigenbindende del derav blir minimert, og/eller oppveid av de gunstige effektene. [0089] Begrepet "pasient" inkluderer humane og andre mammalske pasienter som mottar enten profylaktisk eller 2 terapeutisk behandling. [0090] Som anvendt heri inkluderer begrepet "subjekt" et hvilket som helst humant eller ikke-humant dyr. For eksempel kan metodene og sammensetningene ifølge foreliggende oppfinnelse bli anvendt for å behandle et subjekt som har kreft. I en spesiell utførelsesform er subjektet et menneske. Begrepet "ikke-humant dyr" inkluderer alle virveldyr f.eks. pattedyr og ikke-pattedyr, så som ikke-humane primater, sau, hund, ku, høns, amfibier, reptiler, etc. [0091] Begrepet "prøve" refererer til vev, kroppsvæske eller en celle fra en pasient eller et subjekt. Normalt vil vevet eller cellen bli fjernet fra pasienten, men in vivo diagnose er også tenkt. I tilfellet av en fast tumor, kan en vevsprøve bli tatt fra en kirurgisk fjernet tumor og preparert for testing ved konvensjonelle teknikker. I tilfellet av lymfomer og leukemier kan lymfocytter, leukemiske celler eller lymfevev bli oppnådd og passende preparert. Andre pasientprøver, inkludert urin, tåredråper, serum, cerebrospinalvæske, avføring, spytt, celleekstrakter osv. kan også være nyttige for spesielle tumorer. 3 [0092] Begrepene "anti-kreft middel" og "antineoplastisk middel" refererer til legemidler anvendt for å behandle ondartetheter, så som kreftsvulster. Legemiddelterapi kan anvendes alene, eller i kombinasjon med andre behandlinger, så som kirurgi eller stråleterapi. Flere klasser av legemidler kan bli anvendt i kreftbehandling, avhengig av naturen av det involverte organet. For eksempel blir brystkreft ofte stimulert av østrogen, og kan behandles med legemidler som inaktiverer kjønnshormoner. Likeledes kan prostatakreft behandles med
24 23 legemidler som inaktiverer androgener, det mannlige kjønnshormon. Anti-kreftmidler ifølge oppfinnelsen inkluderer blant annet følgende midler: Anti-kreft middel Kommentarer Eksempler Antistoffer (a) antistoffer andre enn anti-erbb3 antistoffer og; (b) anti-erbb3 antistoffer som binder ulike epitoper Antistoffer som binder IGF-1R (insulinlignende vekstfaktor- type 1 reseptor), som uttrykkes på celleoverflaten av de fleste humane kreftformer A12 (fullstendig humanisert mab) 19D12 (fullstendig humanisert mab) CP (fullstendig humanisert mab) H7C (humanisert mab) alfair3 (mus) scfv/fc (mus/human kimæra) EM/164 (mus) Antistoffer som binder EGFR (epidermal vekstfaktorreseptor); Mutasjoner som påvirker EGFR-ekspresjon eller aktivitet kunne resultere i kreft Antistoffer som binder cmet (mesenkymal epitelial transisjonsfaktor), et medlem av MET familien av reseptortyrosinkinaser) Matuzumab (EMD700) Erbitux /Cetuximab (Imclone) Vectibix /Panitumumab (Amgen) mab 806 Nimotuzumab (TheraCIM) AVEO (AV299) (AVEO) AMG2 (Amgen) D (OA-D) (Genentech) Små molekyler som målsøker IGF1R Anti-ErbB3-antistoffer som binder forskjellige epitoper IGF-1R (insulin-lignende vekstfaktor- type 1 reseptor), som uttrykkes på celleoverflaten av de fleste humane krefttyper Ab #14 (MM ) beskrevet heri Herceptin (Trastuzumab; Genentech) 1B4C3; 2D1D12 (U3 Pharma AG) NVP-AEW41-A BMS-36, 924 (1H-benzoimidazol-2-yl)-1Hpyridin-2-on) BMS-4, 417 Cycloligan TAE226 PQ401
25 24 Anti-kreft middel Kommentarer Eksempler Små molekyler som målsøker EGFR EGFR (epidermal vekstfaktorreseptor); Mutasjoner som påvirker EGFR-ekspresjon eller aktivitet kunne resultere i kreft Iressa /Gefitinib (AstraZeneca) CI-33 (PD 18380) (Pfizer) Lapatinib (GW-716) (GlaxoSmithKline) Tykerb /Lapatinib Ditosylat (SmithKline Beecham) Tarceva /Erlotinib HCL (OSI-774) (OSI Pharma) PKI-166 (Novartis) PD EKB-69 Tyrphostin AG 1478 (4-(3-kloranillino)-6,7- dimetoksykinazolin) Små molekyler som målsøker CMET cmet (mesenkymale epitel transisjonsfaktor), et medlem av MET familien av reseptortyrosinkinaser) PHA6672 ARQ 197 Antimetabolitter En antimetabolitt er et kjemikalie med en lignende struktur som en substans (en metabolitt) krevet for normale biokjemiske reaksjoner, men forskjellig nok til å forstyrre de normale funksjonene av celler, inkludert celledeling.
26 2 Anti-kreft middel Kommentarer Eksempler Alkyleringsmidler Et alkylerende antineoplastisk middel er et alkyleringsmiddel som knytter en alkylgruppe til DNA. Fordi kreftceller generelt prolifererer ubegrenset mer enn friske celler gjør det, er de mer følsomme for DNA-skade og alkyleringsmidler blir anvendt klinisk for å behandle en rekke tumorer. Ribonukleotid reduktaseinhibitor (RNR) Cyklofosfamid/Cytoxan (BMS) Neosar (TEVA) Ifosfamid/Mitoxana (ASTA Medica) Tiotepa (Bedford, Abraxis, Teva) BCNU 1,3-bis (2-kloretyl)-1-nitosourea CCNU 1,-(2-kloretyl)-3-cykloheksyl-1- nitrosourea (metyl-ccnu) Heksametylmelamin (Altretamin, HMM)/Hexalen (MGI Pharma Inc.) Busulfan/Myleran (GlaxoSmithKline) prokarbazin HCL/ Prokarbazin HCL / Matulan (Sigma Tau Pharmaceuticals, Inc.) Dakarbazin (DTIC) Klorambucil/Leukaran (SmithKline Beecham) Melfalan/Alkeran (GlaxoSmithKline) Cisplatin (cisplatinum, CDDP)/Platinol (Bristol Myers) Karboplatin/Paraplatin (BMS) Oksaliplatin/Eloxitan (Sanofi-Aventis US) Topoisomeraseinhibitorer Topoisomeraseinhibitorer er kjemoterapimidler designet for å forstyrre virkningen av topoisomeraseenzymer (topoisomerase I og II), som er enzymer som styrer endringene i DNA-strukturen ved å katalysere brytingen og gjentatte sammenføyningen av fosfodiesterryggraden i DNA tråder i løpet av normal cellesyklus. Doksorubicin HCL/Doxil (Alza) Daunorubicin citrat/daunoxome (Gilead) Mitoksantron HCL/Novantron (EMD Serono) Actinomycin D Etoposid/Vepesid (BMS)/Etopophos (Hospira, Bedford, Teva Parenteral, etc.) Topotekan HCL/Hycamtin (GlaxoSmithKline) Teniposid (VM-26)/Vumon (BMS) Irinotecan HCL (CPT-11)/ Camptosar (Pharmacia & Upjohn)
27 26 Anti-kreft middel Kommentarer Eksempler Mikrotubule målsøkende midler Mikrotubuler er en av komponentene i cytoskjelettet. De har diameter på ~24 nm og lengde som varierer fra flere mikrometer til muligens millimeter i aksoner av nerveceller. Mikrotubuler tjener som strukturelle komponenter innen celler og er involvert i mange cellulære prosesser, inkludert mitose, cytokinese og vesikulær transport. Vinkristin/Oncovin (Lilly) Vinblastin sulfat/velban ) (utgått) (Lilly) Vinorelbin tartrat/navelbine (PierreFabre) Vindesin sulfat/eldisine (Lilly) Paklitaxel/Taxol (BMS) Docetaxel/Taxotere (Sanofi Aventis US) Nanopartikkel paklitaxel (ABI- 007)/Abraxane (Abraxis BioScience, Inc.) Ixabepilone/IXEMPRA (BMS) Kinaseinhibitorer Tyrosinkinaser er enzymer innen cellen som fungerer for å feste fosfatgrupper til aminosyren tyrosin. Ved å blokkere evnen proteintyrosinkinaser har til å fungere, tilveiebringer disse forbindelsene et verktøy for å kontrollere kreftcellevekst Imatinibmesylat/Gleevec (Novartis) Sunitinibmaleat/Sutent (Pfizer) Sorafenib tosylat/nexavar (Bayer) Nilotinib hydroklorid monohydrat/tasigna (Novartis) Proteinsynteseinhibitorer Induserer celleapoptose L-asparaginase/Elspar (Merck & Co) Immunoterapeutiske midler Induserer at kreftpasienter utviser immunresponsivitet Alfa interferon Angiogeneseinhibitor/Avastin (Genentech) IL-2 interleukin 2 (Aldesleukin)/Proleukin (Chiron) IL-12 interleukin 12
28 27 Anti-kreft middel Kommentarer Eksempler Hormoner Hormonterapier assosiert med menopause og aldring søker å øke mengden av visse hormoner i kroppen din for å kompensere for alders- eller sykdomsrelatert hormonnedgang. Hormonterapi som en kreftbehandling vil enten redusere nivået av spesifikke hormoner eller endre kreftens evne til å anvende disse hormonene for å vokse og spre seg. Toremifencitrat/Fareston (GTX, Inc.) Fulvestrant/Faslodex (AstraZeneca) Raloksifen HCL/Evista (Lilly) Anastrazol/Arimidex (AstraZeneca) Letrozol/Femara (Novartis) Fadrozol (CGS16949A) Exemestan/Aromasin (Pharmacia & Upjohn) Leuprolid acetat/eligard (QTL USA) Lupron (TAP Pharm.) Goserelin acetat/zoladex (AstraZeneca) Triptorelin pamoat/trelstar (Watson Labs) Buserelin/Suprefact (Sanofi Aventis) Nafarelin Cetrorelix/Cetrotide (EMD Serono) Bicalutamid/Casodex (AstraZeneca) Nilutamid/Nilandron (Aventis Pharm.) Megestrolacetat/Megace (BMS) Somatostatinanaloger (Octreotid acetat/sandostatin (Novartis)) Glukokortikoider Anti-inflammatoriske legemidler anvendt for å redusere hevelse som forårsaker kreftsmerte. Predinsolon Deksametason/Decadron (Wyeth) Aromatoseinhibitorer Inkluderer imidazoler Ketokonazol mtor inhibitorer Den mtor signaliserende veien ble opprinnelig oppdaget i løpet av studier av det immundempende midlet rapamycin. Denne svært bevarte veien regulerer celleproliferasjon og metabolisme som respons til miljømessige faktorer, sammenknyttende cellevekstfaktor reseptor signalisering via fosfoinositid-3- Sirolimus (Rapamycin) / Rapamune (Wyeth) Temsirolimus (CCI-779)/Torisel (Wyeth) Deforolimus (AP2373) (Ariad Pharm.) Everolimus (RAD001)/Certican (Novartis)
29 28 Anti-kreft middel Kommentarer Eksempler kinase (PI-3K) til cellevekst, proliferasjon og angiogenese. Kjemoterapeutiske midler Adriamycin, -Fluoruracil, Cytoxin, Bleomycin, Mitomycin C, Daunomycin, karminomycin, Aminopterin, Dactinomycin, Mitomyciner, Esperamiciner [0093] Ett eller flere anti-kreft midler kan bli administrert enten samtidig eller før eller etter administrering av et antistoff eller antigenbindende del derav ifølge foreliggende oppfinnelse. [0094] Ulike aspekter av oppfinnelsen er beskrevet mer detaljert i de følgende underseksjonene. II. Fremgangsmåter for fremstilling av antistoffer ifølge oppfinnelsen 1 (I) Monoklonale antistoffer [009] Monoklonale antistoffer ifølge oppfinnelsen kan produseres ved anvendelse av en rekke kjente teknikker, så som standard somatisk celle-hybridiseringsteknikk beskrevet ved Kohler og Milstein (197) Nature 26: 49, viral eller onkogen transformasjon av B-lymfocytter eller fag display-teknikk ved anvendelse av biblioteker av humane antistoffgener. I spesielle utførelsesformer er antistoffene fullstendig humane monoklonale antistoffer. [0096] Følgelig blir, i en utførelsesform, en hybridommetode anvendt for fremstilling av et antistoff som binder ErbB3. I denne metoden, kan en mus eller annet passende vertsdyr bli immunisert med et egnet antigen for å fremkalle lymfocytter som produserer eller er i stand til å produsere antistoffer som spesifikt vil binde til antigenet anvendt for immunisering. Alternativt kan lymfocytter bli immunisert in vitro. Lymfocytter kan så bli fusjonert med myelomceller ved anvendelse av et egnet fusjonsmiddel, så som polyetylenglykol, for å danne en hybridomcelle (Goding, Monoclonal Antibodies: Principles and Practice, pp. 9-3 (Academic Press, 1986)). Kulturmedium som hybridomceller vokser i blir analysert for produksjon av monoklonale antistoffer rettet mot antigenet. Etter at hybridomceller er identifisert som produserer antistoffer med den ønskede spesifisiteten, affiniteten og/eller aktiviteten, kan klonene bli subklonet ved begrensende fortynning prosedyrer og dyrket ved standardmetoder
30 29 (Goding, Monoclonal Antibodies: Principles and Practice, pp. 9-3 (Academic Press, 1986)). Egnede kulturmedier for dette formålet inkluderer, for eksempel D-MEM eller RPMI-1640 medium. I tillegg kan hybridomcellene dyrkes in vivo som ascitestumorer i et dyr. De monoklonale antistoffene utskilt av subklonene kan skilles fra kulturmediet, ascitesfluidet eller serum ved konvensjonelle immunglobulin renseprosedyrer så som for eksempel protein A- sefarose, hydroksylapatitt kromatografi, gelelektroforese, dialyse eller affinitetskromatografi. [0097] I en annen utførelsesform kan antistoffer og antistoffdeler som binder ErbB3 isoleres fra antistoff-fagbiblioteker generert ved anvendelse av teknikkene beskrevet i, for eksempel, McCafferty et al. Nature, 348:2-1 4 (1990). Clackson et al. Nature, 32: (1991), Marks et al. J. Mol. Biol. 222:81-97 (1991) og Hoet et al (0) Nature Biotechnology 23, ; U.S. Patenter nr.,223,409;,403,484 og,71,698 til Ladner et al.; U.S. Patenter nr.,427,908 og,80,717 til Dower et al.; U.S. Patenter nr.,969,8 og 6,172,197 til McCafferty et al.; og U.S. Patenter nr.,88,793; 6,21,404; 6,44,731; 6,,313; 6,82,91 og 6,93,081 til Griffiths et al. I tillegg kan produksjon av høyaffinitets- (nm område) humane antistoffer ved kjedestokking (Marks et al. Bio/Technology, : (1992)), så vel som kombinatorisk infeksjon og in vivo rekombinering som en strategi for å konstruere svært store fag-biblioteker (Waterhouse et al. Nuc. Acids. Res. 21: (1993)) også benyttes. [0098] I en spesiell utførelsesform blir det monoklonale antistoffet eller antigenbindende del derav som binder ErbB3 produsert ved anvendelse av fag display-teknikken beskrevet av Hoet et al. supra. Denne teknikken involverer genereringen av et humant Fab-biblioteket som har en unik kombinasjon av immunoglobulinsekvenser isolert fra humane donorer og som har syntetisk diversitet i tung-kjede-cdrene blir generert. Biblioteket blir så screenet for Fabs som binder til ErbB3. 2 [0099] I enda en annen utførelsesform, kan humane monoklonale antistoffer rettet mot ErbB3 bli generert ved anvendelse av transgene eller transkromosome mus som bærer deler av det humane immunsystemet snarere enn musesystemet (se f.eks. Lonberg, et al. (1994) Nature 368 (6474): 86-89; Lonberg, N. et al.(1994), supra; gjennomgått i Lonberg, N. (1994) Handbook of Experimental Pharmacology 113:49-1; Lonberg, N. og Huszar, D. (199) Intern. Rev Immunol. 13: 6-93, og Harding, F. og Lonberg, N. (199) Ann. N.Y. Acad Sci. 764: Se videre U.S. Patenter nr.,4,806;,69,82;,62,126;,633,42;,789,60;,877,397;,661,016;,814,318;,874,299 og,770,429, alle til Lonberg og Kay; U.S. Patent nr.,4,807 til Surani et al.; PCT publikasjoner nr. WO 92/03918, WO 93/12227, WO 94/28, WO 97/1382, WO 98/24884 og WO 99/4962, alle til Lonberg og Kay, og PCT publikasjon nr. WO 01/14424 til Korman et al.). 3 [00] I en annen utførelsesform, kan humane antistoffer ifølge oppfinnelsen bli øket ved anvendelse av en mus som bærer humane immunoglobulinsekvenser på transgener og transkromosomer, så som en mus som bærer et humant tung kjede transgen og et humant lett kjede transkromosom (se f.eks. PCT publikasjon WO 02/43478 til Ishida et al.).
31 [01] Enda videre er alternative transgene dyresystemer som uttrykker humane immunglobulingener tilgjengelige innen faget og kan anvendes for å øke anti-erbb3 antistoffer ifølge oppfinnelsen. For eksempel kan et alternativt transgent system referert til som Xenomouse (Abgenix, Inc.) bli anvendt; slike mus er beskrevet i, for eksempel, U.S. Patenter nr.,939,98; 6,07,181; 6,114,98; 6,,84 og 6,162,963 til Kucherlapati et al. [02] Videre er alternative transkromosomiske dyresystemer som uttrykker humane immunglobulingener tilgjengelige innen faget og kan anvendes for å øke anti-erbb3 antistoffene ifølge oppfinnelsen. For eksempel kan mus som bærer både et humant tung kjede transkromosom og et humant lett kjede transkromosom bli anvendt, som beskrevet i Tomizuka et al. (00) Proc. Natl. Acad Sci. USA 97: Dessuten har kuer som bærer humane tung- og lett kjede transkromosomer blitt beskrevet i teknikken (Kuroiwa et al. (02) Nature Biotechnology : ) og kan anvendes for å øke anti-erbb3 antistoffer ifølge oppfinnelsen. 1 2 [03] I enda en annen utførelsesform kan antistoffer ifølge foreliggende oppfinnelse bli fremstilt ved anvendelse av en transgen plante og/eller kultiverte planteceller (så som, for eksempel, tobakk, mais og andemat) som produserer slike antistoffer. For eksempel kan transgene tobakksblader som uttrykker antistoffer eller antigenbindende deler derav anvendes for å produsere slike antistoffer ved, for eksempel, anvendelse av en induserbar promoter (se, f.eks. Cramer et al. Curr. Top. Microbol. Immunol. 240:9 118 (1999)). Transgen mais kan også anvendes for å uttrykke slike antistoffer og antigenbindende deler derav (se, f.eks. Hood et al. Adv. Exp. Med Biol. 464: (1999)). Antistoffer kan også bli produsert i store mengder fra transgene plantefrø inkludert antistoffdeler, så som enkeltkjede-antistoffer (scfv-er), for eksempel ved anvendelse av tobakksfrø og potetknoller (se f.eks. Conrad et al. Plant Mol. Biol. 38:1 9 (1998)). Fremgangsmåter for fremstilling av antistoffer eller antigenbindende deler i planter kan også bli funnet i, f.eks. Fischer et al. Biotechnol. Appl. Biochem. :99 8 (1999), Ma et al. Trends Biotechnol. 13:22 7 (199); Ma et al. Plant Physiol. 9:341 6 (199); Whitelam et al. Biochem. Soc. Trans. 22: (1994) og U.S. Patenter nr. 6,040,498 og 6,81,184. [04] Bindingsspesifisiteten for monoklonale antistoffer eller deler derav som binder ErbB3 fremstilt ved anvendelse av en hvilken som helst teknikk, inkludert de beskrevet her, kan bli bestemt ved immunoutfelling eller ved en in vitro bindingsanalyse, så som radioimmunoanalyse (RIA) eller enzym-koblet immunoabsorbent analyse (ELISA). Bindingsaffiniteten for et monoklonalt antistoff eller del derav kan også bli bestemt ved Scatchardanalysen ifølge Munson et al. Anal. Biochem. 7:2 (1980). [0] I visse utførelsesformer kan et ErbB3 antistoff eller del derav fremstilt ved anvendelse av en hvilken som 3 helst av metodene diskutert over bli videre endret eller optimalisert for å oppnå en ønsket bindingsspesifisitet og/eller affinitet ved anvendelse av faglig anerkjente teknikker, så som de beskrevet heri. [06] I én utførelsesform kan delvise antistoff-sekvenser avledet fra et ErbB3 antistoff anvendes for å gi strukturelt og funksjonelt beslektede antistoffer. For eksempel vekselvirker antistoffer med mål-antigener hovedsakelig gjennom aminosyrerester som er lokalisert i de seks tung og lett kjede
32 31 komplementaritetsbestemmende regionene (CDRer). Av denne grunn, er aminosyresekvensene innen CDRer mer mangfoldige mellom individuelle antistoffer enn sekvenser utenfor CDRer. Fordi CDR-sekvensene er ansvarlige for de fleste antistoff-antigen-vekselvirkninger, er det mulig å uttrykke rekombinante antistoffer som etterligner egenskapene av spesifikke naturlig forekommende antistoffer ved å konstruere ekspresjonsvektorer som inkluderer CDR-sekvenser fra det spesifikke naturlig forekommende antistoffet podet på rammeverksekvenser fra et forskjellig antistoff med forskjellige egenskaper (se, f.eks. Riechmann, L. et al. 1998, Nature 332: ; Jones, P. et al. 1986, Nature 321:22-2, og Queen, C. et al. 1989, Proc. Natl. Acad See. USA 86: ). Slike rammeverksekvenser kan oppnås fra offentlige DNA-databaser som inkluderer kimlinje antistoff gensekvenser. 1 [07] Således kan ett eller flere strukturelle trekk ved et anti-erbb3-antistoff ifølge oppfinnelsen, så som CDRene, anvendes for å skape strukturelt beslektede anti-erbb3-antistoffer som bevarer minst én funksjonell egenskap for antistoffene ifølge oppfinnelsen, f.eks. inhibering av EGF-lignende ligandmediert fosforylering av ErbB3; inhibering av én eller flere av heregulin, epiregulin, epigen eller biregulin-mediert signalisering gjennom ErbB3; inhibering av proliferasjon eller celler som uttrykker ErbB3; og/eller reduksjon av nivåer av ErbB3 på celleoverflater. [08] I en spesiell utførelse blir én eller flere CDR regioner valgt fra SEKV ID NR :7-12, SEKV ID NR: 13-18, SEKV IDnummer: 19-24, SEKV ID NR: 39-44, og SEKV ID-nummer: 4-0 kombinert rekombinant med kjente humane rammeverkregioner og CDRer for å skape ytterligere, rekombinant-utviklede, anti-erbb3 antistoffer ifølge oppfinnelsen. Tung og lett kjede variable rammeområdene kan bli avledet fra de samme eller forskjellige antistoffsekvenser. [09] Det er velkjent innen faget at antistoff-tung- og lett-kjede CDR3-domener spiller en spesielt viktig rolle i 2 bindingsspesifisiteten/affiniteten av et antistoff for et antigen (se, Hall et al. J. Imunol., 149: (1992); Polymenis et al. J. Immunol., 12: (1994); Jahn et al. Immunobiol., 193: (199); Klimka et al. Brit. J. Cancer, 83: (00); Beiboer et al. J. Mol. Biol, 296: (00); Rader et al. Proc. Natl. Acad Sci. USA, 9: (1998); Barbas et al. J. Am. Chem. Soc., 116: (1994); Ditzel et al. J. Immunol., 17: (1996)). I visse utførelsesformer, blir følgelig antistoffer generert som inkluderer tung og/eller lett kjede CDR3ene av de spesielle antistoffene beskrevet heri (f.eks. SEKV ID NR: 9, 1, 21, 41, 47 og/eller SEKV ID NR: 12, 18, 24, 44, 0). Antistoffene kan videre inkludere tung og/eller lett kjede CDR1 og/eller CDR2ene av antistoffene ifølge foreliggende oppfinnelse (f.eks. SEKV ID NR: 7-8 og/eller SEKV ID NR: -11, SEKV ID NR: og/eller SEKV ID NR: 16-17, SEKV ID NR: -21 og/eller SEKV ID NR: 22-23, SEKV ID NR: og/eller SEKV ID NR: 42-43; eller SEKV ID NR: 4-46 og/eller SEKV ID NR: 48-49). 3 [01] CDR1, 2 og/eller 3 regionene av de utviklede antistoffene beskrevet over kan omfatte de(n) nøyaktige aminosyresekvensen(e) som de fremlagt heri (f.eks. CDRer ifølge Ab #6, Ab #3, Ab #14, Ab # 17 eller Ab #19, fremlagt i henholdsvis SEKV ID NR :7-12, 13-18, 19-24, og 4-0). Den vanlige fagmannen vil imidlertid erkjenne at noe avvik fra de eksakte CDR-sekvensene kan være mulig, mens en fremdeles bevarer evnen antistoffet har til å binde ErbB3 effektivt (f.eks. konservative aminosyresubstitusjoner). I en annen utførelsesform
33 32 kan følgelig det konstruerte antistoffet være sammensatt av én eller flere CDRer som er, for eksempel 90 %, 9 %, 98 %, 99 % eller 99, % identisk med én eller flere CDRer ifølge Ab #6, Ab #3 eller Ab 14. [0111] I en annen utførelsesform kan én eller flere rester av en CDR bli endret for å modifisere binding for å oppnå en mer foretrukket "on-rate" for binding. Ved anvendelse av denne strategien kan et antistoff som har ultrahøy bindingsaffinitet på, for eksempel, M -1 eller mer bli oppnådd. Affinitetsmodningsteknikker velkjent innen faget og de beskrevet heri, kan bli anvendt for å endre CDR-regionen(e) fulgt av screening av de resulterende bindende molekylene for den ønskede endringen i bindingen. Følgelig, ettersom CDR(er) blir endret, kan endringer i bindingsaffinitet, så vel som immunogenisitet bli overvåket og vurdert slik at et antistoff optimalisert for den beste kombinerte bindingen og lave immunogenisitet blir oppnådd. [0112] I tillegg til, eller i stedet for modifikasjoner innen CDRene, kan modifikasjoner også gjøres innen ett eller 1 2 flere av rammeverkområdene, FR1, FR2, FR3 og FR4, av de tunge og/eller lette kjede variable regionene av et antistoff, så lenge disse modifikasjonene ikke eliminerer bindingsaffiniteten av antistoffet. [0113] I en annen utførelsesform blir antistoffet ytterligere modifisert med hensyn til effektorfunksjon, for å forbedre effektiviteten av antistoffet ved behandling av kreft, for eksempel. For eksempel kan cysteinrest(er) bli innført i Fc-regionen, og dermed tillate dannelse av disulfidbinding mellom kjedene i denne regionen. Det homodimere antistoffet generert slik kan ha forbedret internaliseringsevne og/eller økt komplement-mediert celledreping og antistoff-avhengig cellulær cytotoksisitet (ADCC). Se Caron et al. J. Exp. Med. 176: (1992) og Shopes, B.J. Immunol. 148: (1992). Homodimere antistoffer med forbedret anti-tumoraktivitet kan også fremstilles ved anvendelse av heterobifunksjonelle kryssbindere som beskrevet i Wolff et al. Cancer Research 3: (1993). Alternativt kan det bli utviklet et antistoff som har doble Fc regioner, og derved kan ha økt komplement lysis og ADCC evner. Se Stevenson et al. Anti-Cancer Drug Design 3:219-2 (1989). [0114] Også omfavnet ved foreliggende oppfinnelse er bispesifikke antistoffer og immunkonjugater, som diskutert nedenfor. (ii) Bispesifikke antistoffer [011] Bispesifikke antistoffer ifølge foreliggende oppfinnelse inkluderer minst én bindingsspesifisitet for ErbB3 og minst én bindingsspesifisitet for et annet antigen, så som produktet av et onkogen. Bispesifikke antistoffer kan fremstilles som full-lengde antistoffer eller antistoff-fragmenter (f.eks. F(ab') 2 bispesifikke antistoffer). 3 [0116] Fremgangsmåter for fremstilling av bispesifikke antistoffer er velkjent innen faget (se f.eks. WO og WO ). For eksempel kan produksjon av full-lengde bispesifikke antistoffer være basert på samekspresjon av to immunoglobulin tung kjede-lett kjede-par, hvor de to kjedene har forskjellige spesifisiteter (se, f.eks. Millstein et al. Nature, :37-39 (1983)). Ytterligere detaljer ved generering av bispesifikke antistoffer kan finnes, for eksempel, i Suresh et al. Methods in Enzymology, 121:2 (1986) og i Brennan et al., Science, 229:81
34 33 (198), som beskriver en kjemisk bindingsprosess for å danne bispesifikke antistoffer. Ulike teknikker for fremstilling og isolering av bispesifikke antistoffragmenter direkte fra rekombinant cellekultur har også blitt beskrevet. For eksempel har bispesifikke antistoffer blitt produsert ved anvendelse av leucin-zippere (se f.eks. Kostelny et al. J. Immunol., 148(): (1992)). En annen strategi for fremstilling av bispesifikke antistoffragmenter ved anvendelsen av enkelt-kjede-fv (sfv)-dimerer har også blitt rapportert (se, f.eks. Gruber et al. J. Immunol., 12:368 (1994)). [0117] I en spesiell utførelsesform omfatter det bispesifikke antistoffet et første antistoff eller bindende del derav som binder til ErbB3 og et andre antistoff eller bindende del derav som binder til ErbB2, ERbB3, ErbB4, EGFR, IGF1- R, C-MET, Lewis Y, MUC-1, EpCAM, CA12, prostataspesifikt membranantigen, PDGFR-α, PDGFR-β, C-KIT, eller en hvilken som helst av FGF-reseptorene. (iii) Immunkonjugater 1 2 [0118] Immunkonjugater ifølge foreliggende oppfinnelse kan dannes ved konjugering av antistoffene eller antigenbindende deler derav beskrevet heri til et annet terapeutisk middel. Egnede midler inkluderer, for eksempel, et cytotoksisk middel (f.eks. en kjemoterapeutisk middel), et toksin (f.eks. et enzymatisk aktivt toksin av bakterielt, fungalt, plante eller animalsk opprinnelse, eller fragmenter derav), og/eller en radioaktiv isotop (dvs. et radiokonjugat). Kjemoterapeutiske midler nyttige i genereringen av slike immunkonjugater har blitt beskrevet over. Enzymatisk aktive toksiner og fragmenter derav som kan anvendes inkluderer difteri A-kjede, ikke-bindende aktive fragmenter av difteritoksin, eksotoksin A kjede (fra Pseudomonas aeruginosa), ricin A kjede, abrin A kjede, modeccin A kjede, alfa-sarcin, Aleurites fordii proteiner, diantinproteiner, Phytolaca americana proteiner (PAPI, PAPII og PAP-S), momordica charantia inhibitor, curcin, crotin, sapaonaria officinalis inhibitor, gelonin, mitogellin, restrictocin, fenomycin, enomycin og trikotekenene. En rekke radionuklider er tilgjengelige for produksjonen av anti-radiokonjugerte ErbB3 antistoffer. Eksempler inkluderer 212 Bi, 131 I, 131 In, 90 Y og 186 Re. [0119] Immunkonjugatene ifølge oppfinnelsen kan bli dannet ved anvendelse av en rekke bifunksjonelle 3 proteinkoplingsmidler så som N-succinimidyl-3-(2-pyridylditiol) propionat (SPDP), iminotiolan (IT), bifunksjonelle derivater av imidoestere (så som dimetyl adipimidat HCL), aktive estere (så som disuccinimidyl suberat), aldehyder (så som glutaraldehyd), bis-azido-forbindelser (så som bis (p-azidobenzoyl)heksandiamin), bis-diazonium-derivater (så som bis-(p-diazoniumbenzoyl)-etylendiamin), diisocyanater (så som tolyen 2,6-diisocyanat) og bis-aktive fluorforbindelser (så som 1,-difluor-2,4-dinitrobenzen). For eksempel kan et ricin-immunotoksin bli fremstilt som beskrevet i Vitetta et al, Science 238: 98 (1987). Karbon-14-merket 1-isotiocyanatobenzyl-3-metyldietylen triaminpentaeddiksyre (MX-DTPA) er et eksempel på chelatdannere for konjugering av radionukleotid til antistoffet (se, f.eks. WO94/126)
35 34 III. Fremgangsmåter for screening av antistoffer ifølge oppfinnelsen [01] Etter produksjon av antistoffer eller antigenbindende deler som binder ErbB3, kan slike antistoffer, eller deler derav bli screenet for ulike egenskaper, så som de beskrevet heri, ved anvendelse av en rekke analyser som er velkjent innen faget. [0121] I én utførelsesform blir antistoffene eller antigenbindende deler derav screenet for evnen til å inhibere EGF-lignende ligandmediert fosforylering av ErbB3. Dette kan gjøres ved å behandle celler som uttrykker ErbB3 med en EGF-lignende ligand i nærværet og fraværet av antistoffet eller antigenbindende del derav. Cellene kan så bli lysert, og de ubearbeidede lysatene kan bli sentrifugert for å fjerne uløselig materiale. ErbB3-fosforylering kan bli målt, for eksempel ved Western blott fulgt av undersøkelse med et anti-fosfotyrosin-antistoff som beskrevet i Kim et al. supra og eksemplene under. 1 [0122] I andre utførelsesformer blir antistoffene og antigenbindende deler videre screenet for én eller flere av de følgende egenskapene: (1) inhibering av ErbB3-ligand (f.eks. heregulin, epiregulin, epigen eller biregulin) mediert signalisering gjennom ErbB3, (2) inhibering av proliferasjon av celler som uttrykker ErbB3, (3) evnen til å redusere nivåer av ErbB3 på celleoverflate (f.eks. ved å indusere internalisering av ErbB3), (4) inhibering av VEGF-sekresjon av celler som uttrykker ErbB3, () inhibering av migreringen av celler som uttrykker ErbB3, (6) inhibering av sfæroidevekst av celler som uttrykker ErbB3, og/eller (7) binding til en epitope lokalisert på domene I av ErbB3, hver av disse kan lett måles ved anvendelse av faglig anerkjente teknikker og de som er diskutert heri. [0123] Inhibering av én eller flere av heregulin, epiregulin, epigen eller biregulin-mediert signalisering gjennom 2 3 ErbB3 kan enkelt måles ved anvendelse av rutineanalyser, så som beskrevet i Horst et al. supra. For eksempel kan evnen et antistoff eller antigenbindende del derav har til å inhibere heregulin, epiregulin, epigen eller biregulinmediert signalisering gjennom ErbB3 måles ved kinaseanalyser for kjente substrater av ErbB3 slik som, for eksempel, SHC og PI3K, som beskrevet i, for eksempel Horst et al. supra, Sudo et al. (00) Methods Enzymol, 322:388-92, og Morgan et al. (1990) Eur. J. Biochem., 191: , etter stimulering ved én eller flere av heregulin, epiregulin, epigen eller biregulin. Følgelig kan celler som uttrykker ErbB3 bli stimulert med én eller flere av heregulin, epiregulin, epigen eller biregulin, og inkubert med et kandidat-antistoff eller antigenbindende del derav. Cellelysater påfølgende fremstilt fra slike celler kan bli immunoutfelt med et antistoff for et substrat av ErbB3 (eller et protein i en cellulær vei som involverer ErbB3) så som for eksempel et anti-jnk-1-antistoff, og analysert for kinaseaktivitet (f.eks. JNK kinaseaktivitet eller PI3-kinaseaktivitet) ved anvendelse av faglig anerkjente teknikker. En reduksjon i eller fullstendig forsvinning i nivå eller aktivitet (f.eks. kinaseaktivitet) av et ErbB3 substrat eller protein i en vei som involverer ErbB3 i nærværet av antistoffet eller antigenbindende del derav, i forhold til nivået eller aktiviteten i fraværet av antistoffet eller antigenbindende del derav, er indikerende for et antistoff eller antigenbindende del som inhiberer én eller flere av heregulin, epiregulin, epigen eller biregulin-mediert signalisering.
36 3 [0124] I visse utførelsesformer inhiberer antistoffet eller antigenbindende del derav ErbB3-ligand (f.eks. heregulin, epiregulin, epigen eller biregulin) mediert signalisering ved å redusere bindingen av ett eller flere av heregulin, epiregulin, epigen eller biregulin til ERbB3. [012] For å velge de antistoffene eller antigenbindende deler derav som inhiberer bindingen av én eller flere av heregulin, epiregulin, epigen eller biregulin til ErbB3, kan celler som uttrykker ErbB3 (f.eks. MALME-3M celler, som beskrevet i eksemplene infra), bli brakt i kontakt med en merket ErbB3-ligand (f.eks. radiomerket heregulin, epiregulin, epigen eller biregulin) i fraværet (kontroll) eller nærværet av anti-erbb3-antistoffet eller antigenbindende del derav. Hvis antistoffet eller antigenbindende del derav inhiberer heregulin-, epiregulin-, epigen- eller biregulinbinding til ErbB3, så vil det bli observert en statistisk signifikant reduksjon i mengden av merke gjenvunnet (f.eks. radiomerket heregulin, epiregulin, epigen eller biregulin), i forhold til mengden i fraværet av antistoffet eller antigenbindende del derav [0126] Antistoffet eller antigenbindende del derav kan inhibere bindingen av ErbB3-liganden (f.eks. heregulin, epiregulin, epigen eller biregulin) ved en hvilken som helst mekanisme. For eksempel kan antistoffet eller antigenbindende del derav inhibere binding av ErbB3-liganden (f.eks. én eller flere av heregulin, epiregulin, epigen eller biregulin) til ErbB3 ved binding til det samme setet eller et overlappende sete på ErbB3 som ErbB3-liganden. Alternativt kan antistoffet eller antigenbindende del derav inhibere binding av en ErbB3-ligand ved å endre eller fordreie konformasjonen av ErbB3, slik at det ikke er i stand til å binde til ErbB3-liganden. [0127] Antistoffer og antigenbindende deler derav som reduserer nivåer av ErbB3 på celleoverflater kan identifiseres ved deres evne til å nedregulere ErbB3 på tumorceller. I visse utførelsesformer reduserer antistoffene eller antigenbindende deler derav ErbB3 celleoverflate-ekspresjon ved å indusere internalisering (eller øke endocytose) av Erbb3. For å teste dette, kan ErbB3 bli biotinylert og antallet ErbB3 molekyler på celleoverflaten, kan lett bestemmes, for eksempel ved å måle mengden biotin på et monolag av celler i kultur i nærværet eller fraværet av et antistoff eller antigenbindende del derav, for eksempel som beskrevet i, f.eks. Waterman et al. J. Biol. Chem. (1998), 273: , fulgt av immunoutfelling av ErbB3 og undersøkelse med streptavidin. En reduksjon i deteksjon av biotinylert ErbB3 over tid i nærværet av et antistoff eller antigenbindende del er indikerende for et antistoff som reduserer ErbB3 nivåer på celleoverflater. [0128] Antistoffer eller antigenbindende deler derav ifølge foreliggende oppfinnelse kan også bli testet for deres evne til å inhibere proliferasjon av celler som uttrykker ErbB3, for eksempel tumorceller, ved anvendelse av faglig anerkjente teknikker, så som celletiter glød-analysen beskrevet i eksemplene under (se også, f.eks. Macallan et al. Proc. Natl. Acad Sci. (1998), 9 (2): ; Perez et al. (199) Cancer Research, ). [0129] I en annen utførelsesform blir antistoffene eller antigenbindende deler derav screenet for evnen til å inhibere VEGF-sekresjon fra celler som uttrykker ErbB3. Dette kan gjøres ved anvendelse av velkjente analyser, så som VEGF ELISA kitet tilgjengelig fra R&D Systems (Minneapolis, MN, kat.# DY293B). Likeledes kan antistoffene
37 36 eller deler bli screenet for evnen til å inhibere migreringen av celler som uttrykker ErbB3 (f.eks. MCF-7 celler) ved anvendelse av en transwell-brønn analyse (Millipore Corp, Billerica, MA, kat # ECM2) som beskrevet heri. [01] I en annen utførelsesform blir antistoffene eller antigenbindende deler derav screenet for evnen til å inhibere sfæroidevekst av celler som uttrykker ErbB3. Dette kan gjøres ved anvendelse av en analyse som tilnærmes betingelser for en utviklende tumorvekst (se f.eks. Herman et al. (07) Journal of Biomolecylar Screening elektronisk publikasjon) som beskrevet heri. 1 [0131] Antistoffer eller antigenbindende deler derav som binder til de samme eller overlappende epitoper som ett eller flere antistoffer ifølge foreliggende oppfinnelse kan også identifiseres ved anvendelse av standardteknikker som er kjent innen faget og beskrevet heri. For eksempel, for å screene for antistoffer som binder til den samme eller en overlappende epitope på ErbB3 bundet ved et interessant antistoff, kan det bli gjennomført en kryssblokkerings-analyse, så som den beskrevet i Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow og David Lane (1988). IV. Farmasøytiske sammensetninger [0132] I et annet aspekt tilveiebringer foreliggende oppfinnelse en sammensetning, f.eks. en farmasøytisk sammensetning, som inneholder ett eller en kombinasjon av monoklonale antistoffer, eller antigenbindende del(er) derav, ifølge foreliggende oppfinnelse, formulert sammen med en farmasøytisk akseptabel bærer. I én utførelsesform inkluderer sammensetningene en kombinasjon av flere (f.eks. to eller flere) isolerte antistoffer ifølge oppfinnelsen, som binder forskjellige epitoper på ErbB3. 2 [0133] Som anvendt heri inkluderer "farmasøytisk akseptabel bærer" et hvilket som helst og alle løsemidler, dispergeringsmedier, belegg, antibakterielle og antifungale midler, isotoniske og absorpsjonsforsinkende midler, og lignende som er fysiologisk kompatible. Fortrinnsvis er bæreren egnet for intravenøs, intramuskulær, subkutan, parenteral, spinal eller epidermal administrering (f.eks. ved injeksjon eller infusjon). Avhengig av administrasjonsruten, kan den aktive forbindelsen, dvs. antistoffet, bispesifikt og multispesifikt molekyl, være belagt i et materiale for å beskytte forbindelsen mot virkningen av syrer og andre naturlige betingelser som kan inaktivere forbindelsen. [0134] Et "farmasøytisk akseptabelt salt" refererer til et salt som bevarer den ønskede biologiske aktiviteten av 3 opphavsforbindelsen og ikke gir noen uønskede toksikologiske effekter (se f.eks. Berge S.M., et al. (1977) J. Pharm. Sci. 66:1-19). Eksempler på slike salter inkluderer syreaddisjonssalter og baseaddisjonssalter. Syreaddisjonssalter inkluderer de avledet fra ikke-toksiske uorganiske syrer, så som saltsyre, salpetersyre, fosforsyre, svovelsyre, hydrogenbromidsyre, hydrogenjodidsyre, fosforsyrling og lignende, så vel som fra ikke-toksiske organiske syrer så som alifatiske mono- og dikarboksylsyrer, fenylsubstituerte alkansyrer, hydroksy-alkansyrer, aromatiske syrer, alifatiske og aromatiske sulfonsyrer og lignende. Baseaddisjonssalter inkluderer de avledet fra jordalkalimetaller,
38 37 så som natrium, kalium, magnesium, kalsium og lignende, så vel som fra ikke-toksiske organiske aminer, så som N,N'-dibenzyletylendiamin, N-metylglukamin, klorprokain, kolin, dietanolamin, etylendiamin, prokain og lignende. [013] Farmasøytiske sammensetninger ifølge oppfinnelsen kan bli administrert alene eller i kombinasjonsterapi, dvs. kombinert med andre midler. For eksempel kan kombinasjonsterapien inkludere en sammensetning ifølge foreliggende oppfinnelse med minst én eller flere ytterligere terapeutiske midler, så som anti-kreft legemidlene beskrevet infra. De farmasøytiske sammensetningene ifølge oppfinnelsen kan også bli administrert i forbindelse med strålebehandling og/eller kirurgi. 1 [0136] En sammensetning ifølge foreliggende oppfinnelse kan administreres ved en rekke metoder kjent innen faget. Som det vil erkjennes av den erfarne fagmannen, vil ruten og/eller måten for administrering variere avhengig av de ønskede resultatene. De aktive forbindelsene kan bli fremstilt med bærere som vil beskytte forbindelsen mot rask frigjøring, så som en kontrollert-frigivelses-formulering, inkludert implantater, transdermale plastre og mikroinnkapslede leveringssystemer. Biologisk nedbrytbare, biologisk kompatible polymerer kan anvendes, så som etylenvinylacetat, polyanhydrider, polyglykolsyre, kollagen, polyortoestere og polymelkesyre. Mange fremgangsmåter for fremstillingen av slike formuleringer er patentert eller generelt kjent for fagpersoner. Se for eksempel, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, red., Marcel Dekker, Inc., New York, [0137] For å administrere en forbindelse ifølge oppfinnelsen ved visse administrasjonsruter, kan det være nødvendig å belegge forbindelsen med, eller sam-administrere forbindelsen med, et materiale for å forhindre dens inaktivering. For eksempel kan forbindelsen bli administrert til et subjekt i en egnet bærer, for eksempel, liposomer, eller et fortynningsmiddel. Farmasøytisk akseptable fortynningsmidler inkluderer saltvann og vandige bufferløsninger. Liposomer inkluderer vann-i-olje-i-vann CGF-emulsjoner så vel som konvensjonelle liposomer (Strejan et al. (1984) J. Neuroimmunol. 07:27). [0138] Farmasøytisk akseptable bærere inkluderer sterile vandige løsninger eller dispersjoner og sterile pulvere for den ekstemporerte fremstillingen av sterile, injiserbare løsninger eller dispersjon. Anvendelsen av slike medier og midler for farmasøytisk aktive substanser er kjent innen faget. Bortsett fra i den utstrekning som hvilke som helst konvensjonelle media eller middel er inkompatibelt med den aktive forbindelsen, er anvendelse derav i de farmasøytiske sammensetningene ifølge oppfinnelsen tenkelig. Supplerende aktive forbindelser kan også bli inkorporert i sammensetningene. 3 [0139] Terapeutiske sammensetninger må typisk være sterile og stabile under betingelsene for fremstilling og lagring. Sammensetningen kan bli formulert som en løsning, mikroemulsjon, liposom eller annen ordnet struktur egnet for høy-legemiddel-konsentrasjonen. Bæreren kan være et løsemiddel eller dispersjonsmedium som inneholder for eksempel vann, etanol, polyol (for eksempel glyserol, propylenglykol og flytende polyetylenglykol og lignende), og egnede blandinger derav. Den riktige fluiditeten kan opprettholdes, for eksempel, ved
39 38 anvendelsen av et belegg så som lecitin, ved opprettholdelse av den krevede partikkelstørrelsen i tilfellet av dispersjon og ved anvendelsen av surfaktanter. I mange tilfeller vil det være foretrukket å inkludere isotoniske midler, for eksempel sukkere, polyalkoholer så som mannitol, sorbitol eller natriumklorid i sammensetningen. Langvarig absorpsjon av de injiserbare sammensetningene kan frembringes ved å inkludere i sammensetningen et middel som forsinker absorpsjon, for eksempel, monostearatsalter og gelatin. [0140] Sterile, injiserbare løsninger kan bli fremstilt ved å inkorporere den aktive forbindelsen i den krevede mengden i et passende løsemiddel med én eller en kombinasjon av ingredienser listet opp over, ettersom krevet, fulgt av steriliseringsmikrofiltrering. Generelt fremstilles dispersjoner ved å inkorporere den aktive forbindelsen i et sterilt vehikkel som inneholder et grunnleggende dispersjonsmedium og de krevede andre ingredienser fra de listet opp over. I tilfellet med sterile pulvere for fremstillingen av sterile, injiserbare løsninger, er de foretrukne fremgangsmåter for fremstilling vakuumtørking og frysetørking (lyofilisering) som gir et pulver av den aktive ingrediensen pluss en hvilken som helst ytterligere ønsket ingrediens fra en tidligere steril-filtrert løsning derav. 1 2 [0141] Doseringsregimer blir justert for å tilveiebringe den optimale ønskede respons (f.eks. en terapeutisk respons). For eksempel kan en enkelt bolus bli administrert, flere oppdelte doser kan bli administrert over tid eller dosen kan bli proporsjonalt redusert eller øket som indikert ved nødvendighetene ved den terapeutiske situasjonen. For eksempel kan de humane antistoffene ifølge oppfinnelsen administreres én eller to ganger ukentlig ved subkutan injeksjon eller én eller to ganger i måneden ved subkutan injeksjon. [0142] Det er spesielt fordelaktig å formulere parenterale sammensetninger i doseringsenhetsform for enkel administrasjon og ensartet dosering. Doseringsenhetsform som anvendt heri refererer til fysisk diskrete enheter egnet som enhetsdoser for subjektene som skal behandles; hver enhet inneholder en forutbestemt mengde aktiv forbindelse beregnet å gi den ønskede terapeutiske effekten i assosiasjon med den krevede farmasøytiske bæreren. Spesifikasjonen for doseringsenhetsformene ifølge oppfinnelsen dikteres av og er direkte avhengig av (a) de unike karakteristikkene for den aktive forbindelsen og den spesielle terapeutiske effekten som skal oppnås, og (b) begrensningene iboende i teknikken med å sette sammen en slik aktiv forbindelse for behandlingen av sensitivitet i individer. 3 [0143] Eksempler på farmasøytisk akseptable antioksidanter inkluderer: (1) vannløselige antioksidanter, så som askorbinsyre, cysteinhydroklorid, natriumbisulfat, natriummetabisulfitt, natriumsulfitt og lignende, (2) oljeløselige antioksidanter, så som askorbylpalmitat, butylert hydroksyanisol (BHA), butylert hydroksytoluen (BHT), lecitin, propylgallat, alfa-tokoferol og lignende, og (3) metall-chelatdannere, så som sitronsyre, etylendiamintetraeddiksyre (EDTA), sorbitol, vinsyre, fosforsyre og lignende. [0144] For de terapeutiske sammensetningene, inkluderer formuleringer ifølge foreliggende oppfinnelse de egnet for oral, nasal, topisk (inkludert bukkal og sublingval), rektal, vaginal og/eller parenteral administrasjon. Formuleringene kan passende presenteres i enhetsdoseform og kan fremstilles ved hvilke som helst metoder kjent
40 39 innen faget farmasi. Mengden aktiv ingrediens som kan kombineres med et bærermateriale for å produsere en enkelt doseringsform vil variere avhengig av subjektet som behandles, og den spesielle administrasjonsmåten. Mengden aktiv ingrediens som kan kombineres med et bærermateriale for å produsere en enkelt doseringsform vil generelt være den mengden av sammensetningen som gir en terapeutisk effekt. Generelt, av ett hundre prosent, vil denne mengden variere fra omkring 0,001 prosent til omkring nitti prosent av aktiv ingrediens, fortrinnsvis fra omkring 0,00 prosent til omkring 70 prosent, mest foretrukket fra omkring 0,01 prosent til omkring prosent. [014] Formuleringer ifølge foreliggende oppfinnelse som er egnet for vaginal administrasjon inkluderer også 1 pessarer, tamponger, kremer, geler, pastaer, skum eller sprayformuleringer som inneholder slike bærere som innen faget er kjent for å være passende. Doseringsformer for den topiske eller transdermale administrasjonen av sammensetninger ifølge denne oppfinnelsen inkluderer pulvere, sprayer, salver, pastaer, kremer, losjoner, geler, løsninger, plaster og inhalasjonsmidler. Den aktive forbindelsen kan bli blandet under sterile betingelser med en farmasøytisk akseptabel bærer, og med hvilke som helst konserveringsmidler, buffere eller drivmidler som kan være krevet. [0146] Uttrykkene "parenteral administrasjon" og "administrert parenteralt" som anvendt heri, betyr administrasjonsmåter andre enn enteral og topisk administrasjon, vanligvis ved injeksjon, og inkluderer, uten begrensning, intravenøs, intramuskulær, intraarteriell, intratekal, intrakapsulær, intraorbital, intrakardial, intradermal, intraperitoneal, transtrakeal, subkutan, subkutikulær, intraartikulær, subkapsulær, subaraknoid, intraspinal, epidural og intrasternal injeksjon og infusjon. [0147] Eksempler på egnede vandige og ikke-vandige bærere som kan anvendes i de farmasøytiske 2 sammensetningene ifølge oppfinnelsen, inkluderer vann, etanol, polyoler (så som glyserol, propylenglykol, polyetylenglykol og lignende), og egnede blandinger derav, vegetabilske oljer, så som f.eks. olivenolje, og injiserbare organiske estere, så som etyloleat. Riktig fluiditet kan opprettholdes for eksempel ved anvendelsen av beleggingsmaterialer, så som lecitin, ved opprettholdelse av den krevede partikkelstørrelsen i tilfellet med dispersjoner og ved anvendelse av overflateaktive midler. [0148] Disse sammensetningene kan også inneholde adjuvanser så som konserveringsmidler, fuktemidler, emulgeringsmidler og dispergeringsmidler. Spesielle eksempler på adjuvanser som er velkjente innen faget inkluderer for eksempel uorganiske adjuvanser (så som aluminiumsalter, f.eks. aluminiumfosfat og aluminiumhydroksid), organiske adjuvanser (f.eks. squalen), olje-baserte adjuvanser, virosomer (f.eks. virosomer som inneholder et membran-bundet heagglutinin og nevraminidase avledet fra influensaviruset). 3 [0149] Forhindring av nærværet av mikroorganismer kan bli sikret både ved steriliseringsprosedyrer, supra, og ved inkluderingen av forskjellige antibakterielle og antifungale midler, for eksempel paraben, klorbutanol, fenol, sorbinsyre og lignende. Det kan også være ønskelig å inkludere isotoniske midler, så som sukkere, natriumklorid og
41 40 lignende i sammensetningene. I tillegg kan langvarig absorpsjon av den injiserbare farmasøytiske formen frembringes ved inkluderingen av midler som forsinker absorpsjon, så som aluminiummonostearat og gelatin. [0] Når forbindelsene ifølge foreliggende oppfinnelse administreres som farmasøytika, til mennesker og dyr, kan de gis alene eller som en farmasøytisk sammensetning som inneholder, for eksempel, 0,001 til 90 % (mer foretrukket 0,00 til 70 %, så som 0,01 til %) aktiv ingrediens i kombinasjon med en farmasøytisk akseptabel bærer. [011] Uavhengig av den valgte administrasjonsruten, blir forbindelsene ifølge foreliggende oppfinnelse, som kan bli anvendt i en egnet hydratisert form, og/eller de farmasøytiske sammensetningene ifølge foreliggende oppfinnelse, formulert til farmasøytisk akseptable doseringsformer ved konvensjonelle metoder kjent for fagpersonene [012] Faktiske doseringsnivåer av de aktive ingrediensene i de farmasøytiske sammensetningene ifølge foreliggende oppfinnelse kan bli variert for å oppnå en mengde av den aktive ingrediensen som er effektiv for å oppnå den ønskede terapeutiske responsen for en spesiell pasient, sammensetning og administreringsmåte, uten være toksisk for pasienten. Det valgte doseringsnivået vil avhenge av en rekke farmakokinetiske faktorer, inkludert aktiviteten av de spesielle anvendte sammensetningene ifølge foreliggende oppfinnelse, eller esteren, saltet eller amidet derav, administrasjonsruten, tidspunktet for administrasjon, raten for utskillelse av den spesielle forbindelsen som anvendes, varigheten av behandlingen, andre legemidler, forbindelser og/eller materialer anvendt i kombinasjon med de spesielle anvendte sammensetningene, alderen, kjønn, vekt, tilstand, generell helsetilstand og tidligere medisinsk historie for pasienten som behandles, og lignende faktorer velkjente i de medisinske fagene. En lege eller veterinær som har vanlig fagkunnskap kan lett bestemme og foreskrive den krevende effektive mengden av den farmasøytiske sammensetningen. For eksempel kunne legen eller veterinæren starte doser av forbindelsene ifølge oppfinnelsen anvendt i den farmasøytiske sammensetningen ved nivåer lavere enn det krevet for å oppnå den ønskede terapeutiske effekten og gradvis øke doseringen inntil den ønskede effekten er oppnådd. Generelt vil en egnet daglig dose av en sammensetning ifølge oppfinnelsen være den mengden av forbindelsen som er den laveste dosen effektiv for å produsere en terapeutisk effekt. En slik effektiv dose vil generelt avhenge av faktorene beskrevet over. Det er foretrukket at administrasjon er intravenøs, intramuskulær, intraperitoneal eller subkutan, fortrinnsvis administrert proksimalt til målsetet. Hvis ønsket kan den effektive daglige dosen av en terapeutisk sammensetning bli administrert som to, tre, fire, fem, seks eller flere underdoser administrert separat ved passende intervaller gjennom hele dagen, eventuelt i enhetsdoseringsformer. Selv om det er mulig for en forbindelse ifølge foreliggende oppfinnelse å bli administrert alene, er det foretrukket å administrere forbindelsen som en farmasøytisk formulering (sammensetning). [013] Terapeutiske sammensetninger kan administreres med medisinske anordninger kjent innen faget. I en foretrukket utførelse, kan for eksempel en terapeutisk sammensetning ifølge oppfinnelsen bli administrert med en nålløs hypodermisk injeksjonsenhet, så som enhetene fremlagt i U.S. Patenter nr.,399,163,,383,81,,312,33,
42 41 1,064,413, 4,941,880, 4,790,824 eller 4,96,6. Eksempler på velkjente implantater og moduler som er nyttige i foreliggende oppfinnelse inkluderer: U.S. Patent nr. 4,487,603, som viser en implanterbar mikro-infusjonspumpe for dispensering av medikamenter ved en kontrollert rate; U.S. Patent nr. 4,486,194, som viser en terapeutisk anordning for administrering av medikamenter gjennom huden; U.S. Patent nr. 4,447,233, som viser en medikamentinfusjonspumpe for å levere medisiner ved en presis infusjonsrate; U.S. Patent nr. 4,447,224, som viser en variabel strøm implanterbar infusjonsapparatur for kontinuerlig legemiddellevering; U.S. Patent nr. 4,439,196, som viser et osmotisk legemiddelleveringssystem som har multi-kammer-kompartimenter, og U.S. Patent nr. 4,47,196, som viser et osmotisk legemiddelleveringssystem. Mange andre slike implantater, leveringssystemer og moduler er kjent for fagpersonene. [014] I visse utførelser kan de monoklonale antistoffene ifølge oppfinnelsen bli formulert for å sikre riktig fordeling in vivo. For eksempel utelukker blod-hjerne-barrieren (BBB) mange høyt hydrofile forbindelser. For å sikre at de terapeutiske forbindelsene ifølge oppfinnelsen krysser BBB-en (om ønskelig), kan de bli formulert, for eksempel, i liposomer. For fremgangsmåter for å fremstille liposomer, se, f.eks. U.S. Patenter 4,22,811;,374,48 og,399,331. Liposomene kan omfatte én eller flere enheter som transporteres selektivt inn i spesielle celler eller organer, og således forbedrer målrettet legemiddellevering av (se f.eks. V.V. Ranade (1989) J. Clin. Pharmacol. 29:68). Eksempelvise målsøkende enheter inkluderer folat eller biotin (se, f.eks. U.S. Patent,416,016 til Low et al.); mannosider (Umezawa et al. (1988) Biochem. Biophys. Res. Commun. 13:38); antistoffer (P.G. Bloeman et al. (199) FEBS Lett. 37:140; M. Owais et al. (199) Antimicrob. Agents Chemother. 39:180); surfaktant protein A- reseptor (Briscoe et al. (199) Am. J. Physiol. 1233:134), forskjellige enheter som kan omfatte formuleringene ifølge oppfinnelsene, så vel som komponenter av molekylene ifølge oppfinnelsen; p1 (Schreier et al. (1994) J. Biol. Chem. 269:9090), se også K. Keinanen; M.L. Laukkanen (1994) FEBS Lett. 346:123; J.J. Killion; I.J. Fidler (1994) Immunomethods 4: V. Fremgangsmåter for anvendelse av antistoffer ifølge oppfinnelsen [01] Beskrevet heri er fremgangsmåter for anvendelse av antistoffer og antigenbindende deler derav som binder ErbB3 i en rekke ex vivo og in vivo diagnostiske og terapeutiske applikasjoner. For eksempel kan antistoffer ifølge oppfinnelsen anvendes for å behandle en sykdom assosiert med ErbB3 avhengig signalisering, inkludert en rekke krefttyper. [016] I én utførelsesform beskriver foreliggende søknad en fremgangsmåte for å behandle en sykdom assosiert 3 med ErbB3 avhengig signalisering ved å administrere til et subjekt et antistoff eller antigenbindende del derav ifølge oppfinnelsen i en mengde som er effektiv for å behandle sykdommen. Egnede sykdommer inkluderer, for eksempel, en rekke krefttyper inkludert, men ikke begrenset til, melanom, brystkreft, eggstokk-kreft, renalt karsinom, gastrointestinal kreft, colonkreft, lungekreft og prostatakreft.
43 42 [017] Antistoffet kan administreres alene eller med et annet terapeutisk middel som virker i forbindelse med eller synergistisk med antistoffet for å behandle sykdommen assosiert med ErbB3 mediert signalisering. Slike terapeutiske midler inkluderer, for eksempel, antikreft-midlene beskrevet infra (f.eks. cytotoksiner, kjemoterapeutiske midler, små molekyler og stråling). [018] I en annen utførelsesform beskriver foreliggende søknad en fremgangsmåte for å diagnostisere en sykdom (f.eks. en kreft) assosiert med ErbB3 oppregulering i et subjekt, ved å bringe antistoffer eller antigenbindende deler ifølge oppfinnelsen (f.eks. ex vivo eller in vivo) i kontakt med celler fra subjektet, og måle nivået av binding til ErbB3 på cellene. Unormalt høye nivåer av binding til ErbB3 indikerer at subjektet har en sykdom assosiert med ErbB3 oppregulering. [019] Også innen omfanget av foreliggende oppfinnelse er kits som omfatter antistoffer og antigenbindende 1 deler derav ifølge oppfinnelsen som eventuelt inkluderer instruksjoner for anvendelse ved behandling eller diagnostisering av en sykdom assosiert med ErbB3 oppregulering og/eller ErbB3 avhengig signalisering. Kitene kan inkludere en etikett som angir den tilsiktede anvendelsen for innholdet i kitet. Begrepet etikett inkluderer ethvert skriv, markedsføringsmateriell eller innspilt materiale levert på eller med kitet, eller som ellers ledsager kitet. [0160] Andre utførelsesformer av foreliggende oppfinnelsen er beskrevet i de følgende eksemplene. [0161] Foreliggende oppfinnelse blir videre illustrert ved de følgende eksemplene. Eksempler 2 Materialer og metoder [0162] Gjennom hele eksemplene, ble de følgende materialer og fremgangsmåter anvendt med mindre noe annet er angitt. 3 [0163] Generelt anvender utførelsen av foreliggende oppfinnelse, med mindre annet er angitt, konvensjonelle teknikker for kjemi, molekylærbiologi, rekombinant DNA-teknologi, immunologi (spesielt, f.eks. antistoffteknologi), og standardteknikker i polypeptidfremstilling. Se, f.eks. Sambrook, Fritsch og Maniatis, Molecular Cloning: Cold Spring Harbor Laboratory Press (1989); Antibody Engineering Protocols (Methods in Molecular Biology),, Paul, S., Humana Pr (1996); Antibody Engineering: A Practical Approach (Practical Approach Series, 169), McCafferty, red, Irl Pr (1996.); Antibodies: A Laboratory Manual, Harlow et al, C.S.H.L. Press, Pub. (1999), og Current Protocols in Molecular Biology, red. Ausubel et al. John Wiley & Sons (1992). In vitro og in vivo modellsystemer for å analysere HCV-biologi, er beskrevet, for eksempel i Cell culture models and animal models of viral hepatitis. Part II: hepatitis C, Lab. Anim. (NY)., 34(2): (0) og i The chimpanzee model of hepatitis C virus infections, ILAR J., 42 (2) : (01).
44 43 Cellelinjer [0164] Alle cellelinjene anvendt i forsøkene beskrevet under ble oppnådd fra the National Cancer Institute eller tilveiebragt ved forskere, som angitt. [016] Cellelinjer: MCF7-ATCC kat. nr. HTB-22 T47D-ATCC kat. nr. HTB-133 Colo37 - Disse cellene ble oppnådd fra en akademisk forsker og er beskrevet av Kolb et al. (06) Int. J. Cancer, 1: Du14 - ATCC kat. nr. HTB-81 OVCAR8 - kilde allerede beskrevet i provisorisk søknad. H197 ATCC kat. nr. CRL Pulverisering av tumorceller [0166] En kryopulveriserer (Covaris Inc) ble anvendt for pulveriseringen av tumorer. Tumorer ble lagret i spesielle poser (forhånds-veid før tilsetningen av tumoren) og plassert i flytende nitrogen og mens en håndterer dem. For små tumorer, ble 0 l lysisbuffer først tilsatt til posen som inneholder tumoren, frosset i flytende nitrogen og deretter pulverisert for å forbedre utvinningen av tumoren fra posen. Pulveriserte tumorer ble overført til 2 ml Eppendorf-rør og plassert i flytende nitrogen inntil klare for videre prosessering Lysis av tumorceller 2 [0167] Tumorer ble lysert i lysisbuffer supplementert med protease- og fosfataseinhibitorer. Lysisbuffer ble tilsatt tumoralikvotene i en sluttkonsentrasjon på omkring 62, mg/ml. Tumorprøver ble homogenisert ved virvling i sek og inkubering på is i omkring min. Lysatene ble spunnet i omkring min i Qiagen Qiashredder kolonner for ytterligere homogenisering av prøvene. Klarede lysater ble alikvotert til nye rør for videre behandling. BCA Analyse 3 [0168] BCA-analysen (Pierce) ble utført ved å følge produsentens protokoll på alle tumorprøver. Den totale proteinkonsentrasjonen (i mg/ml) av hver tumorprøve ble senere anvendt for normaliseringen av ELISA resultatene.
45 44 ELISA-analyse [0169] Alle ELISA reagenser for den totale og fosfo-erbb3 ELISAene ble innkjøpt fra R&D Systems som DuoSet kits brønn Nunc Maxisorb plater ble belagt med 0 ul av et antistoff og inkubert over natten ved romtemperatur. Neste morgen ble platene vasket 3 ganger med 00 l/brønn i BioTek platevaskeren med PBST (0,0 % Tween- ). Platene ble deretter blokkert i omkring en 1 time ved romtemperatur med 2 % BSA i PBS. Platene ble vasket 3 ganger med 00 l/brønn i BioTek platevaskeren med PBST (0,0 % Tween-). 0 l cellelysater og standarder fortynnet i 0 % lysisbuffer og 1 % BSA ble anvendt i duplikater for videre prosessering. Prøver ble inkubert i 2 timer ved 4 C på en platerister og vasket 3 ganger med 00 l/brønn i BioTek platevaskeren med PBST (0,0 % Tween-). Omkring 0 l av et deteksjons-antistoff fortynnet i 2 % BSA, PBST ble tilsatt og inkubert i omkring 1 time ved romtemperatur. For fosfor-erbb3, ble deteksjons-antistoffet direkte konjugert til pepperrotperoksidase (HRP) og inkubert i 2 timer ved romtemperatur. Platen ble vasket 3 ganger med 00 l/brønn i BioTek platevaskeren med PBST (0,0 % Tween-). Omkring 0 l streptavidin-hrp ble tilsatt, og inkubert i min ved romtemperatur (med unntak av perbb3). Platene ble vasket 3 ganger med 00 l/brønn i BioTek platevaskeren med PBST (0,0 % Tween-). Omkring 0 l Supersignal Pico ELISA substrat ble tilsatt og platen ble avlest ved anvendelse av en Fusion plateavleser. Dataene ble analysert ved anvendelse av EXCEL. Det ble beregnet gjennomsnitt av duplikate prøver og feilstolpene ble anvendt for å representere standardavviket mellom de to replikatene. Eksempel 1: Produksjon av antistoffer ved anvendelse av fag-display [0170] For å oppnå humane anti-erbb3 antistoffer referert til heri som Ab #6, Ab #3, Ab #14, Ab #17 og Ab #19, 2 ble et humant Fab-fag bibliotek som inkluderer en unik kombinasjon av immunoglobulinsekvenser oppnådd fra humane donorer (Hoet et al. supra) innledende screenet for ErbB3 bindere. [0171] Ved anvendelse av renset ErbB3 og en kinesisk hamster eggstokk-cellelinje som uttrykker celleoverflate ErbB3, ble 73 unike Fab-sekvenser fra biblioteket identifisert. Disse 73 klonene ble så reformatert som Fab bare uten fagen. Ved anvendelse av høy-gjennomstrømmingsmetoder, ble disse Fab-ene uttrykt i liten skala og testet for binding ved anvendelse av ELISA og Flexchip metoden som er en høy-gjennomstrømningsoverflateplasmonresonans (SPR) teknologi. De 73 Fabene uten fag ble satt som flekker på en chipoverflate og bindingskinetikken og epitopeblokkeringen til et ErbB3-his fusjon målprotein eller et ErbB3-Fcprotein (R&D Systems) ble målt. Likevekt bindingskonstanten og "on/off" rater for Fab-ene ble beregnet fra de oppnådde dataene. 3 [0172] Binding av de ulike Fab-ene til MALME-3M celler var det neste som ble undersøkt ved anvendelse av omkring 00 nm av Fab-ene og en 1:70 fortynning av et geite-anti-menneske Alexa 647 sekundært antistoff. Som vist i figurene 1A og 1B, utviste flere kandidat Fab-er betydelig farging av MALME-3M celler.
46 4 Eksempel 2: Optimalisering av anti-erbb3 Fab-er [0173] Etter identifiseringen av Fab-er som blokkerte bindingen av ErbB3-ligand, heregulin, til ErbB3, ble VH- og VL-sekvensene av Fab-ene kodonoptimalisert som følger. [0174] Spesielt ble VH og VL-regionene reformatert ved anvendelse av ekspresjonskonstrukter for ekspresjon som en IgG1 eller IgG2 isotype. Konstruktene inkluderte en Selexis ryggrad som har en kassett designet for substitusjon av de passende tung- og lett-kjede sekvensene. Selexis vektorene inkluderte en CMV promoter og et samsvarende poly-a signal. [017] Nukleinsyresekvensene for de kodonoptimaliserte VH og VL av Ab #6 er fremlagt i henholdsvis SEKV ID NR: 2 og 26, og de for Ab #3 er fremlagt i henholdsvis SEKV ID NR: 27 og 28, som vist i figur Eksempel 3: Bindingsaffinitet for ErbB3 [0176] Dissosiasjonskonstantene av anti-erbb3-antistoffene ble målt ved anvendelse av to uavhengige teknikker dvs. en overflate-plasmonresonans-analyse og en celle-bindingsanalyse ved anvendelse av MALME-3M celler. Overflate-plasmonresonans-analyse [0177] Overflate-plasmonresonans-analysen (også referert til som Flexchip-analysen) ble utført som beskrevet i Wassaf et al. (06) Analytical Biochem., 31: K D -verdien ble beregnet basert på formelen K D = K d /K a. 2 [0178] K D -verdiene av henholdsvis Ab #6 og Ab #3, som målt ved anvendelse av overflate-plasmonresonansanalysen, er avbildet i figurene 2A og 2B. Ab #6 hadde en K D -verdi på omkring 4 nm og Ab #3 hadde en K D -verdi på omkring 8 nm, som vist i henholdsvis figurene 2A og 2B. Cellebindingsforsøk [0179] Cellebindingsanalysen for å bestemme K D -verdiene av Ab #6 og Ab # 3 ble utført som følger. 3 [0180] MALME-3M celler ble løsnet med 2 ml trypsin-edta + 2 ml RMPI + mm EDTA ved romtemperatur i minutter. Komplett RPMI ( ml) ble tilsatt umiddelbart til de trypsiniserte cellene, resuspendert forsiktig og sentrifugert i en Beckman bordsentrifuge ved 10 rpm i minutter. Celler ble resuspendert i BD fargebuffer (PBS + 2 % føtalt bovint serum + 0,1 % natriumazid, Becton Dickinson) ved en konsentrasjon på 2 x 6 celler pr ml og 0 l (1 x celler) alikvoter ble plettert i en 96-brønn titerplate. [0181] En l løsning av 0 nm anti-erbb3 antistoff (Ab #6 eller Ab #3) i BD fargebuffer ble fremstilt i et Eppendorf-rør og seriefortynnet 2-ganger i 7 l BD fargebuffer. Konsentrasjonene av det fortynnede antistoffet spente fra 0 nm til 0,4 nm. 0 l alikvoter av de forskjellige proteinfortynningene ble så tilsatt direkte til 0 l
47 46 cellesuspensjonen for å gi sluttkonsentrasjonene på 0 nm, 0 nm, 2 nm, 12 nm, 6 nm, 3 nm, 1, nm, 0,8 nm, 0,4 nm og 0,2 nm av antistoffet. [0182] Alikvoterte celler i 96-brønnplaten ble inkubert med proteinfortynninger i minutter ved romtemperatur på en plattformrister og vasket 3 ganger med 0 l BD fargebuffer. Celler ble så inkubert med 0 l av en 1:70 fortynning av Alexa 647-merket geite-anti-menneske IgG i BD fargebuffer i 4 minutter på en plattformrister i kjølerommet. Til slutt ble cellene vasket to ganger, pelletisert og resuspendert i l BD fargebuffer +0, g/ml propidiumjodid. Analyse av 000 celler ble gjort i et FACScalibur strømningscytometer ved anvendelse av FL4 kanalen. MFI-verdier og de tilsvarende konsentrasjonene av anti-erbb3-antistoffene ble plottet på henholdsvis y- aksen og x-aksen. K D for molekylet ble bestemt ved GraphPad Prism ved anvendelse av ett-sete bindingsmodellen for en ikke-lineær regresjonskurve. [0183] K D -verdien ble beregnet basert på formelen Y = Bmax * X/K D + X (Bmax = fluorescens ved metning. X = 1 antistoffkonsentrasjon. Y = grad av binding). Som vist i figurene 2C og 2D, hadde Ab #6 og Ab #3 K D -verdier på henholdsvis omkring 4 nm og 1,3 nm, i en celle-bindingsanalyse ved anvendelse av MALME-3M celler. Eksempel 4: Bindingsspesifisitet/Epitopebinding for ErbB3 [0184] Bindingsspesifisiteten av en IgG2-isotype av Ab #6 til ErbB3 ble analysert ved anvendelse av ELISA som følger. Identifikasjon av epitopen bundet av Ab #6 ble også analysert. [018] Spesifikt ble 96-brønn Nunc Maxisorb plater belagt med 0 l av g/ml protein (rekombinant human ErbB3, rekombinant human EGFR eller urelatert protein (BSA)) og inkubert over natten ved romtemperatur. Neste 2 morgen ble platene vasket 3 ganger med 00 l/brønn PBST (0,0 % Tween-) i BioTek platevaskeren. Brønnene ble blokkert i 1 time ved romtemperatur med 2 % BSA i PBS. Platene ble vasket 3 ganger med 00 l/brønn av PBST (0,0 % Tween-) i BioTek platevaskeren. Omkring 0 L av Ab #6 ble tilsatt ved flere fortynninger (1 M og serielle to-gangers fortynninger) i 2 % BSA, PBST. Alle prøvene ble kjørt i duplikat og inkubert i 2 timer ved 4 C på en platerister. Platene ble vasket plater 3 ganger med 00 l/brønn PBST (0,0 % Tween-) i BioTek platevaskeren. 0 l humant IgG deteksjonsantistoff (HRP-konjugert (Bethyl Inc, 1:7000 fortynning i 2 % BSA, PBST)) ble tilsatt og platene ble inkubert i 1 time ved romtemperatur. Platene ble vasket plate 3 ganger med 00 l/brønn PBST (0,0 % Tween-) i BioTek platevaskeren. 0 l Supersignal Pico ELISA substrat ble tilsatt og platen ble avlest på Fusion plateavleseren. Dataene ble analysert ved anvendelse av EXCEL-programmet. Det ble beregnet gjennomsnitt av duplikate prøver og feilstolpene representerer standardavviket mellom de to replikatene. 3 [0186] Som vist i figur 3, bandt Ab #6 rekombinant ErbB3 i en ELISA, men viste ingen vesentlig binding til EGFR, BSA eller TGF-α.
48 47 [0187] Et fragment (trunkeringsmutant) som tilsvarer aminosyrerester -2 av ErbB3 ble klonet inn i gjærdisplay-vektoren pyd2 (en modifisert versjon av pyd1 (Invitrogen) med et stoppkodon konstruert foran Histaggen) mellom Nhe og BsiWI restriksjonssetene. Plasmidet ble transformert inn i gjærstammen EBY0 (Invitrogen), og kloner som inneholder plasmidet valgt på Trp-selektivt medium. Denne klonen ble dyrket i glukoseholdig medium over natten ved C og ekspresjon av ErbB3 trunkeringsmutanten ble indusert ved overføring til et galaktose-holdig medium i 2 dager ved 18 C. Gjær som viser ErbB3 trunkeringsmutanten ble farget med 0 nm Ab #6, fulgt av et geite-anti-menneske antistoff merket med Alexa farge-647. En separat prøve ble farget med geite-anti-menneske antistoffet bare for å vise at det ikke er noen ikke-spesifikk binding til gjær av det sekundære antistoffet. Analyse ble utført ved strømningscytometri på FACS Calibur cellesortereren (BD Biosciences). [0188] Som vist i figur, bandt Ab #6 til trunkeringsmutanten, dvs. aminosyrerester -2 av ErbB3. Eksempel : Nedregulering av total ErbB3 på tumorceller 1 [0189] Evnen Ab #6 har til å nedregulere ErbB3 ekspresjon både in vitro og in vivo i tumorceller ble testet som følger. 2 [0190] MALME-3M-celler ble sådd i 96 brønn vevskulturplater og dyrket i RPMI-1640 medium supplementert med antibiotika, 2 mm L-glutamin og % føtalt bovint serum (FBS) i 24 timer ved 37 C og % karbondioksid. Media ble deretter skiftet til RPMI-1640 MEDIA med antibiotika, 2 mm L-glutamin med og uten antistoffet ved konsentrasjoner på 1 um, nm, 63 nm, 16 nm, 4,0 nm, 1,0 nm, 240 pm, 61 pm og 1 pm. Celler ble dyrket i 24 timer ved 37 C og % karbondioksid, vasket med kald PBS, så høstet med mammalsk proteinekstrakt (MPER) lysis (Pierce, 780) buffer inneholdende mm NaCl, mm natriumpyrofosfat, um bpv (phen), 0 um fenalarsin, 1 mm natrium ortovanadat, og proteaseinhibitor cocktail (Sigma, P714). Cellelysater ble fortynnet to-ganger med 4 % bovint serumalbumin i fosfatbufret saltløsning med 0,1 % tween-, så analysert ved ELISA med muse-antimenneske ErbB3 captureantistoff og biotinylert muse-anti-menneske ErbB3 sekundært deteksjonsantistoff. Signal ble generert med streptavidin konjugert til pepperrotperoksidase reagert med kjemiluminescerende substrat (Pierce, 37070). ELISA-er ble visualisert ved anvendelse av et luminometer. [0191] Som vist i figur 4, reduserte Ab #6 totale ErbB3-nivåer med omkring 46,9 % i MALME-3M celler in vitro, som målt ved ELISA. Medier som ikke inneholder serum og antistoff ble anvendt som kontroll. 3 [0192] I et ytterligere forsøk ble nedreguleringen av ErbB3-reseptorer på MALME-3M celler ved anvendelse av IgG1-og IgG2-isotyper av Ab #6 undersøkt ved anvendelse av FACS-analyse. MALME-3M celler ble trypsinisert fra en 1 cm plate, og vasket én gang med RPMI + % føtalt bovint serum. Cellepellets ble resuspendert ved en tetthet på 1 x 6 celler per ml. To alikvoter på 2 x celler ble tilsatt til en 12-brønn vevskulturplate, og resuspendert i et sluttvolum på 800 ul RPMI + % føtalt bovint serum. Til én brønn ble Ab #6 IgG1 eller Ab-
49 48 #6IgG2 isotypen tilsatt til en sluttkonsentrasjon på 0 nm (behandlet prøve) og til den andre brønnen, ble et ekvivalent volum av PBS (ubehandlet prøve) tilsatt. [0193] Den påfølgende dagen, ble behandlede og ubehandlede celler trypsinisert, vasket og inkubert med 0 nm Ab #6 i BD fargebuffer i minutter på is. Celler ble vasket to ganger med 1 ml BD fargebuffer og inkubert med 0 ul av en 1:00 fortynning av Alexa 647-merket geite-anti-menneske Alexa 647 i 4 minutter på is. Celler ble så vasket og resuspendert i 0 ul BD fargebuffer + 0, ug/ml propidiumjodid. Analyse av.000 celler ble gjort i et FACScalibur strømningscytometer ved anvendelse av FL4 kanalen. [0194] Som vist i figurene A og B nedregulerte både IgG1 og IgG2 isotyper av Ab #6 ErbB3 på MALME-3M celler med henholdsvis omkring 62 % og omkring 66 %. 1 2 [019] For å bestemme om denne reduksjonen skyldtes internalisering av ErbB3 reseptoren på overflaten av MALME-3M-celler, ble det målt ekspresjon av ErbB3 i nærværet av antistoffet over tid. Spesifikt ble MALME-3M celler trypsinisert fra en 1 cm plate, og vasket én gang med RPMI + % føtalt bovint serum. Cellepellets ble resuspendert ved en tetthet på 1 x 6 celler per ml. To alikvoter på 2 x celler ble tilsatt til en 12-brønn vevskulturplate, og resuspendert i et sluttvolum på 800 l RPMI + % føtalt bovint serum. Til én brønn, ble anti- ErbB3-antistoffet tilsatt til en sluttkonsentrasjon på 0 nm (behandlet prøve) og til den andre brønnen, ble et ekvivalent volum av PBS (ubehandlet prøve) tilsatt. Den påfølgende dagen, ble behandlede og ubehandlede celler trypsinisert, vasket og inkubert med 0 nm anti-erbb3-antistoff i BD fargebuffer i minutter på is. Celler ble vasket to ganger med 1 ml BD fargebuffer og inkubert med 0 l av en 1:00 fortynning av Alexa 647-merket geite-anti-menneske Alexa 647 i 4 minutter på is. Celler ble så vasket og resuspendert i 0 l BD fargebuffer + 0, g/ml propidiumjodid. Analyse av.000 celler ble gjort i et FACScalibur strømningscytometer ved anvendelse av FL4 kanalen. [0196] Som vist i figur 6, ble nedregulering av ErbB3 i nærværet av Ab #6 målt ved 0 time (figur 6A), 0, timer (figur 6B), 2 timer (figur 6C) og 24 timer (figur 6D). Som vist i figur 6A-6D, var omkring 0 % av celleoverflate ErbB3- reseptorene nedregulert etter omkring minutter og ved omtrent 24 timer, var omkring 93 % av celleoverflatereseptorene nedregulert. [0197] Evnen Ab #6 har til å forårsake ErbB3 nedregulering in vivo i melanom-celler ble også undersøkt som følger. [0198] Kort fortalt ble T-celle defisiente nu/nu mus (3-4 uker gamle hunnmus med opprinnelse ved NIH, utavlet; 3 albinobakgrunn) innkjøpt fra Charles River Labs (Wilmington, MA). MALME-3M celler for implantasjon ble dyrket i kultur (RPMI medium, % FBS, L-glutamin og antibiotika, 37 C, % CO2) til omkring 80 % konfluens før høsting. Celler ble holdt på is inntil implantasjon. Mus ble implantert via subkutan injeksjon med 0 ul MALME-3M celler på den høyre flanken og tillatt å restituere mens de blir overvåket for innledende tumorvekst.
50 49 [0199] Tumorene ble målt (lengde ganger bredde) med digitalt skyvelære og musene ble dosert med IgG2a (Sigma, M7769-MG) ved intravenøs injeksjon. Mus ble dosert intra-peritonealt annenhver dag med enten 1 g eller 0 g av antistoff nummer seks og tumorer ble målt tre ganger per uke og registrert i et Microsoft EXCELregneark. [00] Slutt-tumormålinger (L x W) ble tatt, musene ble avlivet ved CO 2 -kvelning og tumorer ble skåret ut, hurtigfrosset i flytende nitrogen, og ble lagret ved -80 C (for biokjemisk analyse). Slutt-tumormålinger ble analysert og plottet ved tumorareal og tumorvolum som beskrevet, for eksempel i Burtrum et al. (03) Cancer Res., 63: Dataene ble også analysert ved "normaliserte" og "ikke-normaliserte" metoder for både tumorvolum og tumorareal. For "normaliseringen" av dataene, ved hvert tidspunkt for målingen, ble hver tumor i hver gruppe dividert med den innledende tumorstørrelsen bestemt ved skyvelæremåling. [01] Som vist i figur 7, blant de ulike antistoffene testet i denne undersøkelsen, forårsaket Ab #6 nedregulering 1 av total ErbB3 så tidlig som 24 timer etter injeksjon i tumorer behandlet med enten IgG1 eller IgG2-isotype av Ab #6). PBS ble anvendt som en kontroll [02] I et ytterligere forsøk, ble evnen Ab #6 har til å nedregulere ErbB3 i ADRr xenografter in vivo undersøkt. 2 [03] Kort fortalt ble prøvene pulverisert i en kryopulveriserer (Covaris Inc). Tumorer ble lagret i spesielle poser (forhånds-veid før tilsetningen av tumoren) og plassert i flytende nitrogen mens en håndterer dem. For små tumorer, ble 0 l lysisbuffer først tilsatt til posen med tumoren, frosset i flytende nitrogen og så pulverisert for å forbedre utvinningen av tumoren fra posen. Pulveriserte tumorer ble overført til 2 ml Eppendorf-rør og plassert i flytende nitrogen inntil lysert. Tumorer ble lysert i lysisbuffer supplementert med protease- og fosfataseinhibitorer. Lysisbuffer ble tilsatt til tumoralikvotene i en sluttkonsentrasjon på 62, mg/ml. Tumorprøver ble homogenisert ved virvling i sekunder, og la dem sitte på is i min. Lysatene ble spunnet i minutter i Qiagen Qiashredder kolonner for ytterligere homogenisering av prøvene. Klarede lysater ble alikvotert til nye rør. [04] BCA-analysen ble utført som fremlagt i avsnittet Materialer og metoder supra. 3 [0] De totale nivåene av ErbB3 ble bestemt ved ELISA. ELISA reagensene ble innkjøpt fra R&D Systems som DuoSet kits. 96-brønn Nunc Maxisorb plater ble belagt med 0 l av respektivt capture-antistoff og inkubert over natten ved romtemperatur. Den neste morgenen ble platene vasket 3 ganger med 00 l/brønn i en BioTek platevasker med PBST (0,0 % Tween-) og så blokkert i 1 time ved romtemperatur med 2 % BSA i PBS. Platene ble så vasket tre ganger med 00 l/brønn i BioTek platevaskeren med PBST (0,0 % Tween-). Lysater (0 l) og standarder ble fortynnet i 0 % lysisbuffer og 1 % BSA; alle prøver ble kjørt i duplikat. Plater ble inkubert i 2 timer ved 4 C på en platerister og så vasket tre ganger med 00 l/brønn i en BioTek platevasker med PBST (0,0 % Tween-). Femti mikroliter deteksjonsantistoff fortynnet i 2 % BSA, PBST ble tilsatt og platene ble inkubert i 1 time ved romtemperatur. Plater ble vasket tre ganger med 00 l/brønn i BioTek platevaskeren med PBST (0,0
51 0 % Tween-). Femti mikroliter streptavidin-hrp ble tilsatt og platene ble inkubert i minutter ved romtemperatur. Platene ble igjen vasket tre ganger med 00 l/brønn i en BioTek platevasker med PBST (0,0 % Tween-). Femti mikroliter Supersignal Pico ELISA substrat ble tilsatt og avlesning ble utført på en Fusion plateavleser. Data ble analysert ved anvendelse av EXCEL. Det ble beregnet gjennomsnitt av duplikate prøver og feilstolpene representerer standardavviket mellom de to gjentak. [06] Resultatene av dette forsøket er vist i figur 8. Som vist i Figur 8, nedregulerte Ab #6 ErbB3 i ADRr xenografter in vivo. Eksempel 6: Inhibering av tumorcelleproliferasjon [07] Evnen Ab #6 har til å inhibere cellulær proliferasjon av celler som uttrykker ErbB3 (f.eks. kreftceller), ble undersøkt som følger. 1 [08] MALME3M, ACHN og NCI/ADRr-celler ble sådd i 96 brønn vevskulturplater og dyrket i RPMI-1640 medium supplementert med antibiotika, 2 mm L-glutamin og % føtalt bovint serum (FBS) i 24 timer ved 37 grader Celsius og % karbondioksid. Media ble så skiftet til RPMI-1640 medium med antibiotika, 2 mm L-glutamin og med og uten antistoffet ved 1 um, nm, 63 nm, 16 nm, 4,0 nm, 1,0 nm, 240 pm, 61 pm og 1 pm konsentrasjoner. Celler ble dyrket i 96 timer ved 37 C og % karbondioksid, så høstet med CellTiter-Glo Luminescent cellelevedyktighetsanalyse (Promega, G773) og analysert på et luminometer. Medier som ikke inneholder serum og antistoff ble anvendt som kontroll. 2 [09] Som vist i figurene 9, og 11, inhiberte Ab #6 proliferasjon av MALME-3M celler (figur 9), ADRr eggstokkkreftceller (figur ) og ACHN-celler (figur 11) som uttrykker ErbB3. Spesifikt inhiberte Ab #6 proliferasjon av MALME-3M celler med omkring 19,6 %, målt ved anvendelse av celletiter glød-analysen, og inhiberte proliferasjon av ADRr eggstokk-kreftceller med omkring, %. Også, som vist i Figur 11, inhiberte Ab #6 proliferasjon av ACHNceller ved omtrent 2,4 %. Eksempel 7: Inhibering av ErbB3-fosforylering i tumorceller [02] Evnen Ab #6 har til å inhibere ErbB3-fosforylering in vivo ble undersøkt som følger. 3 [0211] Prøvene ble pulverisert ved anvendelse av teknikken beskrevet i eksempel supra, med hensyn til figur 8. BCA-analysen ble utført som fremlagt i Materialer og metoder seksjonen supra, og ELISA-analysen ble utført som beskrevet i eksempel supra med hensyn til figur 8.
52 1 [0212] Resultatene av dette forsøk er vist i figur 12. Som vist i Figur 12, inhiberte Ab #6 signifikant ErbB3- fosforylering i ADRr eggstokk-xenografter in vivo, som målt ved mengden fosforylert ErbB3 (perbb3) i ng/mg totalt protein. [0213] Evnen Ab #6 har til å inhibere betacellulin (BTC) eller heregulin (HRG) indusert ErbB3-fosforylering ble også undersøkt, som følger. [0214] Eggstokk-ADRr-celler ble preinkubert med Ab #6 i minutter før stimulering med 0 mm BTC, mm HRG eller 333 nm TGF-α. Etter preinkubering ble mediene fjernet og cellene ble stimulert i minutter ved 37 C, % CO2 med 0 nm BTC eller 333 nm TGF-α (for PE498). HRG kontroller ( minutter, nm), % serum og 0 % serum kontroller ble også anvendt. Celler ble vasket med 1X kald PBS og lysert i μl kald lysisbuffer (M-PERbuffer pluss natrium-vanadat (NaVO4, Sigma), 2-glyserofosfat, fenylarsinoksid, BpV og proteaseinhibitorer) ved inkubering på is i minutter. Lysater ble lagret over natten ved -80 C. 1 [021] Som vist i figurene 13A-13C, inhiberte Ab #6 signifikant både betacellulin- og heregulin-mediert fosforylering av ErbB3. [0216] I et ytterligere eksperiment, ble evnen Ab #6 har til å inhibere ErbB3-fosforylering i ovarie tumorcellelinjer OVCAR og OVCAR 8 undersøkt som følger. [0217] OVCAR og OVCAR 8 cellelinjene ble oppnådd fra National Cancer Institute, Division of Cancer Treatment and Diagnostics ("DCTD"). ELISAen ble utført som beskrevet i Materialer og metoder seksjonen supra. 2 [0218] Resultatene av dette forsøket er avbildet i figurene 14A og 14B. Som beskrevet i figurene 14A og 14B, inhiberte Ab #6 ErbB3-fosforylering i både OVCAR og OVCAR 8 eggstokk-kreftcellelinjer. [0219] Som diskutert over, inhiberer Ab #6 betacellulin-mediert fosforylering av ErbB3. For å undersøke om 3 betacellulin-mediert fosforylering av ErbB3 forekommer gjennom ErbB1 eller ErbB3, ble følgende eksperiment utført. [02] ADRr-celler eller MALME-3M celler (1 x ) ble pre-inkubert med 2 μm anti-erbb3 Ab #6 eller 2 M Erbitux (som kontroll) i 0 l BD fargebuffer i minutter på is. Etter minutter ble 0 l 400 nm biotinylert BTC tilsatt til cellene og inkubert i ytterligere minutter på is. Dette ga en sluttkonsentrasjon på 12, M antistoffer og 0 nm BTC. Celler ble så vasket to ganger med 00 l BD fargebuffer og inkubert med 0 l av en 1:0 fortynning av streptavidin-pe (PE = fycoerytrin) (Invitrogen) i BD fargebuffer i 4 minutter. Til slutt ble cellene vasket to ganger, resuspendert i 0 l BD fargebuffer og analysert i et FACScalibur strømningscytometer. Som en positiv kontroll, ble 1 x ADRr eller MALME-3M-celler inkubert med 0 nm BTC i minutter på is, vasket to ganger og inkubert med en 1:0 fortynning av streptavidin-pe i 4 minutter. For å vurdere bakgrunnsfarging fra
53 2 streptavidin-pe-konjugatet ble celler inkubert med 0 l av en 1:0 fortynning av streptavidin-pe bare i 4 minutter. [0221] Resultatene av dette forsøket er avbildet i figurene 1A-1C. Som vist i figur 1A, viser betacellulin (BTC) ikke noen vesentlig binding til ErbB1 negative MALME-3M celler. Som vist i figurene 1B og 1C, viser imidlertid BTC-binding til ErbB1-positive ADRr-celler. [0222] Også, som vist i figurene 1B og 1C, ble denne bindingen blokkert av erbitux, som er et anti-egfrantistoff som spesifikt binder EGFR og ble inkludert som en kontroll for å vise at EGF-lignende ligander binder til EGFR, og som er beskrevet i f.eks. Adams et al. (0), Nature Biotechnology 23, Eksempel 8: Inhibering av heregulin-mediert signalisering i tumorceller 1 2 [0223] Evnen Ab #6 har til å inhibere heregulin-mediert tumorcellesignalisering ble undersøkt som følger. [0224] MALME-3M-celler ble sådd i 96 brønn vevskulturplater og dyrket i RPMI-1640 medium supplementert med antibiotika, 2 mm L-glutamin og % føtalt bovint serum (FBS) i 24 timer ved 37 C og % karbondioksid. Cellene ble serumsultet i RPMI-1640 medium med antibiotika og 2 mm L-glutamin i 24 timer ved 37 C og % karbondioksid. Celler ble forbehandlet med og uten anti-erbb3 antistoff (IgG2 isotype av Ab #6) ved 1 M, nm, 63 nm, 16 nm, 4,0 nm, 1,0 nm, 240 pm og 61 pm konsentrasjoner i minutter og deretter stimulert med HRG1- beta 1-ECD i minutter ved 37 C og % karbondioksid. Celler ble vasket med kald PBS så høstet med mammalsk proteinekstrakt (MPER) lysis (Pierce, 780) buffer som inneholder mm NaCl mm natriumpyrofosfat, um bpv (phen), 0 μm fenalarsin, 1 mm natrium ortovanadat, og proteaseinhibitor cocktail (Sigma, P714 ). Cellelysater ble fortynnet to ganger med 4 % bovint serumalbumin i fosfatbufret saltløsning med 0,1 % Tween-, så analysert ved ELISA for AKT (en nedstrømseffektor av ErbB3) og ErbB3-fosforylering. [022] For å teste for AKT-fosforylering, ble lysater kjørt på en ELISA-plate med et capture-antistoff som er spesifikt for AKT og biotinylert deteksjonsantistoff spesifikt for fosforyleringssetet på serin 473 av AKT. Signal ble generert med streptavidin konjugert til pepperrotperoksidase reagert med kjemiluminescerende substrat (Pierce, 37070). For å analysere for ErbB3-fosforylering, ble lysater kjørt på en ELISA-plate med et capture-antistoff spesifikt for ErbB3 og et anti-fosfotyrosin deteksjonsantistoff konjugert til pepperrotperoksidase. Dette ble så reagert med kjemiluminescerende substrat (Pierce, 37070). ELISAer ble visualisert ved anvendelse av et luminometer. 3 [0226] Som vist i figurene 16A og 16B, var Ab #6 en potent inhibitor av heregulin-mediert signalisering i MALME- 3M-celler, som målt ved redusert fosforylering av ErbB3 (figur 16A) og AKT (figur 16B). Påfallende inhiberte Ab #6 fosforyleringen av AKT med nesten 0 %.
54 3 Eksempel 9: Inhibering av eggstokk-, prostata- og bukspyttkjertel-tumorvekst [0227] For å vurdere virksomheten av Ab #6 in vivo, ble flere xenograftmodeller av human kreft etablert i nakenmus og inhiberingen av tumorvekst ble vurdert ved multiple doser. For eksempel, ble T-celle defisiente nu/nu mus (3-4 uker gamle hunnmus med opprinnelse ved NIH, utavlet; albino bakgrunn) innkjøpt fra Charles River Labs (Wilmington, MA) for xenograftstudier. ADRr-celler for implantasjon ble dyrket i kultur (RPMI medium, % FBS, L-glutamin og antibiotika, 37 C, % CO2) til omkring 8 % konfluens før høsting. Celler ble holdt på is inntil implantasjon. Mus ble implantert via subkutan injeksjon med 0 l ADRr-celler på den høyre flanken og tillatt å restituere mens de blir overvåket for innledende tumorvekst. [0228] Tumorer ble målt (lengde ganger bredde) med digitalt skyvelære og musene ble dosert med IgG2a (Sigma, M7769-MG) ved intravenøs injeksjon. Mus ble dosert intra-peritonealt hver tredje dag med enten g eller 0 g Ab #6 og tumorer ble målt tre ganger per uke og registrert i et Microsoft EXCEL-regneark. 1 [0229] Slutt-tumor-målinger (L x W) ble tatt, ble musene avlivet ved CO2-kvelning og tumorer ble skåret ut, hurtigfrosset i flytende nitrogen og ble lagret ved -80 C (for biokjemisk analyse). Slutt-tumormålinger ble analysert og plottet ved tumorareal og tumorvolum, som beskrevet i Burtrum et al. supra. Dataene ble også analysert ved "normaliserte" og "ikke-normaliserte" metoder for både tumorvolum og tumorareal. For "normaliseringen" av dataene, ved hvert tidspunkt av målingen, ble hver tumor i hver gruppe dividert med den innledende tumorstørrelsen bestemt ved skyvelæremåling. [02] Dataene fra tre forskjellige modeller avledet fra humane tumorcellelinjer, ADRr (eggstokk-), Du14 2 (prostata) og OvCAR8 (eggstokk) er vist i figurene 17A-C og Colo37 xenograftstudie er vist i figur 17D. Dataene fra disse studiene viste at en 0 μg dose av Ab #6 hver tredje dag (Q3d) resulterer i betydelig inhibering av tumorvekst (p<0,0 for flere tidspunkter i løpet av studiene). Denne inhiberende effekten av Ab #6 ble dessuten ytterligere hevet når dosen ble økt til 600 μg, Q3d, i Du14 prostatakreftmodellen så vel som en renal og en pankreatisk karsinom xenograftmodell (ACHN og COLO37). Ytterligere heving av dosen til 0 g Q3d resulterte imidlertid ikke i økt virksomhet (OvCAR8- Figur 17; COLO37) noe som foreslår at 600 μg er metning med hensyn til tumorvekstinhibering. Farmakokinetiske (PK) analyser av serumet fra dyrene fra disse studiene viser en doseavhengig økning i serumretensjonen av Ab #6. Likeledes viste biokjemisk analyse av de intra-tumorale nivåene av Ab #6 fra disse forskjellige studiene et doseavhengig område på 0 til ~ 6 g MM121/μg totalt tumorlysat (data ikke vist). 3 Eksempel : Inhibering av binding av ErbB3-ligander til ErbB3 på kreftceller [0231] I et ytterligere eksperiment ble spesifisiteten for antistoffene ifølge foreliggende oppfinnelse til å inhibere bindingen av ErbB3-ligander til ErbB3, og ikke EGF-lignende ligander til EGFR, undersøkt som følger.
55 4 [0232] I ett eksperiment ble spesifisiteten av Ab #6 og en Fab-versjon av Ab #3 (Ab/Fab #3) til å inhibere bindingen av ErbB3-ligander (f.eks. heregulin og epiregulin) til ErbB3 undersøkt. [0233] For å undersøke evnen Ab #6 og Ab/Fab #3 har til å inhibere bindingen av heregulin til ErbB3, ble følgende eksperiment utført. [0234] ADRr-celler (1 x ) ble inkubert med M av et anti-erbb3 antistoff (f.eks. Ab #6 eller Ab/Fab #3) i 0 l BD fargebuffer i minutter på is. Etter minutter ble 0 l av 40 nm biotinylert heregulin EGF tilsatt til cellene 1 og inkubert i ytterligere minutter på is. Dette ga en endelig konsentrasjon på M antistoff og nm heregulin EGF. Cellene ble så vasket to ganger med 00 l BD fargebuffer og inkubert med 0 l av en 1:0 fortynning av streptavidin-pe (PE = fycoerytrin) (Invitrogen) i BD fargebuffer i 4 minutter. Til slutt ble celler vasket to ganger, resuspendert i 0 l BD fargebuffer og analysert i et FACScalibur strømningscytometer. Som en positiv kontroll ble 1 x ADRr-celler inkubert med nm heregulin EGF i minutter på is, vasket to ganger og inkubert med en 1:0 fortynning av streptavidin-pe i 4 minutter. For å vurdere bakgrunnsfarging fra streptavidin-pe-konjugatet, ble 1 x ADRr-celler inkubert med 0 l av en 1:0 fortynning av streptavidin-pe bare i 4 minutter. [023] Resultatene av dette forsøket er vist i figurene 18A og 18B. Som vist i figurene 18A og 18B, var både Ab #6 og Ab/FAb #3 i stand til å inhibere heregulinbinding til ErbB3. [0236] Likeledes ble evnen Ab #6 har til å inhibere bindingen av en annen ErbB3-ligand, epiregulin, til ErbB3, undersøkt som følger. [0237] ADRr-celler (1 x ) ble pre-inkubert med 2 μm Ab #6 eller 2 M Erbitux (som kontroll) i 0 l BD 2 fargebuffer i minutter på is. Etter minutter ble 0 l 2 M biotinylert Epi tilsatt til cellene og inkubert i ytterligere minutter på is. Dette ga en sluttkonsentrasjon på 12, M antistoffer og 1 M Epi. Celler ble så vasket to ganger med 00 l BD fargebuffer og inkubert med 0 l av en 1:0 fortynning av streptavidin-pe (PE = fycoerytrin) (Invitrogen) i BD fargebuffer i 4 minutter. Til slutt ble cellene vasket to ganger, resuspendert i 0 l BD fargebuffer og analysert i et FACScalibur strømningscytometer. Som en positiv kontroll, ble 1 x ADRr-celler inkubert med 1 M Epi i minutter på is, vasket to ganger og inkubert med en 1:0 fortynning av streptavidin- PE i 4 minutter. For å vurdere bakgrunnsfarging fra streptavidin-pe-konjugatet ble celler inkubert med 0 l av en 1:0 fortynning av streptavidin-pe bare i 4 minutter. 3 [0238] Resultatene av dette forsøket er avbildet i figurene 19A og 19B. Som vist i figur 19A, binder epiregulin til ErbB3-positive ADRr-celler. Videre, som vist i figur 19B, blir denne bindingen inhibert av både Erbitux og Ab #6, noe som foreslår at epiregulin kan binde til både EGFR og ErbB3. [0239] Et ytterligere eksperiment ble utført for å undersøke om Ab #6 er i stand til å inhibere bindingen av en EGFlignende ligand (f.eks. HB-EGF) til tumorceller.
56 [0240] ADRr-celler (1 x ) ble pre-inkubert med 2 μm Ab #6 eller 2 M Erbitux (som kontroll) i 0 l BD fargebuffer i minutter på is. Etter minutter ble 0 l 400 nm biotinylert HB-EGF tilsatt til cellene og inkubert i ytterligere minutter på is. Dette ga en sluttkonsentrasjon på 12, M antistoffer og 0 nm HB-EGF. Celler ble så vasket to ganger med 00 l BD fargebuffer og inkubert med 0 l av en 1:0 fortynning av streptavidin-pe (PE = fycoerytrin) (Invitrogen) i BD fargebuffer i 4 minutter. Til slutt ble celler vasket to ganger, resuspendert i 0 l BD fargebuffer og analysert i et FACScalibur strømningscytometer. Som en positiv kontroll, ble 1 x ADRrceller inkubert med 0 nm HB-EGF i minutter på is, vasket to ganger og inkubert med en 1:0 fortynning av streptavidin-pe i 4 minutter. For å vurdere bakgrunnsfarging fra streptavidin-pe-konjugatet ble celler inkubert med 0 l av en 1:0 fortynning av streptavidin-pe bare i 4 minutter. [0241] Som vist i figurene A og B, binder HB-EGF til ErbB på ADRr-celler og Ab #6 inhiberer ikke denne 1 bindingen, noe som beviser at Ab #6 er spesifikk for inhibering av bindingen av ErbB3-ligander (f.eks. heregulin og epiregulin) til ErbB3. EKSEMPEL 11: Inhibering av VEGF-sekresjon i tumorceller 2 [0242] Evnen Ab #6 har til å inhibere VEGF-sekresjon fra celler som uttrykker ErbB3 (f.eks. kreftceller) ble undersøkt ved anvendelse av VEGF sekresjonsanalysen (VEGF ELISA kit tilgjengelig fra R&D Systems, Minneapolis, MN, kat.# DY293B). Først ble evnen Ab #6 har til å inhibere VEGF sekresjon i ubehandlet og HRG-beta 1 behandlet MCF-7, T47D og COLO-37 celler analysert. Disse studiene viste at COLO-37 utskilte den høyeste mengden VEGF til mediene. Ettersom disse cellene også hadde svært høye HRG-nivåer (data ikke vist), var tilsetning av HRG til mediene ikke i stand til å ytterligere indusere VEGF-utskilling (Fig. 24A). I motsetning til dette var HRG i stand til å indusere VEGF-utskilling i MCF-7-og T47D-celler. [0243] Ab #6 viser en potent inhiberende virkning ved høye nivåer i alle tre cellelinjene med den høyeste i COLO- 37 (fig. 24A). Ab #6 viser også en lignende effekt in vivo ved inhibering av VEGF-sekresjon i tre forskjellige xenografter, den høyeste er i COLO-37 xenograft (fig. 24B). Inhibering av VEGF korrelerer med inhibering av ErbB3-fosforylering (Fig. 24 C). Inhibering av VEGF-sekresjon korrelerer også med inhibering av angiogenese av tumorcellene. Spesielt har det blitt identifisert at myelomcelle-utskilte faktorer, så som VEGF og bfgf, utløser angiogenese (se, f.eks. Leung et al. (1989) Science 246 (493): 16-9; Yen et al. (00) Oncogene 19 (31) :3460-9). Eksempel 12: Inhibering av cellemigrering 3 [0244] Evnen Ab #6 har til å inhibere migreringen av celler som uttrykker ErbB3 (f.eks. MCF-7 celler) ble undersøkt ved anvendelse av en transbrønn-well analyse (Millipore Corp, Billerica, MA, kat # ECM2). Først ble MCF-7 celler serumsultet over natten og så inkubert i nærværet eller fraværet av Ab #6 (8 um sluttkonsentrasjon) i 1 minutter ved romtemperatur. Cellene ble så overført til et øvre kammer som er atskilt fra et nedre kammer av en kollagen
57 6 type I-belagt membran som cellene kan migrere gjennom. % FBS ble tilsatt til mediet i det nedre kammeret for å virke som en kjemoattraktant i nærværet eller fraværet av Ab #6. Kamrene ble inkubert ved 37 C i 16 timer og så ble cellene som migrerte gjennom membranen fjernet ved anvendelse av en løsgjøringsbuffer og inkubert med et celle-bindende fluorescerende fargestoff. Fluorescens ble kvantifisert ved anvendelse av en fluorescerende plateavleser. Den gjennomsnittlige fluorescensen ± SEM (n = 2) er vist i figur 2. [024] Som vist i figur 2, stimulerer % FBS cellemigrering (bane 3), sammenlignet med ubehandlet kontroll (bane 1) og 8 um Ab #6 inhiberer den FBS-induserte cellemigreringen (bane 4). Eksempel 13: Inhibering av sfæroidevekst [0246] Evnen Ab #6 har til å inhibere sfæroideveksten av celler som uttrykker ErbB3 ble undersøkt ved anvendelse 1 av en analyse som tilnærmes betingelser av en utviklende tumorvekst (Herman et al. (07) Journal of Biomolecular Screening elektronisk publikasjon). AdrR og DU14 sfæroider ble initiert ved en frekvens på 1 sfæroide per brønn av en 96 brønn plate ved anvendelse av hengende dråpe-metoden (Herrman et al. 08). Individuelle sfæroider ble så behandlet med enten Ab #6 (8 um sluttkonsentrasjon), Heregulin-β1 EGF-domene (R&D Systems, Minneapolis, MN, kat #396-HB, 3,4 nm sluttkonsentrasjon), eller en kombinasjon av begge, som indikert. Diameterene av sfæroidene ble målt ved anvendelse av lysmikroskopi (X objektiv) på dag 1 og dag 13. [0247] Ab #6 inhiberer sfæroidevekst i AdrR-celler (Fig. 26A). I tillegg stimulerer 3,4 nm HRG sfæroidevekst og Ab #6 inhiberer HRG-effekten (Fig. 26B). Sfæroider avledet fra DU14 økte ikke i størrelse i løpet av 13 dager av forsøket, vekst ble imidlertid signifikant stimulert ved HRG1-beta 1. I disse cellene inhiberer 8 um Ab #6 HRGindusert sfæroidevekst (Fig. 26C). 2 Eksempel 14: Inhibering av signalisering [0248] Evnen Ab #6 har til å inhibere signalisering indusert ved forskjellige ligander ble undersøkt. For eksempel ble effekten av Ab #6 på HRG og BTC-binding til AdrR-celler som uttrykker ErbB3 reseptor testet. Som vist i figurene 27A og B, ved anvendelse av FACS-analyse, konkurrerer Ab #6 med HRG og ikke BTC for binding til AdrRceller: Tilsvarende ville blokkering av Ab #6 av HRG-binding til ErbB3 forhindre signalisering indusert ved HRG. [0249] I tillegg ble ulike ligander testet for indusering av ErBb3 fosforylering. Tre ligander, HRG, BTC og HGF, var i 3 stand til å stimulere ErbB3 indusert fosforylering i AdrR-celler, mens EGF ikke kunne det. Som vist i figur 28, inhiberer Ab #6 HGF-indusert perbb3-fosforylering i AdrR-celler (Fig. 28). Videre, som kjent innen teknikken (se f.eks. Wallenius et al. (00) Am J Pathol. 16 (3) : ), har forbedret HGF signalisering blitt funnet i forskjellige epiteliale og ikke-epiteliale tumorer.
58 7 ErbB3/cMET vekselvirkning og rollen av Ab #6 i modulering av denne vekselvirkningen [0] Det har blitt vist at ikke-småcellet lungekreft som bærer aktiveringsmutasjoner i den epidermale vekstfaktorreseptoren (EGFR) utvikler resistens mot tyrosinkinaseinhibitorer ved å rekruttere MET og HER3 og således aktivere PI3K-AKT celleoverlevelsesveien (Engelmann et al. (07) Science 316: 39-43; Gou (07) PNAS: (2): ). Assosiasjonen mellom EGFR og c-met i cellelinjer som bærer aktiverende EGFR mutasjoner har blitt godt etablert ved sam-immunoutfelling (Engelmann et al. 07; Gou 07). Guo et al. viste nylig at c-met og ErbB3 også eksisterer i et kompleks i en gastrisk cellelinje MKN4 kjent for å være avhengig av amplifisert c-met, ved anvendelse av sam-immunoutfelling. [021] Denne c-met-erbb3 vekselvirkningen forekommer også i AdrR-celler som bærer villtype EGFR og er ikke avhengig av forsterket c-met. HGF (hepatocytt vekstfaktor) induserer ErbB3-fosforylering i AdrR-celler på en doseavhengig måte som vist i figur 28. I tillegg inhiberer Ab #6 HGF-indusert fosforylering erbb3. 1 [022] Effekten av HRG og BTC på både ErbB1 og ErbB3-fosforyleringen har også blitt undersøkt, og HRG og BTC ble funnet å indusere fosforylering av både ErbB1 og ErbB3. HRG ble funnet å være en mer potent induserer av ErbB3-fosforylering mens BTC var en potent induserer av ErbB1 fosforylering (Fig. 29). Denne fosforyleringen er sannsynligvis drevet av komplekset mellom ErbB1 og ErbB3. Kort fortalt induserer HRG-binding til ErbB3 kompleksdannelse mellom ErbB1 og ErbB3, noe som fører til aktiveringen av begge reseptorene. Det samme fenomenet forekommer sannsynligvis for BTC, hvor BTC binding til ErbB1 stimulerer kompleksdannelse mellom ErbB1 og ErbB3, noe som fører til fosforylering av både ErbB1 og ErbB3. Antistoffinhibering av ligand (HRG, BTC, EGF og HGF) stimulert ErbB3-fosforylering. 2 3 [023] Evnen Ab #6 har til å inhibere ligand (HRG, BTC, EGF og HGF) indusert ErbB3-fosforylering ble undersøkt basert på den følgende metoden: 1. AdrR-celler ble plettert i 96 brønnplate ved en tetthet på.000 celler/brønn/0 ul i RPMI medium som inneholder % FBS og tillatt å vokse over natten; 2. Neste dag ble celler serum-sultet ved å endre medium til FBS-fritt medium og tillatt å vokse over natten; 3. Celler ble forbehandlet med forskjellige konsentrasjoner av Ab #6 (fra 0,01 nm til 0 nm) eller buffer (kontroll), i 2 timer; 4. Cellene ble så stimulert med nm HRG og HGF i minutter, eller nm BTC og EGF i minutter;. Reaksjonen ble stoppet ved å fjerne kulturmediet, og vasking av cellene én gang med iskald PBS; 6. Cellene ble så lysert i 2 mm Tris, ph +7,, mm NaCl, 1 mm EDTA, 1,0 % Triton X-0, 1,0 % CHAPS, % v/v glyserol, som inneholder 1X proteaseinhibitor og 1X fosfataseinhibitor og; 7. ErbB3-fosforylering ble målt i cellelysater med Human Phospho-ErbB3 ELISA kit (R&D Systems, Minneapolis, MN, kat. nr DYC1769) i henhold til produsentens instruksjoner.
59 8 1 Antistoffinhibering av ErbB2-ErbB3 proteinkompleksdannelse. [024] AdrR-celler ble pre-inkubert med buffer (kontroll) eller nm Ab #6 i 60 minutter ved romtemperatur, så behandlet med nm HRG eller nm BTC eller kontrollbuffer i minutter. Cellene ble lysert i 2 mm Tris, ph +7,, mm NaCl, 1 mm EDTA, 1,0 % Triton X-0, 1,0 % CHAPS, % v/v glyserol, inneholdende 0,2 mm PMSF, 0 mtu/ml aprotinin og 0 um leupeptin, og det ubearbeidede lysatet ble sentrifugert kort for å fjerne uløselig materiale. Supernatant ble overført til et nytt Eppendorf-rør, og anti-erbb3-antistoff (Santa Cruz sc-28) ble tilsatt ved 1:00 fortynning. Supernatanter ble inkubert over natten med forsiktig risting ved 4C. 60 ul av Immobilized Protein A/G agarosekuler (Pierce, Rockford, IL, kat # 421) ble først vasket med 1X PBS. Cellelysat-antistoffblandingen ble tilsatt til de PBS-vaskede kulene, og inkubert i 2 timer med forsiktig risting ved 4 C. Immunoutfellingene ble så vasket med iskald lysisbuffer 3 ganger, resuspendert i ul 2X SDS prøvebuffer, varmedenaturert ved 9 C i 7 minutter og kjørt på 4-12 % Bis-Tris geler. SDS-PAGE og elektro-overført til PVDF membran i Tri-glysin-buffer med % MeOH. Membranen ble blokkert i 1 time i ml blokkeringsbuffer (Li-Cor Biosciences, Lincoln, NE, kat# ) og så inkubert med anti-erbb2-antistoffet ved 1:00 (Cell Signaling Technology, Danvers, MA, kat # 29D8) i ml blokkeringsbuffer (Li-Cor Biosciences, kat# ). Signalet ble detektert ved anvendelse av geite-anti-kanin IRDye800 ved 1:000 (2 ul) i ml blokkeringsbuffer (Li-Cor Biosciences, kat # ). [02] Ab #6 ble også vist å fullstendig inhibere HRG stimulert ErbB2/3 kompleksdannelse (Fig. 29B).
60 9 P a t e n t k r a v 1. Isolert monoklonalt antistoff eller antigenbindende del derav som binder til human ErbB3 og omfatter: 1 (a) en tung kjede variabel region, hvori CDR1en omfatter aminosyresekvensen ifølge SEKV ID NR: 7, CDR2en omfatter aminosyresekvensen ifølge SEKV ID NR: 8 og CDR3en omfatter aminosyresekvensen ifølge SEKV ID NR: 9, og en lett kjede variabel region, hvori CDR1en omfatter aminosyresekvensen ifølge SEKV ID NR:, CDR2en omfatter aminosyresekvensen ifølge SEKV ID NR: 11 og CDR3en omfatter aminosyresekvensen ifølge SEKV ID NR: 12; eller (b) en tung kjede variabel region, hvori CDR1en, CDR2en og CDR3en omfatter aminosyresekvenser som er minst 90 % identiske med CDR1, CDR2 og CDR3 aminosyresekvensene definert i (a), og en lett kjede variabel region, hvori CDR1en, CDR2en og CDR3en omfatter aminosyresekvenser som er minst 90 % identiske med CDR1, CDR2 og CDR3 aminosyresekvensene definert i (a), og hvori antistoffet eller antigenbindende del derav, er stand til å inhibere EGF-lignende ligandmediert fosforylering av ErbB3. 2. Antistoff eller antigenbindende del derav ifølge krav 1, hvori tung kjede CDR1en omfatter aminosyresekvensen ifølge SEKV ID NR: 7, tung kjede CDR2en omfatter aminosyresekvensen ifølge SEKV ID NR: 8, tung kjede CDR3en omfatter aminosyresekvensen ifølge SEKV ID NR: 9, lett kjede CDR1en omfatter aminosyresekvensen ifølge SEKV ID NR:, lett kjede CDR2en omfatter aminosyresekvensen ifølge SEKV ID NR: 11 og lett kjede CDR3en omfatter aminosyresekvensen ifølge SEKV ID NR: Antistoff eller antigenbindende del derav ifølge krav 1 eller 2, hvori tung kjede variabel regionen omfatter 2 aminosyresekvensen ifølge SEKV ID NR: 1 og lett kjede variabel regionen omfatter aminosyresekvensen ifølge SEKV ID nr Antistoff eller antigenbindende del derav ifølge krav 1, hvori den EGF-lignende liganden er betacellulin.. Antistoff eller antigenbindende del derav ifølge krav 4, som er i stand til å inhibere betacellulinmediert fosforylering av ErbB3 i AdrR-celler in vitro. 6. Antistoff eller antigenbindende del derav ifølge ett av de foregående krav, hvori antistoffet er valgt fra gruppen bestående av et humant antistoff, et humanisert antistoff, et bispesifikt antistoff og et kimært antistoff Antistoff eller antigenbindende del derav ifølge ett av de foregående krav, hvori antistoffet eller antigenbindende del derav er valgt fra gruppen bestående av et Fab, Fab'2, scfv og et domene-antistoff.
61 60 8. Antistoff eller antigenbindende del derav ifølge ett av de foregående krav, hvori antistoff-isotypen er valgt fra gruppen bestående av et IgG1, et IgG2, et IgG3, et IgG4, et IgM, et IgA1, et IgA2, et IgAsec, et IgD og et IgE antistoff. 9. Sammensetning som omfatter antistoffet eller antigenbindende del ifølge ett av de foregående krav i en farmasøytisk akseptabel bærer.. Isolert nukleinsyresammensetning som koder for et humant antistoff som binder human ErbB3 ifølge krav 1, denne sammensetningen omfatter en nukleinsyre som omfatter en sekvens som koder for en tung kjede variabel region, denne sekvensen er minst 90 % identisk med, eller er en sekvens som hybridiserer under høy stringens til, SEKV ID NR: 2 og en nukleinsyre som omfatter en sekvens som koder for en lett kjede variabel region, denne sekvensen er minst 90 % identisk med, eller er en sekvens som hybridiser under høy stringens til SEKV ID NR: Nukleinsyresammensetning ifølge krav, hvori nukleotidsekvensen som koder for en tung kjede variabel region omfatter SEKV ID NR: 2 og nukleotidsekvensen som koder for en lett kjede variabel region omfatter SEKV ID NR: Nukleinsyresammensetning ifølge krav eller 11, hvori hver nukleinsyre er omfattet i en ekspresjonsvektor. 13. Vertscelle som omfatter nukleinsyresammensetningen ifølge ett av kravene til Transgent ikke-humant pattedyr eller en transgen plante som uttrykker antistoffet eller antigenbindende del ifølge ett av kravene 1 til Vertscelle som produserer et antistoff eller antigenbindende del derav ifølge ett av kravene 1 til Vertscelle ifølge krav 1, hvori antistoffet er kodet for av en tung kjede variabel region nukleotidsekvens fremlagt i SEKV ID NR: 2 og en lett kjede variabel region nukleotidsekvens fremlagt i SEKV ID NR: 26, og konservative sekvensmodifikasjoner derav Kit som omfatter et isolert monoklonalt antistoff eller antigenbindende del derav ifølge ett av kravene 1 til 8, og instruksjoner for anvendelse ved behandling eller diagnostisering av en sykdom assosiert med ErbB3 avhengig signalisering eventuelt hvori sykdommen er kreft. 18. Antistoff eller antigenbindende del derav ifølge ett av kravene 1 til 8 for anvendelse i en fremgangsmåte for å inhibere EGF-lignende ligandmediert fosforylering av ErbB3 eller behandling av kreft i et humant subjekt.
62 Antistoff eller antigenbindende del derav ifølge ett av kravene 1 til 8 for anvendelse ifølge krav 18, hvori kreften er kreft i bukspyttkjertelen, melanom, brystkreft, eggstokk-kreft, nyrekarsinom, gastrointestianal/colonkreft, lungekreft, klarcellesarkom eller prostatakreft.. Antistoff eller antigenbindende del derav ifølge ett av kravene 1 til 8 for anvendelse ifølge krav 18, hvori: - antistoffet eller antigenbindende del derav blir administrert intravenøst, intramuskulært eller subkutant til subjektet, og/eller - antistoffet eller antigenbindende del derav blir administrert i kombinasjon med et andre terapeutisk middel, hvori det andre terapeutiske middel er (a) et andre anti-kreft-antistoff, eller (b) et anti-kreft small molecule Antistoff eller antigenbindende del derav ifølge ett av kravene 1 til 8 for anvendelse ifølge krav, hvori anti-kreft-antistoffet er et anti-igf1r antistoff, et anti-egfr-antistoff eller et anti-cmet antistoff. 22. Antistoff eller antigenbindende del derav ifølge ett av kravene 1 til 8 for anvendelse ifølge krav, hvori det gjeldende anti-kreft small molecule er en antimetabolitt, et alkyleringsmiddel, en topoisomeraseinhibitor, et mikrotubulimålrettende middel, en kinaseinhibitor, en proteinsynteseinhibitor, et immunoterapeutikum, et hormon eller analog derav, en somatostatinanalog, et glukokortokoid, en aromataseinhibitor, en mtor-inhibitor, et small molecule som målsøker IGF1R eller et small molecule som målsøker EGFR Antistoff eller antigenbindende del derav ifølge ett av kravene 1 til 8 for anvendelse i en fremgangsmåte for å diagnostisere kreft assosiert med ErbB3 i et humant subjekt, som omfatter (a) å bringe ex vivo eller in vivo celler fra subjektet i kontakt med nevnte antistoff eller antigenbindende del derav, og (b) måle nivået av binding av nevnte antistoff eller antigenbindende del til ErbB3 på cellene, hvori unormalt høye nivåer av binding av nevnte antistoff eller antigenbindende del til ErbB3 indikerer at subjektet har en kreft assosiert med ErbB3.
63
64
65
66
67
68
69
70
71
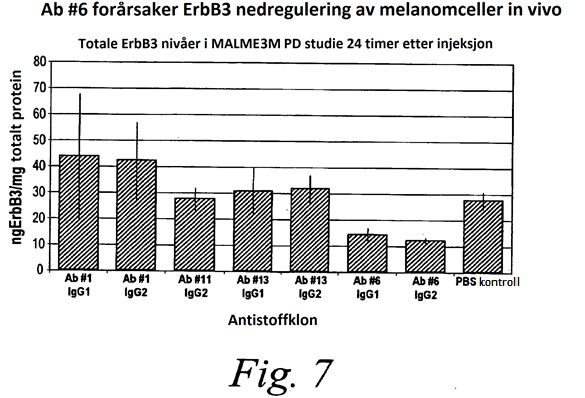
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91

92
93
94

95

96

97
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2310382 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/4412 (2006.01) A61P 35/00 (2006.01) C07D 401/14 (2006.01) C07D 403/12 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2310382 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/4412 (2006.01) A61P 35/00 (2006.01) C07D 401/14 (2006.01) C07D 403/12 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2178851 B1 (19) NO NORGE (51) Int Cl. C07D 261/08 (2006.01) A61K 31/42 (2006.01) A61P 3/06 (2006.01) C07D 413/12 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2178851 B1 (19) NO NORGE (51) Int Cl. C07D 261/08 (2006.01) A61K 31/42 (2006.01) A61P 3/06 (2006.01) C07D 413/12 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2148670 B1 (19) NO NORGE (51) Int Cl. A61K 31/137 (2006.01) A61P 25/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.02 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2148670 B1 (19) NO NORGE (51) Int Cl. A61K 31/137 (2006.01) A61P 25/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.02 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2129377 B1 (19) NO NORGE (51) Int Cl. A61K 31/451 (2006.01) A61K 9/08 (2006.01) A61P 25/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.23
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2129377 B1 (19) NO NORGE (51) Int Cl. A61K 31/451 (2006.01) A61K 9/08 (2006.01) A61P 25/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.23
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 240726 B1 (19) NO NORGE (1) Int Cl. H0K 3/36 (2006.01) H0K 3/42 (2006.01) H0K 3/46 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.17 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 240726 B1 (19) NO NORGE (1) Int Cl. H0K 3/36 (2006.01) H0K 3/42 (2006.01) H0K 3/46 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.17 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2274977 B1 (19) NO NORGE (1) Int Cl. A01K 83/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2274977 B1 (19) NO NORGE (1) Int Cl. A01K 83/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2114970 B1 (19) NO NORGE (51) Int Cl. C07F 9/58 (2006.01) A61K 31/44 (2006.01) A61P 1/00 (2006.01) A61P 11/06 (2006.01) A61P 19/02 (2006.01) A61P
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2114970 B1 (19) NO NORGE (51) Int Cl. C07F 9/58 (2006.01) A61K 31/44 (2006.01) A61P 1/00 (2006.01) A61P 11/06 (2006.01) A61P 19/02 (2006.01) A61P
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2246321 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/135 (2006.01) C07C 211/42 (2006.01) Patentstyret (21) Oversettelse publisert 2011.12.12
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2246321 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/135 (2006.01) C07C 211/42 (2006.01) Patentstyret (21) Oversettelse publisert 2011.12.12
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2125711 B1 (19) NO NORGE (51) Int Cl. C07C 321/20 (2006.01) A61K 31/216 (2006.01) A61K 31/421 (2006.01) A61K 31/4402 (2006.01) A61K 31/495 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2125711 B1 (19) NO NORGE (51) Int Cl. C07C 321/20 (2006.01) A61K 31/216 (2006.01) A61K 31/421 (2006.01) A61K 31/4402 (2006.01) A61K 31/495 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2011486 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.09.17 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2011486 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.09.17 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2445326 B1 (19) NO NORGE (51) Int Cl. H05K 5/02 (2006.01) B43K 23/12 (2006.01) B43K 24/06 (2006.01) H01R 13/60 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2445326 B1 (19) NO NORGE (51) Int Cl. H05K 5/02 (2006.01) B43K 23/12 (2006.01) B43K 24/06 (2006.01) H01R 13/60 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 270722 B1 (19) NO NORGE (1) Int Cl. F21V 23/02 (06.01) F21S 8/02 (06.01) F21V 23/00 (06.01) Patentstyret (21) Oversettelse publisert 14.03. (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 270722 B1 (19) NO NORGE (1) Int Cl. F21V 23/02 (06.01) F21S 8/02 (06.01) F21V 23/00 (06.01) Patentstyret (21) Oversettelse publisert 14.03. (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2272978 B1 (19) NO NORGE (51) Int Cl. C12Q 1/68 (2006.01) Patentstyret (21) Oversettelse publisert 2012.08.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2272978 B1 (19) NO NORGE (51) Int Cl. C12Q 1/68 (2006.01) Patentstyret (21) Oversettelse publisert 2012.08.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP2770 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2770 B1 (19) NO NORGE (1) Int Cl. B23K 3/00 (06.01) C21D 6/00 (06.01) C21D 9/04 (06.01) C22C 38/00 (06.01) C22C 38/44 (06.01) Patentstyret
NO/EP2770 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2770 B1 (19) NO NORGE (1) Int Cl. B23K 3/00 (06.01) C21D 6/00 (06.01) C21D 9/04 (06.01) C22C 38/00 (06.01) C22C 38/44 (06.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP22342 (12) Oversettelse av europeisk patentskrift (11) NO/EP 22342 B1 (19) NO NORGE (1) Int Cl. F2D 23/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske Patentmyndighets
NO/EP22342 (12) Oversettelse av europeisk patentskrift (11) NO/EP 22342 B1 (19) NO NORGE (1) Int Cl. F2D 23/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 223094 B1 (19) NO NORGE (1) Int Cl. A43B 7/32 (06.01) A43B 7/12 (06.01) A43B 7/34 (06.01) A43B 13/12 (06.01) A43B 13/41 (06.01) B29D 3/14 (.01) Patentstyret
(12) Oversettelse av europeisk patentskrift (11) NO/EP 223094 B1 (19) NO NORGE (1) Int Cl. A43B 7/32 (06.01) A43B 7/12 (06.01) A43B 7/34 (06.01) A43B 13/12 (06.01) A43B 13/41 (06.01) B29D 3/14 (.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2613860 B1 (19) NO NORGE (51) Int Cl. B01D 15/18 (2006.01) C11B 3/10 (2006.01) C11C 1/00 (2006.01) C11C 1/08 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2613860 B1 (19) NO NORGE (51) Int Cl. B01D 15/18 (2006.01) C11B 3/10 (2006.01) C11C 1/00 (2006.01) C11C 1/08 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2170890 B1 (19) NO NORGE (51) Int Cl. C07D 487/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.03.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2170890 B1 (19) NO NORGE (51) Int Cl. C07D 487/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.03.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 213696 B1 (19) NO NORGE (1) Int Cl. B23K 9/32 (2006.01) B23K 9/28 (2006.01) Patentstyret (21) Oversettelse publisert 2014.04.07 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 213696 B1 (19) NO NORGE (1) Int Cl. B23K 9/32 (2006.01) B23K 9/28 (2006.01) Patentstyret (21) Oversettelse publisert 2014.04.07 (80) Dato for Den
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2238877 B1 (19) NO NORGE (1) Int Cl. A47J 31/08 (06.01) Patentstyret (21) Oversettelse publisert 13.03.11 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2238877 B1 (19) NO NORGE (1) Int Cl. A47J 31/08 (06.01) Patentstyret (21) Oversettelse publisert 13.03.11 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2216387 B1 (19) NO NORGE (51) Int Cl. C10L 5/44 (2006.01) C10L 5/14 (2006.01) C10L 5/36 (2006.01) Patentstyret (21) Oversettelse publisert 2013.05.06
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2216387 B1 (19) NO NORGE (51) Int Cl. C10L 5/44 (2006.01) C10L 5/14 (2006.01) C10L 5/36 (2006.01) Patentstyret (21) Oversettelse publisert 2013.05.06
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 242166 B1 (19) NO NORGE (1) Int Cl. G06K 19/077 (06.01) G06K 19/06 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 242166 B1 (19) NO NORGE (1) Int Cl. G06K 19/077 (06.01) G06K 19/06 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2285808 B1 (19) NO NORGE (51) Int Cl. C07D 471/20 (2006.01) A61K 31/407 (2006.01) A61K 31/424 (2006.01) A61K 31/437 (2006.01) A61K 31/438 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2285808 B1 (19) NO NORGE (51) Int Cl. C07D 471/20 (2006.01) A61K 31/407 (2006.01) A61K 31/424 (2006.01) A61K 31/437 (2006.01) A61K 31/438 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 11438 B1 (19) NO NORGE (1) Int Cl. E04B 1/343 (06.01) B63B 29/02 (06.01) Patentstyret (21) Oversettelse publisert.02.23 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 11438 B1 (19) NO NORGE (1) Int Cl. E04B 1/343 (06.01) B63B 29/02 (06.01) Patentstyret (21) Oversettelse publisert.02.23 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse
 Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse") (12) Oversettelse av europeisk patentskrift (11) NO/EP 217368 B2 (19) NO NORGE (1) Int Cl. B42D / (06.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse publisert.04. (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 217368 B2 (19) NO NORGE (1) Int Cl. B42D / (06.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse publisert.04. (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2672278 B1 (19) NO NORGE (1) Int Cl. G01R 1/067 (2006.01) G01R 1/04 (2006.01) G01R 19/1 (2006.01) Patentstyret (21) Oversettelse publisert 201.04.20
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2672278 B1 (19) NO NORGE (1) Int Cl. G01R 1/067 (2006.01) G01R 1/04 (2006.01) G01R 19/1 (2006.01) Patentstyret (21) Oversettelse publisert 201.04.20
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2636033 B1 (19) NO NORGE (51) Int Cl. Patentstyret G09B 23/28 (2006.01) G09B 23/30 (2006.01) (21) Oversettelse publisert 2015.11.09 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2636033 B1 (19) NO NORGE (51) Int Cl. Patentstyret G09B 23/28 (2006.01) G09B 23/30 (2006.01) (21) Oversettelse publisert 2015.11.09 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2128505 B1 (19) NO NORGE (51) Int Cl. F16L 9/12 (2006.01) F16L 3/14 (2006.01) F16L 11/127 (2006.01) F24F 13/02 (2006.01) H05F 3/02 (2006.01) Patentstyret
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2128505 B1 (19) NO NORGE (51) Int Cl. F16L 9/12 (2006.01) F16L 3/14 (2006.01) F16L 11/127 (2006.01) F24F 13/02 (2006.01) H05F 3/02 (2006.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 237066 B1 (19) NO NORGE (1) Int Cl. E06C 1/12 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 237066 B1 (19) NO NORGE (1) Int Cl. E06C 1/12 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2252286 B1 (19) NO NORGE (51) Int Cl. A61K 31/357 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.16 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2252286 B1 (19) NO NORGE (51) Int Cl. A61K 31/357 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.16 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2096736 B1 (19) NO NORGE (1) Int Cl. H02K 1/32 (2006.01) H02K 3/24 (2006.01) H02K 9/00 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2096736 B1 (19) NO NORGE (1) Int Cl. H02K 1/32 (2006.01) H02K 3/24 (2006.01) H02K 9/00 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2173868 B1 (19) NO NORGE (51) Int Cl. C12N 9/50 (2006.01) C07K 14/415 (2006.01) C12N 15/29 (2006.01) C12N 15/57 (2006.01) C12N 15/81 (2006.01) A23J
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2173868 B1 (19) NO NORGE (51) Int Cl. C12N 9/50 (2006.01) C07K 14/415 (2006.01) C12N 15/29 (2006.01) C12N 15/57 (2006.01) C12N 15/81 (2006.01) A23J
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 88493 B1 (19) NO NORGE (1) Int Cl. G06F 1/00 (06.01) H01L 23/34 (06.01) G06F 1/ (06.01) Patentstyret (21) Oversettelse publisert 13.04.22 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 88493 B1 (19) NO NORGE (1) Int Cl. G06F 1/00 (06.01) H01L 23/34 (06.01) G06F 1/ (06.01) Patentstyret (21) Oversettelse publisert 13.04.22 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2082973 B1 (19) NO NORGE (1) Int Cl. B6D 81/34 (2006.01) Patentstyret (21) Oversettelse publisert 2014.06.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2082973 B1 (19) NO NORGE (1) Int Cl. B6D 81/34 (2006.01) Patentstyret (21) Oversettelse publisert 2014.06.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 218466 B1 (19) NO NORGE (1) Int Cl. B67C 3/26 (06.01) B6D 47/ (06.01) B67C 7/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02. (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 218466 B1 (19) NO NORGE (1) Int Cl. B67C 3/26 (06.01) B6D 47/ (06.01) B67C 7/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02. (80) Dato
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2384729 B1 (19) NO NORGE (1) Int Cl. A61G /12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2384729 B1 (19) NO NORGE (1) Int Cl. A61G /12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") 1 3 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2207775 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/5377 (2006.01) A61P 3/06 (2006.01) C07D 401/14 (2006.01) C07D 413/14 (2006.01)
1 3 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2207775 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/5377 (2006.01) A61P 3/06 (2006.01) C07D 401/14 (2006.01) C07D 413/14 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2317621 B1 (19) NO NORGE (1) Int Cl. H02G 3/12 (06.01) Patentstyret (21) Oversettelse publisert 1.02.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2317621 B1 (19) NO NORGE (1) Int Cl. H02G 3/12 (06.01) Patentstyret (21) Oversettelse publisert 1.02.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2146022 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.11.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2146022 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.11.03 (80) Dato for Den Europeiske Patentmyndighets
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2184425 B1 (19) NO NORGE (51) Int Cl. E05B 17/20 (2006.01) E05B 63/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.02.06 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2184425 B1 (19) NO NORGE (51) Int Cl. E05B 17/20 (2006.01) E05B 63/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.02.06 (80) Dato for
(12) Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse
 Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2175588 B2 (19) NO NORGE (51) Int Cl. H04L 12/14 (2006.01) H04L 29/08 (2006.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2175588 B2 (19) NO NORGE (51) Int Cl. H04L 12/14 (2006.01) H04L 29/08 (2006.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22799 B1 (19) NO NORGE (1) Int Cl. A61K 31/23 (06.01) A61K 31/047 (06.01) A61K 31/231 (06.01) A61K 31/232 (06.01) A61K 31/3 (06.01) A61K 31/93 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22799 B1 (19) NO NORGE (1) Int Cl. A61K 31/23 (06.01) A61K 31/047 (06.01) A61K 31/231 (06.01) A61K 31/232 (06.01) A61K 31/3 (06.01) A61K 31/93 (06.01)
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 21847 B1 (19) NO NORGE (1) Int Cl. F24F 7/08 (06.01) F24F 11/04 (06.01) F24F 12/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 21847 B1 (19) NO NORGE (1) Int Cl. F24F 7/08 (06.01) F24F 11/04 (06.01) F24F 12/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2231428 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) Patentstyret (21) Oversettelse publisert 12.11.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2231428 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) Patentstyret (21) Oversettelse publisert 12.11.26 (80) Dato for Den Europeiske Patentmyndighets
(86) Europeisk innleveringsdag
 Europeisk innleveringsdag") (12) Oversettelse av europeisk patentskrift (11) NO/EP 297978 B1 (19) NO NORGE (1) Int Cl. A41B 9/02 (06.01) Patentstyret (21) Oversettelse publisert 14.03.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 297978 B1 (19) NO NORGE (1) Int Cl. A41B 9/02 (06.01) Patentstyret (21) Oversettelse publisert 14.03.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2491293 B1 (19) NO NORGE (1) Int Cl. F17C 3/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.2 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2491293 B1 (19) NO NORGE (1) Int Cl. F17C 3/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.2 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2261144 B1 (19) NO NORGE (1) Int Cl. B6G 21/00 (06.01) B6G 21/08 (06.01) Patentstyret (21) Oversettelse publisert 13.07.08 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2261144 B1 (19) NO NORGE (1) Int Cl. B6G 21/00 (06.01) B6G 21/08 (06.01) Patentstyret (21) Oversettelse publisert 13.07.08 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2311023 B1 (19) NO NORGE (51) Int Cl. G09F 17/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2311023 B1 (19) NO NORGE (51) Int Cl. G09F 17/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2097141 B1 (19) NO NORGE (51) Int Cl. A62B 35/00 (2006.01) Patentstyret (21) Oversettelse publisert 2013.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2097141 B1 (19) NO NORGE (51) Int Cl. A62B 35/00 (2006.01) Patentstyret (21) Oversettelse publisert 2013.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2708433 B1 (19) NO NORGE (1) Int Cl. B61B 1/02 (2006.01) B61B 12/02 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.12 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2708433 B1 (19) NO NORGE (1) Int Cl. B61B 1/02 (2006.01) B61B 12/02 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.12 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 20789 B1 (19) NO NORGE (1) Int Cl. B61D 1/00 (06.01) B61D 17/ (06.01) B61D 23/00 (06.01) Patentstyret (21) Oversettelse publisert 12.06.04 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 20789 B1 (19) NO NORGE (1) Int Cl. B61D 1/00 (06.01) B61D 17/ (06.01) B61D 23/00 (06.01) Patentstyret (21) Oversettelse publisert 12.06.04 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22442 B1 (19) NO NORGE (1) Int Cl. G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13..28 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22442 B1 (19) NO NORGE (1) Int Cl. G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13..28 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 7044 B1 (19) NO NORGE (1) Int Cl. A61K 36/18 (06.01) A61K 33/04 (06.01) A61K 33/18 (06.01) A61K 33/ (06.01) A61K 36/22 (06.01) A61K 36/28 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 7044 B1 (19) NO NORGE (1) Int Cl. A61K 36/18 (06.01) A61K 33/04 (06.01) A61K 33/18 (06.01) A61K 33/ (06.01) A61K 36/22 (06.01) A61K 36/28 (06.01)
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 26098 B1 (19) NO NORWAY (1) Int Cl. C07K 14/08 (06.01) C12Q 1/70 (06.01) Norwegian Industrial Property Office (21) Translation Published 16..24
(12) Translation of european patent specification (11) NO/EP 26098 B1 (19) NO NORWAY (1) Int Cl. C07K 14/08 (06.01) C12Q 1/70 (06.01) Norwegian Industrial Property Office (21) Translation Published 16..24
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 261673 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) B60H 1/00 (06.01) Patentstyret (21) Oversettelse publisert 1.01.12 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 261673 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) B60H 1/00 (06.01) Patentstyret (21) Oversettelse publisert 1.01.12 (80) Dato for Den Europeiske
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 17118 B1 (19) NO NORGE (1) Int Cl. B60M 1/06 (06.01) B60M 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.09.29 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 17118 B1 (19) NO NORGE (1) Int Cl. B60M 1/06 (06.01) B60M 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.09.29 (80) Dato for Den Europeiske
(12) Translation of european patent specification
 Translation of european patent specification") NO/EP20 (12) Translation of european patent specification (11) NO/EP 20 B1 (19) NO NORWAY (1) Int Cl. C07K 16/28 (2006.01) A61P 3/00 (2006.01) A61P 37/00 (2006.01) Norwegian Industrial Property Office
NO/EP20 (12) Translation of european patent specification (11) NO/EP 20 B1 (19) NO NORWAY (1) Int Cl. C07K 16/28 (2006.01) A61P 3/00 (2006.01) A61P 37/00 (2006.01) Norwegian Industrial Property Office
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP28769 (12) Oversettelse av europeisk patentskrift (11) NO/EP 28769 B1 (19) NO NORGE (1) Int Cl. F17D 1/18 (06.01) F16L 3/00 (06.01) Patentstyret (21) Oversettelse publisert 1.04. (80) Dato for Den
NO/EP28769 (12) Oversettelse av europeisk patentskrift (11) NO/EP 28769 B1 (19) NO NORGE (1) Int Cl. F17D 1/18 (06.01) F16L 3/00 (06.01) Patentstyret (21) Oversettelse publisert 1.04. (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 224294 B1 (19) NO NORGE (1) Int Cl. F16K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.10 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 224294 B1 (19) NO NORGE (1) Int Cl. F16K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.10 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2213923 B1 (19) NO NORGE (1) Int Cl. F16L 19/02 (06.01) F16L 19/028 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2213923 B1 (19) NO NORGE (1) Int Cl. F16L 19/02 (06.01) F16L 19/028 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 08940 B1 (19) NO NORGE (1) Int Cl. B6D 2/2 (06.01) A47G 19/34 (06.01) B6D 83/06 (06.01) G01F 11/26 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 08940 B1 (19) NO NORGE (1) Int Cl. B6D 2/2 (06.01) A47G 19/34 (06.01) B6D 83/06 (06.01) G01F 11/26 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 19724 B1 (19) NO NORGE (1) Int Cl. B63H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 12.12. (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 19724 B1 (19) NO NORGE (1) Int Cl. B63H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 12.12. (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 222267 B1 (19) NO NORWAY (1) Int Cl. A01N 1/02 (06.01) A23G 9/38 (06.01) C07K 14/43 (06.01) Norwegian Industrial Property Office (21) Translation
(12) Translation of european patent specification (11) NO/EP 222267 B1 (19) NO NORWAY (1) Int Cl. A01N 1/02 (06.01) A23G 9/38 (06.01) C07K 14/43 (06.01) Norwegian Industrial Property Office (21) Translation
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 222 B1 (19) NO NORGE (1) Int Cl. F16F 1/376 (06.01) F16F 1/373 (06.01) F16F 1/08 (06.01) Patentstyret (21) Oversettelse publisert 13.02.18 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 222 B1 (19) NO NORGE (1) Int Cl. F16F 1/376 (06.01) F16F 1/373 (06.01) F16F 1/08 (06.01) Patentstyret (21) Oversettelse publisert 13.02.18 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP918 (12) Oversettelse av europeisk patentskrift (11) NO/EP 918 B1 (19) NO NORGE (1) Int Cl. H02J 7/00 (06.01) Patentstyret (21) Oversettelse publisert 14.02.03 (80) Dato for Den Europeiske Patentmyndighets
NO/EP918 (12) Oversettelse av europeisk patentskrift (11) NO/EP 918 B1 (19) NO NORGE (1) Int Cl. H02J 7/00 (06.01) Patentstyret (21) Oversettelse publisert 14.02.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22619 B1 (19) NO NORGE (1) Int Cl. B21D 1/4 (2006.01) B21K 21/04 (2006.01) F42B /02 (2006.01) F42B /188 (2006.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22619 B1 (19) NO NORGE (1) Int Cl. B21D 1/4 (2006.01) B21K 21/04 (2006.01) F42B /02 (2006.01) F42B /188 (2006.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 24012 B1 (19) NO NORGE (1) Int Cl. B2C 1/00 (2006.01) B2C 1/06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.12.22 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 24012 B1 (19) NO NORGE (1) Int Cl. B2C 1/00 (2006.01) B2C 1/06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.12.22 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2231500 B1 (19) NO NORGE (51) Int Cl. B66F 9/00 (2006.01) B60P 1/02 (2006.01) B60P 3/022 (2006.01) B62B 3/065 (2006.01) B66D 1/00 (2006.01) B66F 9/06
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2231500 B1 (19) NO NORGE (51) Int Cl. B66F 9/00 (2006.01) B60P 1/02 (2006.01) B60P 3/022 (2006.01) B62B 3/065 (2006.01) B66D 1/00 (2006.01) B66F 9/06
PATENTKRAV. og en første lett kjede og en andre lett kjede, hvor første og andre lette kjeder er forskjellige.
 1 PATENTKRAV 1. Isolert monoklonalt antistoff som bærer en forskjellig spesifisitet i hvert kombinasjonssete og som består av to kopier av et enkelt tung kjede polypeptid og en første lett kjede og en
1 PATENTKRAV 1. Isolert monoklonalt antistoff som bærer en forskjellig spesifisitet i hvert kombinasjonssete og som består av to kopier av et enkelt tung kjede polypeptid og en første lett kjede og en
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2243894 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2243894 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2217383 B1 (19) NO NORGE (1) Int Cl. B0B 12/00 (06.01) B0B 11/00 (06.01) G01F 11/02 (06.01) G01F 1/07 (06.01) G07C 3/04 (06.01) Patentstyret (21)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2217383 B1 (19) NO NORGE (1) Int Cl. B0B 12/00 (06.01) B0B 11/00 (06.01) G01F 11/02 (06.01) G01F 1/07 (06.01) G07C 3/04 (06.01) Patentstyret (21)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2497702 B1 (19) NO NORGE (1) Int Cl. B62H 3/02 (06.01) B62H /00 (06.01) B62M 6/80 (.01) Patentstyret (21) Oversettelse publisert 1.03.16 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2497702 B1 (19) NO NORGE (1) Int Cl. B62H 3/02 (06.01) B62H /00 (06.01) B62M 6/80 (.01) Patentstyret (21) Oversettelse publisert 1.03.16 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 21976 B1 (19) NO NORGE (1) Int Cl. F24J 2/1 (06.01) F16L 11/22 (06.01) F16L 9/14 (06.01) F16L 9/13 (06.01) F24J 2/46 (06.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 21976 B1 (19) NO NORGE (1) Int Cl. F24J 2/1 (06.01) F16L 11/22 (06.01) F16L 9/14 (06.01) F16L 9/13 (06.01) F24J 2/46 (06.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 264739 B1 (19) NO NORGE (1) Int Cl. A61F 13/00 (06.01) A61B 17/132 (06.01) A61F 13/64 (06.01) A61F 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 264739 B1 (19) NO NORGE (1) Int Cl. A61F 13/00 (06.01) A61B 17/132 (06.01) A61F 13/64 (06.01) A61F 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 216340 B1 (19) NO NORGE (1) Int Cl. B60C 11/11 (06.01) B60C 11/03 (06.01) B60C 11/12 (06.01) Patentstyret (21) Oversettelse publisert 12.12.03 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 216340 B1 (19) NO NORGE (1) Int Cl. B60C 11/11 (06.01) B60C 11/03 (06.01) B60C 11/12 (06.01) Patentstyret (21) Oversettelse publisert 12.12.03 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2264391 B1 (19) NO NORGE (1) Int Cl. F27D 3/1 (2006.01) C21B 7/12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.11.18 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2264391 B1 (19) NO NORGE (1) Int Cl. F27D 3/1 (2006.01) C21B 7/12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.11.18 (80) Dato for Den
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2173379 B1 (19) NO NORWAY (1) Int Cl. A61K 39/39 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2016.01.2 (80) Date
(12) Translation of european patent specification (11) NO/EP 2173379 B1 (19) NO NORWAY (1) Int Cl. A61K 39/39 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2016.01.2 (80) Date
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2244923 B1 (19) NO NORGE (1) Int Cl. B61K 9/ (2006.01) Patentstyret (21) Oversettelse publisert 2013.09.30 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2244923 B1 (19) NO NORGE (1) Int Cl. B61K 9/ (2006.01) Patentstyret (21) Oversettelse publisert 2013.09.30 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2148223 B1 (19) NO NORGE (1) Int Cl. G01V 3/ (06.01) G01V 3/24 (06.01) Patentstyret (21) Oversettelse publisert 13.03.04 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2148223 B1 (19) NO NORGE (1) Int Cl. G01V 3/ (06.01) G01V 3/24 (06.01) Patentstyret (21) Oversettelse publisert 13.03.04 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 229688 B1 (19) NO NORGE (1) Int Cl. B29B 17/02 (06.01) D21B 1/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.18 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 229688 B1 (19) NO NORGE (1) Int Cl. B29B 17/02 (06.01) D21B 1/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.18 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 21181 B1 (19) NO NORGE (1) Int Cl. F16L 2/00 (2006.01) F16L 33/26 (2006.01) H01P 1/04 (2006.01) Patentstyret (21) Oversettelse publisert 2013.10.28
(12) Oversettelse av europeisk patentskrift (11) NO/EP 21181 B1 (19) NO NORGE (1) Int Cl. F16L 2/00 (2006.01) F16L 33/26 (2006.01) H01P 1/04 (2006.01) Patentstyret (21) Oversettelse publisert 2013.10.28
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2477830 B1 (19) NO NORGE (1) Int Cl. B60K 1/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2477830 B1 (19) NO NORGE (1) Int Cl. B60K 1/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 273 B1 (19) NO NORGE (51) Int Cl. B41J 2/175 (06.01) Patentstyret (21) Oversettelse publisert 14.05.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 273 B1 (19) NO NORGE (51) Int Cl. B41J 2/175 (06.01) Patentstyret (21) Oversettelse publisert 14.05.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 211778 B1 (19) NO NORWAY (1) Int Cl. A61K 38/16 (06.01) C07K 7/08 (06.01) Norwegian Industrial Property Office (21) Translation Published 1.09.07
(12) Translation of european patent specification (11) NO/EP 211778 B1 (19) NO NORWAY (1) Int Cl. A61K 38/16 (06.01) C07K 7/08 (06.01) Norwegian Industrial Property Office (21) Translation Published 1.09.07
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 240126 B1 (19) NO NORGE (1) Int Cl. C07D 211/62 (06.01) A61K 31/16 (06.01) A61K 31/44 (06.01) A61K 31/0 (06.01) A61K 31/06 (06.01) C07D 7/277 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 240126 B1 (19) NO NORGE (1) Int Cl. C07D 211/62 (06.01) A61K 31/16 (06.01) A61K 31/44 (06.01) A61K 31/0 (06.01) A61K 31/06 (06.01) C07D 7/277 (06.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2113323 B1 (19) NO NORGE (1) Int Cl. B23B 31/02 (2006.01) B23B 31/20 (2006.01) Patentstyret (21) Oversettelse publisert 2012.11.19 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2113323 B1 (19) NO NORGE (1) Int Cl. B23B 31/02 (2006.01) B23B 31/20 (2006.01) Patentstyret (21) Oversettelse publisert 2012.11.19 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 23196 B1 (19) NO NORGE (1) Int Cl. A01M 7/00 (06.01) Patentstyret (21) Oversettelse publisert 13.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 23196 B1 (19) NO NORGE (1) Int Cl. A01M 7/00 (06.01) Patentstyret (21) Oversettelse publisert 13.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2190 B1 (19) NO NORGE (1) Int Cl. C23C 14/28 (06.01) C23C 14/0 (06.01) Patentstyret (21) Oversettelse publisert 12.04. (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2190 B1 (19) NO NORGE (1) Int Cl. C23C 14/28 (06.01) C23C 14/0 (06.01) Patentstyret (21) Oversettelse publisert 12.04. (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2146836 B1 (19) NO NORGE (1) Int Cl. A47G 9/ (06.01) B26D 3/00 (06.01) B26D 3/28 (06.01) B29C 44/6 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2146836 B1 (19) NO NORGE (1) Int Cl. A47G 9/ (06.01) B26D 3/00 (06.01) B26D 3/28 (06.01) B29C 44/6 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2242771 B1 (19) NO NORGE (1) Int Cl. C07K 16/28 (06.01) A61P 3/00 (06.01) Patentstyret (21) Oversettelse publisert 13.09. (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2242771 B1 (19) NO NORGE (1) Int Cl. C07K 16/28 (06.01) A61P 3/00 (06.01) Patentstyret (21) Oversettelse publisert 13.09. (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 211333 B1 (19) NO NORGE (1) Int Cl. B28B 7/00 (06.01) Patentstyret (21) Oversettelse publisert 13.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 211333 B1 (19) NO NORGE (1) Int Cl. B28B 7/00 (06.01) Patentstyret (21) Oversettelse publisert 13.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 238426 B1 (19) NO NORGE (1) Int Cl. G01S 1/68 (06.01) B63C 9/32 (06.01) F41B 13/00 (06.01) F41B 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 238426 B1 (19) NO NORGE (1) Int Cl. G01S 1/68 (06.01) B63C 9/32 (06.01) F41B 13/00 (06.01) F41B 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 9863 B1 (19) NO NORGE (1) Int Cl. E04B 2/96 (06.01) Patentstyret (21) Oversettelse publisert 13.09.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 9863 B1 (19) NO NORGE (1) Int Cl. E04B 2/96 (06.01) Patentstyret (21) Oversettelse publisert 13.09.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 198722 B1 (19) NO NORGE (1) Int Cl. C23C 4/06 (06.01) C23C 4/18 (06.01) Patentstyret (21) Oversettelse publisert 12.02.13 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 198722 B1 (19) NO NORGE (1) Int Cl. C23C 4/06 (06.01) C23C 4/18 (06.01) Patentstyret (21) Oversettelse publisert 12.02.13 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2003466 B1 (19) NO NORGE (1) Int Cl. G01S /02 (2010.01) Patentstyret (21) Oversettelse publisert 2014.07.14 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2003466 B1 (19) NO NORGE (1) Int Cl. G01S /02 (2010.01) Patentstyret (21) Oversettelse publisert 2014.07.14 (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2229212 B1 (19) NO NORWAY (51) Int Cl. A61N 1/36 (2006.01) H01L 27/144 (2006.01) Norwegian Industrial Property Office (21) Translation Published
(12) Translation of european patent specification (11) NO/EP 2229212 B1 (19) NO NORWAY (51) Int Cl. A61N 1/36 (2006.01) H01L 27/144 (2006.01) Norwegian Industrial Property Office (21) Translation Published
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2012637 B1 NORGE (19) NO (1) Int Cl. A47K 13/00 (2006.01) Patentstyret (4) Oversettelse publisert: 20.08.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2012637 B1 NORGE (19) NO (1) Int Cl. A47K 13/00 (2006.01) Patentstyret (4) Oversettelse publisert: 20.08.09 (80) Dato for Den Europeiske Patentmyndighets
EP Patentkrav
 1 EP26727 Patentkrav 1. Monoklonalt antistoff, eller et funksjonelt fragment derav som spesifikt gjenkjenner ett eller flere Siglec- polypeptider bestående av en aminosyresekvens som beskrevet i hvilket
1 EP26727 Patentkrav 1. Monoklonalt antistoff, eller et funksjonelt fragment derav som spesifikt gjenkjenner ett eller flere Siglec- polypeptider bestående av en aminosyresekvens som beskrevet i hvilket
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 246764 B1 (19) NO NORGE (1) Int Cl. F2C 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 246764 B1 (19) NO NORGE (1) Int Cl. F2C 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2131860 B1 (19) NO NORGE (51) Int Cl. A61K 39/395 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2131860 B1 (19) NO NORGE (51) Int Cl. A61K 39/395 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.24 (80) Dato for Den Europeiske Patentmyndighets