Uttrykk, aktivitet og sekresjon av legumain er forskjellig i M1- og M2-differensierte makrofager
|
|
|
- Elise Ulriksen
- 6 år siden
- Visninger:
Transkript
1 Uttrykk, aktivitet og sekresjon av legumain er forskjellig i M1- og M2-differensierte makrofager Nuriddin Abdukadir Mastergradsoppgave i farmasi Seksjon for farmasøytisk biovitenskap Farmasøytisk institutt Det matematiske-naturvitenskapelige fakultet UNIVERSITETET I OSLO September 2016 I
2 II
3 Uttrykk, aktivitet og sekresjon av legumain er forskjellig i M1- og M2-differensierte makrofager Nuriddin Abdukadir Mastergradsoppgave i farmasi Veiledere: Professor Harald Thidemann Johansen Professor Rigmor Solberg Doktorgradsstipendiat Ngoc Nguyen Lunde III
4 Nuriddin Abdukadir 2016 Uttrykk, aktivitet og sekresjon av legumain er forskjellig I M1- og M2-differensierte makrofager Nuriddin Abdukadir Trykk: Reprosentralen, Universitetet i Oslo IV
5 Forord Denne masteroppgaven ble utført ved Seksjon for farmasøytisk biovitenskap, ved Farmasøytisk institutt, Universitetet i Oslo i perioden august 2015 til september Jeg vil først og fremst rette en stor takk til mine hovedveiledere, professor Harald Thidemann Johansen og professor Rigmor Solberg. Deres gode veiledning, oppmuntring og hjelp med skriveprosessen har vært uunværlig og jeg er veldig takknemlig for å ha fått anledning til å jobbe i forskningsgruppen deres. Takk for det kunnskapsrike innspillet deres og alle de spennende diskusjonene. Takk til veileder og doktorgradsstipendiat Ngoc Nguyen Lunde for all hjelp på laboratoriet og gode råd. Din dedikasjon til ditt arbeid har vært både motiverende og imponerende. Takk til overingeniør Hilde Nilsen for opplæring i laboratoriearbeidet, all din praktiske bistand og ditt positive humør. Jeg vil også takke alle på masterkontoret for et minneverdig år. Spesielt takk til Mathias Austli, Kristina B. B. Larsen og Anh Duong for alle de hyggelige kaffepausene hvor vi har delt frustrasjoner og gleder. Sist, men ikke minst vil jeg takke min kjære samboer Tina for å ha vært min største støttespiller under hele studietiden. Oslo, september 2016 Nuriddin Abdukadir V
6 VI
7 Sammendrag Cysteinproteasen legumain (asparaginylendopeptidase, AEP) er assosiert med flere inflammatoriske sykdomstilstander. Ved aterosklerose er uttrykk og aktivitet av legumain høyere i ustabile, sammenlignet med stabile områder i aterosklerotiske plakk. Legumain er også lokalisert til makrofager i aterosklerotiske lesjoner, noe som indikerer at denne proteasen hovedsakelig uttrykkes av makrofager. Legumain er i stand til å aktivere eller prosessere andre proteaser som prommp-2, cathepsin B, H og L, samt direkte bryte ned fibronektin som er en av hovedbestandelene av ekstracellulært matriks. Det antas dermed at legumain spiller en rolle ved utviklingen av aterosklerose og destabilisering av plakk. Det er også vist at behandling med statiner fører til nedregulering av legumain i sirkulerende monocytter. I denne studien har THP-1 monocytter blitt differensiert til M1- og M2-makrofager med lav (5 ng/ml) eller høy (62 ng/ml) konsentrasjon av forbolesteren PMA etterfulgt av proinflammatorisk (IFNγ og LPS) eller antiinflammatorisk (IL-4) stimuli for å gi opphav til henholdsvis M1 og M2. THP-1 cellene ble også behandlet med atorvastatin under differensieringen. Legumainuttrykk ble analysert i cellelysater ved immunoblotting, og enzymaktivitet ble målt med fluorescerende peptidsubstrat eller etter inkubering med fibronektin. Sekrert legumain ble målt ved ELISA i kondisjonerte medier. I denne studien ble det sett av M1- og M2-differensierte THP-1 makrofager uttrykte ulike former av legumain og at M1 sekrerte signifikant mer legumain enn M2. AEP-aktivitet i THP-1 makrofager var forårsaket av både legumain og ukjent(e) cysteincathepsin(er). Behandling med atorvastatin under differensieringen førte til nedregulering i uttrykket av moden legumain i M1, samt mindre prosessering av prolegumain og nedregulering av moden legumain i M2. VII
8 Abstract The cysteine protease legumain (asparaginylendopeptidase, AEP) is associated with several inflammatory diseases. In atherosclerosis, the expression and activity of legumain is increased in unstable regions of atherosclerotic plaques, when compared to stabile regions. Legumain is also localized to macrophages in atherosclerotic lesions, which indicates that this protease is primarily expressed by macrophages. Legumain is capable of activating or processing other proteases such as prommp-2, cathepsin B, H, L, as well as directly degrading fibronectin which is one of the constituents of extracellular matrix. It is thus assumed that legumain plays a role in atherogenesis and plaque destabilization. It has been demonstrated that treatment with statins leads to down regulation of legumain in circulating monocytes. In this study THP-1 monocytes have been differentiated to M1- and M2- macrophages with low (5 ng/ml) or high (62 ng/ml) concentrations of the phorbolic ester PMA, followed by pro-inflammatory or anti-inflammatory stimuli, giving rise to M1 and M2, respectively. The THP-1 cells were treated with atorvastatin during differentiation. Expression of legumain was analyzed in cell lysates with immunoblotting, and enzyme activity was measured with fluorescent peptide substrate or after incubation with fibronectin. Secreted legumain was measured with ELISA in conditioned media. In this study is was observed that M1- and M2-differentiated THP-1 macrophages expressed different forms of legumain, and that M1 secreted significantly more legumain than M2. The AEP-activity in THP-1 macrophages was due to legumain in addition to unknown cysteine cathepsin(s). M1/M2-differentiation with concomitant atorvastatin treatment caused a down regulation in the expression of mature legumain in M1, and less processing of prolegumain as well as a down regulation of mature legumain in M2. VIII
9 Innholdsfortegnelse Forord... V Sammendrag... VII Abstract... VIII Innholdsfortegnelse... IX Forkortelsesliste... XI 1 Innledning Proteaser Legumain Vevsuttrykk og lokalisering av legumain Aktivering og prosessering av legumain Fysiologiske funksjoner og substrater for legumain Legumain ved sykdomstilstander Statiners pleiotrope effekter Cysteinproteasehemmere Selektiv aktivitetsbasert probe og hemmer av legumain Makrofager M1-makrofager M2-makrofager Makrofagdifferensiering Mål med oppgaven Materialer og metoder Kjemikalier og reagenser Utstyrsliste Celledyrking THP-1 celler Tining, utsåing, splitting og telling av celler Høsting av kondisjonerte medier Høsting av cellelysater Differensiering av THP-1 monocytter til M1- og M2-makrofager med lav konsentrasjon PMA (metode 1) Differensiering av THP-1 monocytter til M1- og M2-makrofager med høy konsentrasjon PMA (metode 2) Inkubering av cellelysater med MP-L Behandling av celler med atorvastatin Totalproteinmåling Måling av asparaginylendopeptidase (AEP)-aktivitet Måling av legumainaktivitet Acetonfelling Elektroforese og immunoblotting Reblotting Enzyme-linked immunosorbent assay (ELISA) IX
10 3.10 Statistiske analyser Resultater Differensiering av THP-1 monocytter til M1- og M2- makrofager Legumain uttrykkes forskjellig i M1- og M2-makrofager differensiert med lav konsentrasjon PMA (5 ng/ml) Ulik morfologi av M1- og M2-makrofager Legumain uttrykkes forskjellig i M1- og M2-makrofager behandlet med høy konsentrasjon av PMA (62 ng/ml) Høyere sekresjon av legumain fra M- enn M2-makrofager Degradering av fibronektin Inkubering av M1- og M2-lysater med aktivitetsbasert legumainprobe Effekt av atorvastatin på M1- og M2-makrofager Økende konsentrasjoner atorvastatin hemmer adherens av M1- og M2-makrofager Atorvastatin førte til endret aktivitet, prosessering og sekresjon av legumain i M1- og M2-makrofager Atorvastatin hemmer prosessering av cathepsin L i makrofager Atorvastatin hemmer sekresjon av legumain fra M1-makrofager AEP-aktivitet versus legumainaktivitet i M1- og M2-makrofager behandlet med atorvastatin Diskusjon Konklusjon Litteraturliste Vedlegg A: Oppskrift for løsninger Vedlegg B: Foredrag-Norsk Selskap for Farmakologi og Toksikologi (NSFT) Vedlegg C: Posterpresentasjon-Gordon Research Seminar X
11 Forkortelsesliste AEP: Asparaginylendopeptidase Asn: Asparagin Asp: Aspartat CCL: Kjemokin (C-C motiv) ligand CD: Cluster of differentiation CEA: Karotis-endarektomi CXCL: Kjemokin (C-X-C) ligand DMSO: Dimetylsulfoksid DTT: Ditiotreitol E-64: Trans-epoksysuksinyl-L-leucylamido-(4-guanidino)butan ECM: Ekstracellulært matriks ELISA: Enzyme-linked immunosorbent assay enos: Endotel NO syntase FPP: Isoprenoidene farnesyl pyrofosfat GAPDH: Glyseraldehyd 3-fosfat dehydrogenase GFR: Glomerulær filtrasjonsrate GM-CSF: Granulocyttmakrofagkoloni-stimulerende faktor GPP: Geranylgeranyl pyrofosfat HMG-CoA: 3-hydrosky-3-metyl-glutaryl-koenzym HRP: Horseradish Peroxidase XI
12 IFNγ: Interferon gamma IL: Interleukin IL-1β: Interleukin-1 beta K i: Inhibisjonskonstant KO: Knock-out LDL: Low-density lipoprotein LDLR: Low-density lipoprotein receptor LPS: Lipopolysakkarid M4C: Stabiltransfekterte HEK293 cellelinje som overuttrykker Cystatin E/M M38L: stabiltransfektert HEK293 cellelinje som overuttrykker legumain M-CSF: Makrofagkoloni-stimulerende faktor MHC-II klasse II: Major histocompatibility complex MMP: Matriksmetalloproteinase mrna: Budbringer-ribonukleinsyre NF-κB: Nuclear factor kappa-light-chain-enhancer of activated B cells NK: Naturlige dreperceller PBS: Fosfatbufret saltvann PMA: phorbol 12-myristate 13-acetate prommp: Promatriksmetalloproteinase RGD: Arginylglysylaspartat ROS: Reaktive oksygenspesier XII
13 Rpm: Revolutions per minute, omdreininger per minutt RPMI medium: Roswell Park Memorial Institute medium SDS-PAGE: Natriumdodesylsulfat-polyakrylamidgelektroforese TGFβ: Transforming growth factor beta, transformerende vekstfaktor beta TLR: Toll-like receptor TNFα: Tumor-nekrosefaktoralfa TREM2: Triggering receptor expressed on myeloid cells 2 T-TBS: Trisbufret saltvann med Tween Z-Ala-Ala-Asn-AMC: Benzyloksykarbonyl-alanin-alanin-asparagin-7-amino-4- metylkumarin XIII
14 1 Innledning 1.1 Proteaser Proteaser (også kalt peptidaser eller proteinaser) er proteolytiske enzymer som katalyser spaltingen av peptidbindinger i proteiner ved hydrolyse, og spiller en viktig rolle i mange fysiologiske prosesser [1]. I databasen MEROPS er proteaser inndelt hierarkisk og gruppert sammen i seks ulike klasser etter deres katalytiske sete; aspartat-, cystein-, glutamat-, metallo-, serin- og treoninproteaser (Figur 1.1). Videre er disse igjen delt inn i klaner basert på deres likhet i tertiærstruktur. Det antas at proteaser i samme klan har felles evolusjonært opphav. Proteaser med signifikant likhet i aminosyresekvensen grupperes i familier. Dermed kan en klan bestå av flere familier av proteaser [2]. Proteaser, avhengig av deres spaltningsposisjon i polypeptidkjeden, betegnes som enten eksopeptidaser eller endopeptidaser. Eksopeptidaser spalter i enden av polypeptidkjeden fra N-terminal eller C- terminal ende (aminopeptidaser eller karboksypeptidaser), mens endopeptidaser angriper innad i kjeden [1]. Figur 1.1.: MEROPS klassifisering av proteaser i pattedyr. Proteaser klassifiseres i klasser (blå bokser) etter dere katalytiske sete og klassene er videre delt inn i klaner (oransje bokser) etter deres tertiærstruktur. Proteaser deles videre inn i familier (røde bokser) basert på likhet i aminosyresekvens. Figuren er modifisert fra Rawlings et al. (2016) [2] 1
15 1.1.1 Legumain Legumain er en asparginyl endopeptidase (AEP) som tilhører C13-familien innenfor klan CD av cysteinproteaser [2] (Figur 1.1). Den ble først oppdaget i plantefrø fra Vicia sativa og senere i parasitten Schitosoma mansoni [3, 4]. Først i 1997 ble legumain karakterisert i pattedyr [5]. Legumain er strukturelt beslektet med caspasene (C14)[6]. I likhet med andre familier innenfor CD-klanen viser legumain et strengt krav for aminosyren i P1-posisjonen for at spalting skal kunne skje, og spalter spesifikt i peptidsubstrater karboksyterminalt for asparagin (Asn) [7]. Det er derimot vist at legumain også kan spalte etter aspartat (Asp) ved sure betingelser. Aspartat har en pka på ca. 4 og vil ved ph 3 være i protonert form og strukturelt ligne asparagin og derved gjenkjennes av legumain. [8]. 1.2 Vevsuttrykk og lokalisering av legumain I pattedyr er uttrykket av legumain høyest i nyrer, milt, lever og testikler [5, 9]. Legumain er også detektert i human placenta [5]. I pattedyr er legumain hovedsakelig lokalisert intracellulært i sene endosomer og lysosomer, men tilstedeværelse av proteolytisk aktiv legumain har også blitt sett både i cellekjernen og ekstracellulært [10, 11]. 1.3 Aktivering og prosessering av legumain Legumain uttrykkes som et inaktivt proenzym og gjennomgår ph-avhengig autoaktivering og omdanning til aktiv endopeptidase [12]. Dette proenzymet, kalt prolegumain (56 kda), består av et katalytisk domene og et C-terminalt prodomene. Det C-terminale prodomenet består av et aktiveringspeptid (AP, Lys 287 -Asn 323 ; Figur 1.2, blått) og et legumain stabilisering- og aktivitetsmoduleringsdomene (LSAM; Figur 1.2, oransje) [7]. I det sure miljøet i lysosomer autoaktiveres prolegumain til en 47 kda inaktiv og en aktiv mellomform på 46 kda ved autoprosessering på C- og N-terminal ende (Figur 1.2.). Videre omdanning til den modne aktive 36 kda-formen er ikke autokatalytisk, men det er ikke kjent hvilke(n) protease(r) som er involvert i denne omdannelsen [12]. Omdanning av prolegumain til aktiv form medfører endring i ph-stabilitet av proteasen. Mens prolegumain er stabil ved nøytral ph, vil moden legumain denatureres ved ph > 6. Legumain har et Arg-Gly-Asp 120 (RGD) motiv som kan binde til αvβ3-integrinreseptoren på cellemembranen. Ved binding til denne reseptoren øker 2
![legumain sitt ph optimum fra 5,5 til 6 [13]. Prolegumain Mellomform Moden legumain Ukjent protease Figur 1.2.: Prosessering og aktivering av legumain.](/docs-images/63/49456397/images/16-0.jpg "Dannelse av moden legumain (36 kda) fra prolegumain (56 kda) skjer via flere trinn.")
16 legumain sitt ph optimum fra 5,5 til 6 [13]. Prolegumain Mellomform Moden legumain Ukjent protease Figur 1.2.: Prosessering og aktivering av legumain. Dannelse av moden legumain (36 kda) fra prolegumain (56 kda) skjer via flere trinn. Ved sur ph gjennomgår legumain autokatalytisk prosessering til inaktiv 47 kda-form (ph 5) og aktiv 46 kda-form (ph 4,5). Videre omdannelse til moden aktiv 36 kda legumain foregår ikke via autokatalyse, men utføres av en foreløpig ukjent protease. Figuren er modifisert fra Dall og Brandstetter (2016) [7]. Endogene hemmere av legumain er cystatin C, cystatin E/M og cystatin F [14]. Av disse er cystatin E/M den mest potente. Derimot så hemmes ikke legumain av E-64, som er en generell hemmer av cysteinproteaser som tilhører C1-familien (Figur 1.1) [15]. 1.4 Fysiologiske funksjoner og substrater for legumain Legumain spiller blant annet roller i antigenprosessering og immunsignalering. Det har blitt rapportert at legumain er involvert i prosessering av tetanus C-fragment for presentering til MHC-II (klasse II major histocompatibility complex ) [16]. I studier av dendrittiske celler har det blitt vist at legumain spalter Toll-like reseptor 9 (TLR9), som fører til aktivering av denne reseptoren [17]. Det har blitt gjort studier av legumain knock-out (KO) mus. Legumain KO-mus var født fertile og uten morfologiske abnormaliteter, men musene hadde derimot en redusert veksthastighet [18]. I den samme studien ble det observert at KO-mus hadde forstørrede lysosomer og redusert prosessering av cysteincathepsinene B, H og L. Disse cathepsinene prosesseres normalt av legumain. En annen KO-studie på mus har vist at legumain spiller en viktig rolle i opprettholdelse av normal nyrefunksjon. Legumain KO-mus hadde redusert glomerulær filtrasjonsrate (GFR) og akkumulering av proteiner i proksimale nyretubuli [19]. 3
17 De lysosomale cysteinproteasene cathepsin B, H og L tilhører klan CA og er i C1- familien (Figur 1.1). Disse cathepsinene uttrykkes både som single-chain og two-chain proteolytiske aktive former etter omdannelse fra deres pro-former ved sur ph, og det er kjent at legumain tar del i prosessering av single-chain til two-chain -formen [18, 20]. Ved in vitro assay med cathepsin B, H og L er cathepsin L funnet å ha høyest protease-aktivitet [21]. Bestanddeler av ekstracellulær maktrisk (ECM) som elastin, kollagen og proteoglykan er identifisert som substrater for cathepsiner [20]. En annen gruppe proteolytisk enzymer som kan degradere ECM er matriksmetalloproteinaser (MMPs) [22], og det er vist at legumain er i stand til å aktivere prommp-2 til aktiv MMP-2 in vitro [23]. I tillegg til (pro)legumain, tetanus C-fragment, TLR9, cysteincathepsin B, H og L og prommp-2 som er nevnt over, finnes det noen andre kjente substrater for legumain, inkludert fibronektin. Fibronektin er et høymolekylært glykoprotein som foreligger som en dimer bestående av to sub-enheter på 250 kda som er bundet sammen ved en sulfid-bro. Fibronektin er en av hovedbestanddelene av ekstracellulær matriks (ECM) og har et RGDmotiv som gjør at det kan binde til integriner [24]. Fibronektin er også i stand til å binde til flere komponenter av ECM som fibrin, heparan sulfat proteoglykan og kollagen [25]. I nyrer fra legumain KO-mus er det en akkumulering av fibronektin i det tubulointerstitiale område, og det er i tillegg vist at legumain kan spalte fibronektin in vitro [26]. Ved å aktivere prommp-2 og spalte fibronektin er det mulig at legumain kan regulere degraderingen av ECM. 1.5 Legumain ved sykdomstilstander Foruten sin rolle i normalfysiologi er legumain assosiert med flere inflammatoriske sykdomstilstander som aterosklerose og kreft [27-29]. Legumain er overuttrykt i flere kreftsvulster og er assosiert med økt invasjonsevne og metastasering [30]. I tillegg til å være uttrykt i tumorceller, er legumain funnet i omkringliggende stromaceller og er høyt uttrykt i tumor-assosierte makrofager (TAMs), samt ekstracellulært i mikromiljøet rundt svulster [31, 32]. 4
18 Ved aterosklerose er både uttrykk og aktivitet av legumain høyere i ustabile områder av aterosklerotiske plakk, sammenlignet med stabile områder [27, 33]. Legumain aktiverer andre proteaser som prommp-2, cathepsin B, H og L [18, 33], som bidrar i nedbrytning av ekstracellulær matriks (ECM). [26]. Ved direkte å bryte ned fibronektin og ved å aktivere andre proteaser kan legumain lede til plakkdestabilisering [26, 27]. Det er vist at legumain er først og fremst lokalisert til makrofager i aterosklerotiske plakk, noe som indikerer at enzymet hovedsakelig uttrykkes der [28, 33]. 1.6 Statiners pleiotrope effekter Statiner er kolesterolsenkende legemidler som benyttes i behandling av hyperkolesterolemi og for å motvirke hjerte/kar-sykdom. Statiner hemmer det hastighetsbestemmende enzymet i biosyntesen av kolesterol, HMG-CoA reduktase, som katalyserer omdannelsen av HMG-CoA til mevalonsyre [34]. I tillegg til å redusere de novo biosyntese av kolesterol har statiner kolesterolsenkende effekt ved å oppregulere uttrykket av low-density lipoprotein (LDL)- reseptor, hvilket fører til økt reabsorpsjon av kolesterol fra sirkulasjonen [34]. Flere store kliniske studier har indikert at statiner har flere gunstige effekter i forebyggingen av aterosklerose og koronare sykdommer som ikke er direkte relatert til deres effekter på lipidprofilen; såkalte kolesterol-uavhengige eller pleiotrope effekter [35]. Det er foreslått at noen av disse pleiotrope effektene kan skyldes at statiner modulerer endotelfunksjon, reduserer reaktive oksygenspesier (ROS) og har antiinflammatoriske og plakkstabiliserende effekter [34]. Ved å hemme enzymet HMG-CoA reduktase fører statiner også til en redusert syntese av isoprenoidene farnesyl pyrofosfat (FPP) og geranylgeranyl pyrofosfat (GPP). FPP og GPP er intermediære i kolesterolbiosyntesen og fungerer som lipidvedheng som regulerer den kovalente tilkoblingen av en farnesylenhet eller geranylgeranylenhet til målproteiner. Tilkobling av disse enhetene til målproteiner (isoprenylering) har betydning for forankring, subcellulær lokalisering og transport av membran-assosierte proteiner [34, 36]. Viktige substrater for isoprenylering er medlemmer av Ras og Rho familien av GTPaser og det er vist at isoprenylerte Rho proteiner nedregulerer uttrykk av endotel NO syntase (enos). Da statiner fører til en redusert syntese av isoprenoider er det foreslått at statiner kan motvirke Rho proteiner og gi en oppregulering av endotel NO-syntase (enos), [34, 36]. Det er også 5
19 vist at statiner er i stand til å hemme aktivering av den inflammatoriske transkripsjonsfaktoren nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) i kultur [37]. Effekt av statiner på genuttrykk i monocytter/makrofager har blitt undersøkt i flere studier. I monocytter isolert fra pasienter behandlet med atorvastatin er det sett en reduksjon i legumain mrna nivået [38]. 1.7 Cysteinproteasehemmere Endogene hemmere av cysteinproteaser er cystatinene, og de inndeles i 3 typer. Cystatiner er kompetitive, reversible hemmere [39]. Type 1 cystatiner er cystatin A og B, som mangler et signalpeptid og er normalt intracellulære [39]. Blant type 2 cystatinene er cystatin C, E/M og F, og det er kun cystatiner i denne gruppen som hemmer legumain. Cystatin E/M er den mest potente hemmeren av legumain med en K i på 0,0016 nm, mens cystatin C og F har K i på henholdsvis 0.2 nm og 19 nm [11]. Type 3 cystatiner er kininogenene som, i tillegg til å være forløpere til kininer, også er hemmere av cysteinproteaser. Type 3 cystatiner finnes hovedsakelig i blod og synovialvæske [39]. Cystatin E/M er en potensiell tumorsupressor og er vist å hemme invasjon av human melanoma og orale carinoma celler [11]. Cystatin E/M er hovedsakelig sekretorisk, men er vist til å kunne internalisere og hemme legumain intracellulært [11]. E-64 er et epoksid som først ble isolert fra Aspergillus japonicus og vist å være en irreversibel hemmer av cysteincathepsiner [15, 40], men hemmer ikke legumain [5, 14] Selektiv aktivitetsbasert probe og hemmer av legumain Nylig har det blitt syntetisert og karakterisert selektive aktivitetsbaserte prober for legumain [41] deriblant MP-L01 som er brukt i denne oppgaven. Denne proben er merket med biotin (Figur 1.3, Tag ), og dette gjør det mulig å detektere binding av probe til legumain ved bruk av streptavidin. Proben har en elektrofil acyloksymetylgruppe som danner en kovalent, irreversibel binding til cystein i det aktive setet (AOMK; Figur 1.3., Warhead ). Peptidmotivet i P4-P1 ble selektert etter screening av et aminosyrebibliotek bestående av en blanding av naturlige og unaturlige aminosyrer i P4-P2. Aminosyren i P1-posisjon er ikke 6
20 asparagin (Asn), men aspartat (Asp). Ved sur ph protoneres Asp og blir funksjonelt likt asparagin (Asn), som er den foretrukne aminosyren i P1-posisjon for legumain [8]. I tillegg ble probe uten biotinmerking (MP-L01*; Poreba, Marcin, Wroclaw University, Polen) brukt i denne oppgaven. MP-L01 er vist å være selektiv for binding til aktiv legumain (36 kda og 46 kda) og ikke prolegumain (56 kda), samt at proben tas opp og inhiberer legumain i levende celler [41]. A B Figur 1.3.: Generell struktur av selektiv aktivitetsbasert MP-L01 probe for legumain. A, Biotinmerking (rødt). Ahx=aminokapronsyre, spacer. P4-P2=naturlige og unaturlige aminosyrer. P1(grønt)=Aspartat. AOMK (blått)=acyloksymetylgruppe som danner en kovalent, irreversibel binding til cystein i det aktive setet. B, Strukturformel for MP-L Makrofager Makrofager spiller en sentral rolle i immunrespons og opprettholdelse av homeostase og finnes i alle organer og vev. De er en del av det mononukleære fagocytt-systemet som inkluderer forløperceller til monocytter, sirkulerende monocytter, dendrittiske celler og vevsmakrofager. Sirkulerende monocytter differensieres til makrofager etter proinflammatorisk, metabolske eller immunologiske stimuli [42, 43]. Makrofager har flere 7
21 funksjoner, blant annet fagocytose av apoptopiske celler og cellerester og antigenpresentasjon til T-celler [44, 45]. Makrofager er en heterogen gruppe celler, med ulike funksjonelle fenotyper. Hvilken fenotype makrofager differensieres til avhenger av lokale stimuli i mikromiljøet [46, 47]. I litteraturen klassifiseres makrofager som M1 eller M2, eller henholdsvis klassisk aktiverte makrofager (M1) eller alternativt aktiverte makrofager (M2). Betegnelsen av makrofager som M1 eller M2 oppstod for å reflektere hvilken T-hjelpecelle (T-hjelpecelle 1 (Th1) eller (Th2)) som bidrar i makrofagdifferensieringen. M1-makrofager utviser en mer proinflammatorisk fenotype, mens M2-makrofager har en regulatorisk funksjon i vevsreparasjon og remodellering [46-48] M1-makrofager M1-stimuli gruppes etter deres evne til å igangsette inflammatoriske responser i makrofager, og IFNγ er det cytokinet som hovedsakelig er assosiert med aktivering av makrofager til en M1-fenotype. IFNγ produseres hovedsakelig av Th1, men også av naturlige dreperceller (NK) og andre makrofager. Denne M1-fenotypen kan også oppnås i kombinasjon med andre cytokiner som tumor-nekrosefaktoralfa (TNFα), granulocyttmakrofagkoloni-stimulerende faktor (GM-CSF) eller lipopolysakkarid (LPS). Etter M1-stimuli utskiller makrofager proinflammatoriske cytokiner som IL-1, IL-6, IL-12 [46, 49] M2-makrofager Blant makrofager som betegnes som M2 finnes det i dag mange undergrupper. M2- makrofager ble opprinnelig karakterisert som en fenotype som oppstår etter stimulering av makrofager med IL-4 og IL-13, men i dag så er flere andre cytokiner og signalstoffer anerkjent som igangsettere av M2-differensiering, deriblant IL-10, TGFβ, M-CSF og glukokortikoider [44, 46, 49] Makrofagdifferensiering For å igangsette differensiering av makrofager til M1- eller M2-fenotyper in vitro benyttes det flere metoder [50]. Med hensyn til metoder for å oppnå M1- og M2-makrofager er det variasjon i type stimuli, varighet av stimulering, konsentrasjon av stimuli og celletyper som 8
22 er brukt. Eksempelvis har det blitt benyttet primære monocytter isolert fra humant blod [50, 51] og en human monocyttisk cellelinje (THP-1) [52]. 9
23 2 Mål med oppgaven Etablere en metode i laboratoriet differensiering av THP-1 monocytter til henholdsvis M1- og M2-makrofager. Undersøke uttrykk, aktivitet og sekresjon av legumain i M1- og M2-makrofager. Undersøke hvordan uttrykk, aktivitet og sekresjon av legumain påvirkes etter behandling av M1- og M2-makrofager med atorvastatin under differensieringen. 10
24 3 Materialer og metoder 3.1 Kjemikalier og reagenser Kjemikalier Albuminstandard (23209) Aceton Atorvastatin (lakton-form) (A791780) Bio-Rad Protein Assay Dye reagent Concentrate ( ) Renset bovint legumain fra storfenyre CHAPS, C 32 H 58 N 2 O 7 S (C3023-IG) Cystatin E/M (fra M4C-kondisjonert medium Dinatrium-EDTA, NA 2 C 10 H 16 N2O 8 DMSO, dimetylsulfoksid, (CH 3 ) 2 SO (D2650) DTT, Ditiotreitol, C 4 H 10 O 2 S 2 (438117) E-64, Trans-epoksysuksinyl-L-leucylamido- (4-guanidino)butan FBS, føtalt kalveserum, fetal bovine serum (A15-151) Fibronektin fra human plasma (F2006) Geit anti-human polyklonalt antistoff mot legumain (AF2199) Geit anti-human polyklonalt antistoff mot CD163 (AF1607) Geit anti-human polyklonalt antistoff mot CD206 (AF2534) Geit anti-human polyklonalt antistoff mot Cathepsin L (MAB952) Leverandør Thermo Scientific, Rockford, IL, USA Merck, Darmstadt, Tyskland Toronto Research Chemicals Inc., Toronto, ON, Kanada Bio-Rad Laboratories, Hercules, CA, USA Johansen, HT, seksjon for farmasøytisk biovitenskap, Universitetet i Oslo Sigma-Aldrich, St. Louis, MI, USA Lunde, NN, seksjon for farmasøytisk biovitenskap, Universitetet i Oslo Ferak, Berlin, Tyskland Sigma-Aldrich Sigma-Aldrich Sigma-Aldrich PAA Laboratories, Pasching, Australia Sigma Aldrich R&D Systems, Minneapolis, MN, USA R&D Systems, Minneapolis, MN, USA Santa Cruz Biotechnology, TX, USA 11
25 Geit antihuman polyklonalt antistoff mot CD80 (AF140) Geit anti-mus polyklonalt antistoff mot GAPDH (Sc-47724) Hydrogenklorid, HCl (26614) IRDye 680LT, Esel anti-geit immunoglobulin ( ) Sigma-Aldrich Licor Biosciences, Cambridge, Storbritannia IRDye 800CW, Streptavidin ( ) Kanin anti-geit polyklonalt antistoff immunoglobulin (PO160) LPS fra Escherichia coli (L5418) M38L-lysat Mus anti-human monoklonalt antistoff mot fibronektin (MAB1918) Natriumdihydrogenfosfat, NaH 2 PO 4 H 2 O (6346) Natriumhydroksid, NaOH (6495) Natriumpyrovatløsning, 100 mm, C 3 H 3 NaO 3 (S11-003) Natriumklorid, NaCl ( ) Natriumcitrat, (Na 3 C 6 H 5 O 7 2H 2 O) NuPAGE Antioxidant NuPAGE LDS Sample buffer (4X) NuPAGE MOPS Running buffer (20X) (NP0001) Penicillin/Streptomycin ( U/ml) (P11-010) PMA, Forbol 12-myristat 13-acetat, 12-Otetradekanoylphorbol 13-acetate (P1585) Ponceau-løsning (P7170IL Precision Plus Protein Dual Color Standards ( ) Rekombinant human IFNγ (285-IF) Dako Cytomation, Glostrup, Danmark Sigma-Aldrich Lunde, NN, seksjon for farmasøytisk biovitenskap, Universitetet i Oslo R&D Systems Merck Merck PAA Laboratories Merck Merck Life Technologies Life Technologies PAA Laboratories Sigma-Aldrich Sigma-Aldrich Bio-Rad Laboratories R&D Systems 12
26 Rekombinant human IL-4 (204-IL) Restore Western Blot Stripping Buffer (21059) Substrate Reagent Pack (DY999) Reagent dilutent concentrate (10X) (DY995) DuoSet Human total legumain (DY4769) Tris, C 4 H 11 NO 3 (T1506) Tris/Glycin buffer (10X) Tryptanblått 0,4 % Tween 20 ( ) Tørrmelk Z-Ala-Ala-Asn-AMC, legumainsubstrat Thermo Scientific R&D Systems Sigma-Aldrich Bio-Rad Laboratories Life Technologies Bio-Rad Laboratories Normilk, Levanger, Norge Bachem, Budendorf, Sveits 3.2 Utstyrsliste Utstyr BioHit Optifit tip pipettespisser (10, 200, 1000 μl) Corning 75 cm 2 dyrkningsflaske (3275) Corning Sentristar, sentrifugerør (15 ml) (430791) Corning Sentristar, sentrifugerør (50 ml) (430921) Costar 6-brønnersbrett (3506) Costar 96-brønners sort mikrotiterplate (3915) Costar 5 ml pipette (4051) Costar 10 ml pipette (4101) Costar 25 ml pipette (4251) Countess Automated Cell Counter Dri-Black DM-2A ECL Semi-dry Blotter Leverandør Sartorius Biohit Liquid Handling Oy, Finland Corning Incorporated, NY, USA Invitrogen, Life Technologies Techne, Cambridge, Storbritannia Amersham Biosciences, Buckinghamshire, 13
27 Eppendorfrør (0,5, 1.5, 2 og 5 ml) Heraeus Fresco 21 Centrifuge ( ) Holten Lamin Air LAF-benk Inkubatorskap (Galaxy 170S) Melocular Imager ChemiDoc LabTM XRS + (721BR03414) Nunc 96-brønners blank mikrotiterplate (260836) Nitrocellulosemembran, Hybond-ECL- Amersham NuPAGE 4-12 % Bis-Tris Protein Gels Novex (NP0322PK) Odyssey CLx Near-Infrared Imaging System Olympus CKX41 lysmikroskop SUB Aqua vannbad Victor X4 Multimode Plate Reader, mikroplateleser XCell SureLock Electrophoresis Cell Storbritannia Eppendorf AG, Hamburg, Tyskland Thermo Scientific Medinor, Oslo, Norge Eppendorf, NY, USA Bio-Rad Laboratories Nunc, Roskilde, Danmark Ge Healthcare Bio-Sciences, Oslo, Norge Life Technologies, Carlsbad, CA, USA Li-Cor Biosciences Olympus, Tokyo, Japan Grant, Cambridge, England Perkin Elmer Life and Analytical Sciences, Boston, MA, USA Life Technologies 14
28 3.3 Celledyrking THP-1 celler THP-1 celler (ATCC; TIB-202) er en monocyttcellelinje isolert fra blodet til en ett år gammel gutt med akutt monocyttisk leukemi [53]. Denne cellelinjen har blitt brukt til å studere blant annet funksjoner, mekanismer og signaleringsveier i monocytter og makrofager [54] og for å differensiere THP-1 celler til M1- og M2-makrofager [55] Tining, utsåing, splitting og telling av celler Cellene ble tatt ut av nitrogentanken, der de ble oppbevart ved -192 C, og tint i sekunder på vannbad ved 37 C under konstant bevegelse. Cellene ble tint til det var en liten isklump igjen. Deretter ble cellematerialet overført til en celleflaske (10 ml) som inneholdt 9 ml THP-1 dyrkningsmedium (se under), som på forhånd var varmet opp til 37 C. Cellene ble så dyrket ved 37 C og 5 % CO 2 over natt. Cellene ble så sentrifugert i 5 minutter ved 800 rpm før medium ble sugd av og nytt medium tilsatt. Cellene ble splittet når konsentrasjonen i celleflasken var nær 1 mill./ml, som var omtrent en gang i uken. En million celler ble sentrifugert i 5 minutter ved 800 rpm. Cellepelleten ble deretter resuspendert i 10 ml THP-1 dyrkningsmedium og overført til ny celleflaske. THP-1 celler ble telt ved å blande 20 μl trypanblått med 20 μl cellesuspensjon. Etter blandingen av trypanblått til cellesuspensjon ble 10 μl overført til et tellekammer og telt i en automatisk celleteller (Countess TM ). THP-1 dyrkningsmedium som ble benyttet bestod av RPMI 1640 medium med 2 mm L- glutamain og 1,5 g/l natriumbikarbonat, 10 % føtalt kalveserum, 4,5 g/l glukose, 10 mm HEPES, 1 mm natriumpyruvat, 0,05 mm 2-mercaptoetanol, 100 U/ml penicillin, 100 µg/ml streptomycin og 2,5 µg/ml Fungizone (Amphoterricin). 15
29 3.3.3 Høsting av kondisjonerte medier Høsting av kondisjonerte medier ble gjort ved å forsiktig avpipettere og sentrifugere mediene ved 800 rpm i 5 minutter. Deretter ble supernatanten overført til nytt eppendorfrør og oppbevart ved -70 C før videre analyser Høsting av cellelysater For høsting av cellelysater ble cellene vasket forsiktig med 1 ml PBS-løsning (vedlegg A 1.1). PBS ble tilsatt forsiktig langs veggen av brønnen og deretter fjernet. THP-1 celler behandlet med lav konsentrasjon PMA (5 ng/ml) ble tilsatt 500 µl lysisbuffer (vedlegg A 1.2) og THP-1 celler behandlet med høy konsentrasjon PMA (62 ng/ml) ble tilsatt 250 µl lysisbuffer. Cellene ble skylt med PBS og skrapet løse før lysatet ble overført til eppendorfrør. Deretter ble cellelysatene frosset ved -70 C og tint opp på varmeblokk ved 30 C. Dette ble gjentatt tre ganger for å lysere cellene fullstendig. Etter denne prosessen ble lysatene sentrifugert ved x g i 10 minutter ved 4 C. Supernatanten ble deretter pipettert over i nytt eppendorfrør. Cellelysater ble oppbevart ved -70 C før videre analyser Differensiering av THP-1 monocytter til M1- og M2-makrofager med lav konsentrasjon PMA (metode 1) Etter telling av THP-1 celler i celleflasker ved hjelp av automatisk celleteller, ble ønsket volum av cellesuspensjon sentrifugert i 5 minutter ved 800 rpm. Supernatanten ble så fjernet og cellepelleten ble resuspendert i THP-1 dyrkningsmedium til en ønsket cellekonsentrasjon (f. eks. 0,5 mill./ml). Cellesuspensjonen ble tilsatt en hjelpeløsning av PMA (1 µg/ml) som ga sluttkonsentrasjon på 5 ng/ml. Denne hjelpeløsningen ble laget ved å fortynne stamløsning (100 µg/ml) med dyrkningsmedium. Celler ble så sådd ut i 6-brønners brett (2 ml/brønn) og inkubert ved 37 C og 5 % CO 2 i 48 timer. Medium ble fjernet og hver brønn ble vasket forsiktig med 1 ml dyrkningsmedium. For M1-differensiering ble 2 ml dyrkningsmedium med LPS (1 µg/ml) og IFNγ (20 ng/ml) tilsatt og for M2-differensiering ble 2 ml dyrkningsmedium med IL-4 (20 ng/ml) tilsatt. Til kontroll ble 2 ml dyrkningsmedium tilsatt. Etter 72 timer ble alle brønner inspisert for adherens før høsting. 16
30 3.3.6 Differensiering av THP-1 monocytter til M1- og M2-makrofager med høy konsentrasjon PMA (metode 2) THP-1 celler i celleflasker ble telt og ønsket volum ble sentrifugert i 5 minutter ved 800 rpm. Supernatanten ble fjernet og cellepellet ble resuspendert i THP-1 dyrkningsmedium til ønsket cellekonsentrasjon (0,5 mill./ml) og tilsatt PMA (100 µg/ml) til en sluttkonsentrasjon på 62 ng/ml. Celler ble så sådd ut i 6-brønners brett (2 ml/brønn) og inkubert ved 37 C og 5 % CO 2 i 6 timer før videre stimulering. For M1-differensiering ble 2 ml dyrkingsmedium med LPS (10 ng/ml) og IFNγ (5 ng/ml) tilsatt og for M2-differensiering ble 2 ml dyrkningsmedium med IL-4 (25 ng/ml) tilsatt. Til kontroll ble 2 ml dyrkningsmedium tilsatt. Etter 72 timer ble alle brønner inspisert for adherens før høsting Inkubering av cellelysater med MP-L01 Cellelysater av M1- og M2-makrofager (3.3.4) ble tilsatt en syntetisk irreversibel hemmer og aktivitetsbasert probe for legumain (MP-L01) [41] og inkubatene analysert med Odyssey CLx Near-Infrared Imaging System etter gelelektroforese og immunoblotting. M1- og M2-cellelysater (19 µl) ble tilsatt 1 µl 20 µm MP-L01 (se under). Lysat fra legumainoveruttrykende M38L celler [11] og renset bovint legumain ble brukt som kontroller på binding av MP-L01 til legumain. Til 19 µl M38L-lysat og 19 µl renset bovint legumain (36,2 µm) ble også tilsatt 1 µl 20 µm MP-L01, slik at sluttkonsentrasjon av MP-L01 i inkubatene var 1 µm. Disse blandingene ble inkubert i 30 minutter ved 30 C. MP-L01 (20 µm) ble laget ved å fortynne 1 µl av stamløsning på 10 mm i 500 µl i legumainassaybuffer. Til prøver uten probe ble det tilsatt 0,2 % DMSO. Stamløsning av renset bovint legumain var på 275 µm og 2,5 µl bovint renset legumain (275 µm) ble fortynnet til totalt 19 µl med assaybuffer til en konsentrasjon på 36 µm før tilsetning av MP- L Behandling av celler med atorvastatin THP-1 celler ble behandlet med atorvastatin etter PMA-behandling og rett etter tilsetning av LPS/IFNγ eller IL-4. Sluttkonsentrasjoner av atorvastatin i brønner var 0, 1, 5, 10, 25 og 50 µm. Det ble laget flere hjelpeløsninger av stamløsning (40 mm i DMSO) av atorvastatin for å oppnå disse sluttkonsentrasjonene. Dette ble gjort for å kunne tilsette likt volum av 17
31 atorvastatinfortynningene (10 µl) direkte i cellebrønnene. Til kontrollen (0 µm atorvastatin) ble det tilsatt 0,5 % DMSO. 3.4 Totalproteinmåling Kvantifisering av mengden totalprotein i cellelysatene ble gjort i henhold til metode beskrevet av Bradford [56]. Metoden baserer seg på at det skjer en endring i absorpsjonsmaksimum når fargestoffet Comassie Brilliant Blue G-250 bindes til protein. Ved binding av Comassie Brilliant Blue G-250 til protein vil det skje en fargeendring fra rødt til blått og absorpsjonsmaksimum skiftes fra 465 til 595 nm. Bio-Rad Protein Assay Dye Reagent Concentrate som inneholder fargestoffet ble fortynnet 1:5 i destillert vann. Albumin i konsentrasjoner 0, 50, 100, 200, 300 og 400 μg/ml ble brukt til å lage en standardkurve. Proteinkonsentrasjonene i lysatene ble beregnet ut fra denne standardkurven. Volum på 10 μl standarder og lysater ble tilsatt til en klar 96-brønners mikrotiterplate før tilsetting av 200 μl Bio-Rad Protein Assay Dye Reagent Concentrate -fortynning. Absorbans ved 595 nm ble målt i mikroplateleser (Wallac 1420 VictorX4 Multilabel counter) 5 minutter etter tilsetning av fargeløsningen. Både standarder og prøver ble målt som triplikater. 3.5 Måling av asparaginylendopeptidase (AEP)-aktivitet Metoden som er brukt for måling av AEP-aktivitet er basert på metode beskrevet av Chen et al. [5] og Johansen et al. [57]. For måling av AEP-aktivitet ble prøvematerialet fortynnet i legumainassaybuffer (vedlegg A 1.3). Denne bufferen inneholdt reduksjonsmiddelet DTT (ditiotreitol, 1 mm) slik at cysteinproteasene aktiveres. Det ble så tilsatt et fluorescerende peptidsubstrat, Z-Ala-Ala-Asn-AMC, som spaltes av asparaginylendopeptidaser. Ved spalting avgis leaving -gruppen AMC som har høyere fluorescens i fri form enn når det er bundet i peptidsubstratet. Økning i fluorescens etter spalting ble målt i en mikroplateleser (Wallac 1420 VictorX4 Multilabel counter). Lysisbuffer (vedlegg A 1.2) ble brukt som blindprøve. 18
32 Volum på 20 μl prøve eller blindprøve ble tilsatt en sort 96-brønners mikrotiterplate. Prøve og blindprøve ble tilsatt i triplikater og mikrotiterplaten ble så plassert i mikroplateleseren. Hver brønn ble automatisk tilsatt 100 μl legumainassaybuffer. Legumainsubstratløsning, 50 μl (34 μm; se vedlegg A 1.4) ble tilsatt etter 10 minutter for å sikre aktivering av cysteinproteaser. Mikrotiterplaten ble ristet i 3 sekunder etter tilsetning av legumainassaybuffer og substratløsning. Temperaturen ble holdt ved 30 C, da hastigheten av den enzymatiske reaksjonen er temperaturavhengig. Fluorescens ble målt totalt 10 ganger i løpet av 60 minutter. Målinger ble gjort umiddelbart etter tilsetning av substratløsning og repetert med mellomrom på seks minutter. Mikrotiterplaten ble ristet i 3 sekunder før hver måling. Endring i fluorescens per tidsenhet ( F/sek) ble så kalkulert automatisk i mikroplateleseren. 3.6 Måling av legumainaktivitet For å bestemme hvor mye av AEP-aktiviteten som skyldes legumainaktivitet i lysatene ble forbindelsen E-64 benyttet. E-64 er en potent irreversibel hemmer av cysteincathepsiner som ikke hemmer legumain [5]. E-64 ble tilsatt cellelysater før måling av AEP-aktivitet ved to enkle flourescensmålinger. Første måling ble gjort 90 minutter etter tilsetning av E-64 og siste måling ble gjort 6 timer etter tilsetning av E-64. Volum på 20 μl prøve og blindprøve (lysisbuffer) ble pipettert i en sort 96-brønners mikrotiterplate i duplikater. Hver brønn ble tilsatt 17 μl 10 μm E-64 (vedlegg A 1.5) eller 17 μl vann. Sluttkonsentrasjon av E-64 i hver brønn var 1 μm. Platen ble deretter inkubert ved 37 C i 90 minutter og så satt i mikroplateleser (Wallac 1420 VictorX4 Multilabel counter). Legumainassaybuffer (83 μl) ble tilsatt fra autoinjektor i hver brønn etterfulgt av 50 μl tilsetning av substratoppløsning (34 μm). Platen ble ristet i 10 sekunder og fluorescens målt. Etter første måling (90 min) ble platen igjen inkubert i 4,5 timer frem til ny måling ble gjort etter totalt 6 timer. Legumainaktivitet ble kalkulert ved å subtrahere målt fluorescens ved 1,5 time fra målt fluorescens ved 6 timer. 19
33 3.7 Acetonfelling Totalproteinkonsentrasjonene i cellelysatene var relativt lave og det var derfor nødvendig å oppkonsentrere cellelysatene før elektroforese kunne utføres (kapitel 3.6). Først ble mengde cellelysat som inneholdt ønsket mengde totalprotein beregnet og overført til merkede eppendorfrør. Til denne mengden cellelysat ble det tilsatt fire ganger så stor mengde iskald aceton (100 %). Innholdet ble blandet godt og inkubert i 1 time ved -20 C. Etter inkubering ble rørene sentrifugert i 20 minutter ved G og 4 C, hvorpå supernatanten ble forsiktig avpipettert. Pelleten ble så vasket med 1 ml aceton (100 %) og deretter sentrifugert i 20 minutter ved G og 4 C. Tilslutt ble proteinpelleten tørket i ca minutter ved romtemperatur. 3.8 Elektroforese og immunoblotting Elektroforese og immunoblotting er en metode for å separere og detektere proteiner i biologiske prøver. Separasjon av proteiner ble utført ved hjelp av natriumdodesylsulfatpolyakrylamidgelektroforese (SDS-PAGE). Proteinene ble så overført fra polyakrylamidgel til en nitrocellulosemembran ved elektrisk overføring (blotting) [58]. Deretter ble membranen inkubert med et spesifikt primært antistoff (Tabell 3.1). Det primære antistoffet bindes til proteinet av interesse. Et sekundært antistoff bindes deretter til det primære antistoffet. I dette arbeidet er det blitt benyttet to metoder for deteksjon av protein på membran; deteksjon av kjemiluminescens på ChemiDoc XRS + og deteksjon av infrarød fluorescens på Odyssey CLx Near-Infrared Imaging System. Etter at prøver ble justert for proteinmengde ved acetonfelling (kapitel 3.5) ble proteinpelleten løst opp i 13 μl vann, 5 μl 4X sample buffer og 2 μl 0,5 M DTT. Prøvene ble så blandet godt og varmet i 10 minutter ved 72 C. Oppvarming av prøver med DTT fører til at proteiner denaturer og disulfidbroer i proteinstrukturen brytes. Natriumdodekylsulfat (SDS) bidrar i tillegg til denatureringen og gir proteinene en uniform negative ladning. Den relative vandringen av proteinene i det 20
34 elektriske feltet vil da kun avhenge av deres størrelse. Tabell 3.1. Oversikt over primærantistoffer som er brukt i denne oppgaven Protein Primærantistoff Fortynning Legumain Geit anti-human polyklonalt 1:200 antistoff mot legumain (AF2199) Cathepsin L Geit anti-human polyklonalt 1:500 antistoff to cathepsin L (MAB952) CD80 Geit anti-human polyklonalt 1:2000 antistoff mot CD80 (AF140) CD163 Geit anti-human polyklonalt 1:2000 antistoff mot CD163 (AF1607) CD206 Geit anti-human polyklonalt 1:200 antistoff mot CD206 (AF2534) Fibronektin Mus anti-human monoklonalt 1:1000 antistoff mot fibronektin (MAB1918) GAPDH Mus anti-human monoklonalt 1: antistoff mot GAPDH (Sc-47724) Etter klargjøring av prøver til elektroforese ble prøver påsatt en ferdigstøpt 12-brønners polyakrylamidgel (NuPAGE 4-12 % Bis-Tris Gel) som på forhånd var plassert i et elektroforesekar fylt med elektroforesebuffer (vedlegg A 1.6). Antioksidant (NuPAGE Antioxidant; 500 μl) ble så tilsatt elektroforesekaret, før prøve (10 eller 15 μl) og standard (Precision Plus Protein ) ble applisert på polyakrylamidgelen i ferdigstøpte brønner. For fremkalling på ChemiDoc XRS + ble det applisert 5 μl standard, mens 1 μl standard ble applisert ved fremkalling på Odyssey CLx Near-infrared Imaging System. Elektroforesen gikk ved 200 volt i 50 minutter. Filterpapir og nitrocellulosemembranen ble fuktet med blottebuffer (vedlegg A 1.7) og proteinene ble overført til nitrocellulosemembranen i en blottemaskin ved 25 volt i 30 minutter. Etter blotting ble membranen Ponceau-farget for visualisering av protein og overført fra gel til membran. Membranen ble så vasket med T-TBS (vedlegg A 1.8) og blokket i 1 time med henholdsvis 5 % tørrmelk i PBS for fremkalling på ChemiDoc XRS + og 5 % tørrmelk i PBS for fremkalling på Odyssey CLx Near-infrared Imaging System. Blokking ble gjort for å forhindre uspesifikk binding av antistoff til membranen ved å mette uspesifikke 21
35 bindingsseter. Etter blokking ble membranen tilsatt primært antistoff (Tabell 3.1) fortynnet i Blotto (ChemiDoc XRS) eller T-TBS (Odyssey CLx Near-Infrared Imaging System) og inkubert over natt. Påfølgende dag ble membranen vasket 4 ganger á 10 min med T-TBS før tilsetting av sekundært antistoff (Tabell 3.2 og Tabell 3.3), fortynnet i henholdsvis Blotto eller T-TBS, og inkubert i 1 time. Tabell 3.2. Oversikt over sekundærantistoffer som er brukt ved fremkalling på ChemiDoc XRS Sekundærantistoff brukt ved fremkalling på ChemiDoc Fortynning XRS Kanin anti-geit polyklonalt antistoff mot immunoglobulin 1:2000 (PO160) Geit anti-mus monoklonalt antistoff ( ) 1:2000 1:1500 Etter tilsetning av sekundærantistoff som ble benyttet for fremkalling Odyssey CLx Nearinfrared Imaging System (Tabell 3.3) ble membranen lysbeskyttet frem til fremkalling og under påfølgende oppbevaring i kjøleskap. Etter inkubering med sekundærantistoff ble membranen igjen vasket 4 ganger á 10 minutter i T-TBS før fremkalling. For fremkalling på ChemiDoc XRS ble membranen etter siste vask lagt i totalt 5 ml fremkallingsvæske (SuperSignal West Dura Extended Duration Substrate) i 5 minutter før fremkalling. Sekundærantistoffene er konjugert med Horseradish Peroxidase (HRP) og etter tilsetning av fremkallingsvæsken vil det dannes kjemiluminiscerende lyskompleks som detekteres i ChemiDoc XRS. For fremkalling på Odyssey CLx Near-infrared Imaging System benyttes det en infrarød laser til deteksjon av proteinbånd. Sekundærantistoffene som anvendes her inneholder infrarøde fluorescerende fargestoffer som eksiteres av laser og deretter emitterer infrarødt lys som detekteres. De sekundære antistoffene som er merket med IRDye detekteres enten ved 700 nm (rødt) eller 800 nm (grønt). Kolokalisering av proteiner som detekteres både ved 700 nm og 800 nm ses i form av et gult signal. 22
36 Tabell 3.3. Oversikt over sekundærantistoffer som er brukt ved fremkalling på Odyssey CLx Near-Infrared Imaging System Sekundærantistoff brukt ved fremkalling på Odyssey CLx Fortynning Near-Infrared Imaging System IRDye 680LT Esel anti-geit immunoglobulin ( ) 1: IRDye 800CW Streptavidin ( ) 1: Reblotting Etter fremkalling ble membran skylt to ganger med T-TBS og så strippet med 10 ml strippebuffer (Restore Western Blot stripping Buffer) i 20 minutter. Membranen ble så skylt to ganger til med T-TBS og deretter vasket i 15 minutter med T-TBS før den ble blokket i 1 time. Etter blokking ble membranen tilsatt nytt primær- og sekundærantistoff som beskrevet ovenfor. 3.9 Enzyme-linked immunosorbent assay (ELISA) ELISA ble brukt for å kvantifisere mengde av legumain i kondisjonert medium. ELISA-kit fra R&D (Human Total Legumain DuoSet ELISA) ble benyttet og produsentens protokoll ble fulgt. En 96-brønners mikrotiterplate blir først behandlet med capture antibody. Dette antistoffet ble fortynnet til 2 μg/ml i PBS og 100 μl ble tilsatt til hver brønn. Platen ble så dekket med plastfilm og inkubert over natt. På dette vis behandles overflaten av brønnene med spesifikt antistoff som vil binde til legumain som eventuelt er til stede i prøvene. Etter inkubering over natt ble brønnene vasket 3 ganger med 400 μl vaskebuffer (0,05 % Tween i PBS), tilsatt 300 μl fortynningsreagens ( Reagent Dilution fortynnet 1:10 i vann) og inkubert i 1 time. Platen ble så vasket igjen som over. Etter siste vask ble 100 μl fortynning av prøver og standarder i fortynningsreagens tilsatt brønnene. Platen ble så inkubert i 2 timer, etterfulgt av ny vask. Deretter ble 100 μl av det sekundære antistoffet tilsatt brønnene. Platen ble igjen inkubert i 2 timer for så å bli vasket. Streptavidinkonjugert-HRP (100 μl) fortynnet i fortynningsreagens (1:200) ble så tilsatt hver brønn og platen ble inkubert i 20 minutter. Det sekundære antistoffet er konjugert med biotin og streptavidinkonjugert-hrp vil da binde seg 23
37 til det sekundære antistoffet. Etter inkubering med Streptavidinkonjugert-HRP ble platen vasket en siste gang før 100 μl substratløsning ble tilsatt hver brønn og inkubert i 20 minutter. Etter denne siste inkuberingen ble 50 μl stoppløsning tilsatt hver brønn og absorbansen ble umiddelbart målt i mikroplateleser (Wallac 1420 VictorX4 Multilabel counter) ved 450 nm. Alle inkuberinger ble gjort ved romtemperatur Statistiske analyser I denne oppgaven er resultatene angitt som gjennomsnitt ± standardfeil (SEM). Gjennomsnittet er beregnet ut i fra antall paralleller av forsøkene (n) og varierer fra 1-5. Signifikansnivået ble satt til p 0,05, og statistisk signifikante forskjeller er angitt med en stjerne (*). Den statistiske metoden som ble brukt i denne oppgaven er en to-utvalgs t-test (Mann-Whitney test). De statiske analysene ble beregnet ved hjelp av GraphPad Prism 6. 24
38 4 Resultater 4.1 Differensiering av THP-1 monocytter til M1- og M2- makrofager Det er tidligere vist økte nivåer av legumain mrna, legumainprotein og legumainaktivitet i makrofagrike områder av ustabile aterosklerotiske plakk [27, 33]. Både M1- og M2- makrofager finnes i aterosklerotiske plakk og tidligere eksperimentelle studier underbygger at M1 hovedsakelig er til stede i ustabile områder, mens M2 hovedsakelig er til stede i stabile områder [59]. Et mål i dette arbeidet var å undersøke forskjell i uttrykk, aktivitet og sekresjon av legumain mellom makrofagsubtypene Legumain uttrykkes forskjellig i M1- og M2-makrofager differensiert med lav konsentrasjon PMA (5 ng/ml) THP-1 celler ble benyttet som cellemodell for å differensiere makrofager til henholdsvis M1 og M2, og for å undersøke effekt av differensiering på uttrykk av legumain. Molekylære former av legumain i cellelysatene ble undersøkt ved immunoblotting. THP-1 celler kun behandlet med lav dose PMA (5 ng/ml; kontrollmakrofager) uttrykte bare prolegumain (56 kda; Figur 4.1.A). I lysat fra M1 ble det detektert proteinbånd ved 56 kda og 36 kda, som korresponderte til prolegumain og moden legumain. I M2 ble det observert et dobbeltbånd 56 kda, som også ble observert i M1. Dessuten viste M1-makrofager høyere uttrykk av prolegumain, sammenlignet med både M2-makrofager og kontrollen. For å verifisere differensieringen av THP-1-celler til M1- og M2-makrofager, ble immunoblottet tilsatt antistoff mot CD80 og CD163 (Figur 4.1.A). CD80 og CD163 er membranreseptorer som i litteraturen er beskrevet som henholdsvis M1-markør (CD80) og M2-markør (CD163) [54]. Etter behandling med LPS og IFN ble det sett høyt uttrykk av CD80 (55 kda), mens i celler behandlet med IL-4 og i PMA-kontrollen ble det ikke observert 25
 L P S (1 g /m l) IF N (2 0 n g /m l) IL -4 (2 0 n g /m l) + + + - - + - - + - + - P M A (5 n g /m l) L P S (1 g /m l) IF N (2 0 n g")
 L P S (1 g /m l) IF N (2 0 n g /m l) IL -4 (2 0 n g /m l) + + + - + - - - + - - + Figur 4.1.: Ulikt uttrykk av legumain i M1- og M2 makrofager behandlet med lav konsentrasjon av PMA.")
39 T o ta lp ro te in (µ g /m l) A E P -a k tiv ite t (d F /s e c ) /to ta lp r o te in (µ g /m l) A E P -a k tiv ite t (d F /s e k ) A k D a L e g u m a in C D B C D G A P D H P M A (5 n g /m l) L P S (1 g /m l) IF N (2 0 n g /m l) IL -4 (2 0 n g /m l) P M A (5 n g /m l) L P S (1 g /m l) IF N (2 0 n g /m l) IL -4 (2 0 n g /m l) C D * 0 0 P M A (5 n g /m l) L P S (1 g /m l) IF N (2 0 n g /m l) IL -4 (2 0 n g /m l) P M A (5 n g /m l) L P S (1 g /m l) IF N (2 0 n g /m l) IL -4 (2 0 n g /m l) Figur 4.1.: Ulikt uttrykk av legumain i M1- og M2 makrofager behandlet med lav konsentrasjon av PMA. PMA-stimulerte (5 ng/ml, 48 timer) THP-1 celler (1 mill./2 ml; 6- brønners brett) ble vasket og inkubert videre i 72 timer ved 37 C med enten LPS (1 μg/ml) og INF (20 ng/ml; M1), IL-4 (20 ng/ml; M2) eller dyrkningsmedium (kontrollmakrofager). A, Cellelysat ble oppkonsentrert til gelelektroforese ved acetonfelling og lik mengde totalprotein (10 μg/brønn) ble applisert. Uttrykk av legumain, CD80 og CD163 ble analysert 26
40 etter immunoblotting og spesifikke antistoffer. GAPDH ble brukt som housekeeping kontroll. B, AEP-aktiviteten ble målt i cellelysat ved bruk av et spesifikt fluorescerende peptidsubstrat (Z-Ala-Ala-Asn-AMC) (n=1). C, Totalproteinkonsentrasjonen (µg/ml) i cellelysat (n=1). D, AEP-aktiviteten (df/sek) i cellelysater korrigert for totalprotein (df/sek/totalprotein) (n=4, *p<0,05 for M1 vs. M2 eller kontroll, uparet test). CD80. Dette bekreftet at differensiering av THP-1 celler til M1-makrofager ble oppnådd etter behandling med LPS og IFN. Immunoblotting med antistoff mot CD163 (130 kda) viste tilstedeværelse av CD163 i celler behandlet med IL-4 (M2-makrofager) i form av et svakt proteinbånd som også ble sett i kontrollen, mens denne markøren var fraværende i M1- makrofager. GAPDH ble brukt som housekeeping kontroll på blottet (Figur 4.1.A). Proteolytisk aktivitet overfor peptidsubstratet Z-Ala-Ala-Asn-AMC (AEP-aktivitet), ble målt i cellelysatene. AEP-aktiviteten (df/sek) i cellelysatene uten korrigering for totalproteinkonsentrasjon viste at aktiviteten var høyere i M2-makrofager og lavere i M1- makrofager i forhold til kontroll (Figur 4.1.B). Totalproteinkonsentrasjon ble målt i cellelysatene og viste at innholdet generelt var lavt og at M2-lysatene (Figur 4.1.C) inneholdt mer totalprotein enn M1-lysatene og kontroll. Når AEP-aktiviteten ble korrigert for totalprotein (df/sek/totalprotein) var aktiviteten signifikant høyere i både M2-makrofager og kontroll sammenlignet med M1-makrofager (Figur 4.1.D). Under cellearbeidet ble det observert mange løse celler etter PMA-behandling (5 ng/ml) av THP-1-monocytter i 72 timer, samt at vasking av celler før ytterligere stimulering eller før høsting også førte til at celler løsnet og ga lavt utbytte. For videre undersøkelser av M1- og M2-makrofager var det ønskelig å bytte til en differensieringsmetode som ga større celleadherens. En alternativ metode er beskrevet av Halvorsen et al. [60] som anvender en betydelig høyere PMA-konsentrasjon (62 ng/ml), og hvor de native THP-1 cellene eksponeres for PMA i kun 6 timer. Konsentrasjonene av LPS, IFN og IL-4 avviker også noe fra betingelsene brukt ved den første differensieringsmetoden. Se tabell 4.1 for oversikt av metode 1 (lav PMA) versus metode 2 (høy PMA). 27
 (kons/tid) PMA 5 ng/ml 62 ng/ml Kontrollmakrofag 48 timer 6 timer LPS + IFN 1 μg/ml + 10 ng/ml + M1 20 ng/ml 72 timer 5 ng/ml 72 timer IL-4 20")
 og M2- makrofager (Figur 4.1.C) hadde ulik morfologi: M1-makrofager hadde en mer forgrenet form, mens M2-makrofager og kontrollmakrofager var mer sirkulære i form.")
 THP-celler (1 mill.")
41 Tabell 4.1: Oversikt over metodene som ble brukt for å differensiere THP-1 monocytter til M1- og M2-makrofager. Behandling Metode 1 Metode 2 Makrofagtype (kons/tid) (kons/tid) PMA 5 ng/ml 62 ng/ml Kontrollmakrofag 48 timer 6 timer LPS + IFN 1 μg/ml + 10 ng/ml + M1 20 ng/ml 72 timer 5 ng/ml 72 timer IL-4 20 ng/ml 72 timer 25 ng/ml 72 timer M Ulik morfologi av M1- og M2-makrofager Figur 4.2. viser kontrollmakrofager (A), samt M1-makrofager (B) og M2-makrofager (C) differensiert etter metode 2. Det ble observert at M1-makrofager (Figur 4.2.B) og M2- makrofager (Figur 4.1.C) hadde ulik morfologi: M1-makrofager hadde en mer forgrenet form, mens M2-makrofager og kontrollmakrofager var mer sirkulære i form. Celler stimulert med høy PMA (metode 2), sammenlignet med lav PMA (metode 1), var også mer adherente. Figur 4.2.: M1- og M2-makrofager utviste ulik karakteristikk under mikroskop. M1- og M2-makrofager differensiert fra THP-1 monocytter etter metode 2. PMA-stimulerte (62 ng/ml, 6 timer) THP-celler (1 mill./2 ml; 6-brønners brett) ble vasket og inkubert videre i 72 timer ved 37 C med enten LPS (10 ng/ml) og INF (5 ng/ml; M1), IL-4 (25 ng/ml; M2) eller dyrkningsmedium (kontrollmakrofager) før høsting. A, Kontrollmakrofager. B, M1- makrofager. C, M2-makrofager. Bilder av celler er tatt rett før høsting med 10 X forstørrelse i lysmikroskop. 28
42 4.1.3 Legumain uttrykkes forskjellig i M1- og M2-makrofager behandlet med høy konsentrasjon av PMA (62 ng/ml) THP-1 celler ble sådd ut og stimulert med høy konsentrasjon PMA (62 ng/ml) i 6 timer. Etter PMA-stimuleringen ble celler vasket og inkubert i enten 18 timer eller 72 timer med dyrkningsmedium tilsatt enten LPS (10 ng/ml) og IFN (5 ng/ml) for å differensiere til M1- makrofager eller IL-4 (25 ng/ml) for å differensiere til M2-makrofager. I M1-makrofager ble det ved immunoblotting observert kraftig proteinbånd for prolegumain (56 kda) etter differensiering i både 18 og 72 timer (Figur 4.3.A), mens M1-makrofager differensiert i 72 timer uttrykte mer moden legumain (36 kda) enn etter 18 timer. Uttrykk av prolegumain i M2-makrofager (18 og 72 timer) ble sett i form av svakt dobbeltbånd ved 56 kda, som også ble observert i kontrollen. I M2-makrofager og kontroll ble det kun observert moden legumain etter differensiering i 72 timer. Immunoblotting viste i tillegg et overraskende kraftig proteinbånd ved omtrent 75 kda i M2-makrofager som ble differensiert i 72 timer. Uttrykk av CD80 ble observert i THP-1 celler behandlet med LPS (10 ng/ml) og IFN (5 n g/ml)(m1) i både 18 timer og 72 timer (Figur 4.3.A). Et tilsvarende uttrykk ble ikke observert i makrofager stimulert med IL-4 (25 ng/ml) eller i kontroll. Svakt proteinbånd for CD163 ble observert i alle cellelysatene, med unntak av celler behandlet med IL-4 (25 ng/ml) i 72 timer. Overraskende nok ble det kraftigste proteinbåndet for CD163 sett i 72 timers kontrollen. Figur 4.3. B og C viser henholdsvis totalproteinkonsentrasjonen og AEP-aktiviteten i cellelysater etter 72 timers differensiering. Totalproteinkonsentrasjonen i cellelysater etter metode 2 (Tabell 4.1) var høyere i M2-makrofager enn i M1-makrofager og kontroll, tilsvarende det som ble observert ved den første differensieringsmetoden (Figur 4.1.B; Metode 1, Tabell 4.1). I samsvar med den første differensieringsmetoden (metode 1) ga også protokollen med høy PMA-konsentrasjon i 6 timer (metode 2) høyere AEP-aktivitet i lysater fra M2-makrofager enn M1-makrofager (Figur 4.3.C). Det ble derfor besluttet å bruke metode 2 i videre studier av makrofager i kultur. 29
 + + + + + + L P S (1 0 n g /m l) IF N (5 n g /m l) - - - - + + + + - - - - IL -4 (2")
43 T o ta lp ro te in (µ g /m l) A E P -a k tiv ite t (d F /s e k )/ to ta ltp r o te in (µ g /m l) A K M 1 M 2 T im e r; d iff k D a 75 L e g u m a in C D 8 0 C D G A P D H 37 P M A (6 2 n g /m l) L P S (1 0 n g /m l) IF N (5 n g /m l) IL -4 (2 5 n g /m l) B C P M A (6 2 n g /m l) L P S (1 0 n g /m l) IF N (5 n g /m l) IL -4 (2 5 n g /m l) P M A (6 2 n g /m l) L P S (1 0 n g /m l) IF N (5 n g /m l) IL -4 (2 5 n g /m l) Figur 4.3.: Ulikt uttrykk av legumain i M1- og M2-makrofager behandlet med høy konsentrasjon av PMA. PMA-stimulerte (62 ng/ml, 6 timer) THP-celler (1 mill./2 ml; 6- brønners brett) ble vasket og inkubert videre i 72 timer ved 37 C med enten LPS (10 ng/ml) og INF (5 ng/ml; M1), IL-4 (25 ng/ml; M2) eller dyrkningsmedium (kontrollmakrofager) før høsting. A, Cellelysat ble oppkonsentrert til gelelektroforese ved acetonfelling og lik mengde 30
44 L e g u m a in s e k re s jo n (p g /m l) totalprotein (15 μg/brønn) ble applisert. Uttrykk av legumain, CD80 og CD163 ble analysert ved immunoblotting og fremkalling med spesifikke antistoffer. GAPDH ble brukt som housekeeping kontroll. B, Totalproteinkonsentrasjonen (µg/ml) i cellelysat (n=2-5). C, AEP-aktiviteten (df/sek) i cellelysat ved bruk av et spesifikt fluorescerende peptidsubstrat (Z- Ala-Ala-Asn-AMC) og korrigert for totalprotein (df/sek/totalprotein) (n=2-5) Høyere sekresjon av legumain fra M1- enn M2-makrofager I tillegg til å undersøke det intracellulære uttrykket av legumain og AEP-aktivitet i M1- og M2-makrofager var det ønskelig å studere om M1- og M2-makrofager kunne skille ut legumain og om det var forskjell mellom makrofagtypene med hensyn på mengde legumain som ble utskilt. Figur 4.1. A og 4.3.A viste uttrykk av molekylære former av legumain i M1- og M2-makrofager som ble differensiert med henholdsvis metode 1 og 2 (Tabell 4.1) og det ble sett at uttrykk av prolegumain var høyere i M1-makrofager enn M2-makrofager etter differensiering ved begge metodene. For å kvantifisere sekresjon av legumain fra makrofager differensiert med metode 2 ble det utført ELISA-målinger av totallegumain i kondisjonerte medier fra høsting av cellene. Figur 4.4. viser at konsentrasjonen av legumain i de kondisjonerte mediene fra M-1 makrofager er betraktelig høyere enn M2-makrofager, og inneholdt mer enn 10 ganger mer legumain enn M2-makrofager * M 1 M 2 Figur 4.4.: Konsentrasjon av legumain i kondisjonerte medier fra M1- og M2-makrofager. THP-1 celler ble behandlet med PMA (62 ng/ml) i 6 timer, vasket og inkubert i 72 timer ved 37 C med enten LPS (10 ng/ml) og INF (5 ng/ml; M1) eller IL-4 (25 ng/ml; M2). Legumain ble målt ved ELISA i kondisjonerte medier fra henholdsvis M1- og M2-makrofager. Legumainkonsentrasjon (pg/ml) er angitt som gjennomsnittlige verdier ± SEM (n=3). 31
45 *p<0,05 vs. M2, uparet t-test Degradering av fibronektin Lysater av M1- og M2- makrofager viste ulik aktivitet overfor et lavmolekylært peptidsubstrat for legumain (Fig. 4.1.B, D og 4.2.C). I tillegg var det interessant å undersøke om lysater fra M1 og M2 også hadde ulik proteolytiske aktivitet overfor et høymolekylært substrat. Fibronektin er et slikt substrat som i litteraturen er vist å kunne bli spaltet av legumain [26]. Derfor ble rekombinant human fibronektin inkubert med M1- og M2-lysater, renset bovint legumain fra storfenyrer eller cellelysat fra en monoklonal cellelinje som overuttrykker legumain (M38L). Figur 4.5. viser at fibronektin gjennomgår omfattende spaltning og degradering etter inkubering med renset bovint legumain (bleg), som ble brukt som positiv kontroll. Dette eksperimentet ble utført med en relativt høy konsentrasjon av legumain, vekt-ratio ble estimert til 1:10 for enzym:substrat. Inkubatet av fibronektin med lysat fra en cellelinje som overuttrykker legumain (M38L) viste også omfattende degradering. Når inkubat med M38L lysat ble tilsatt inhibitor av legumain (MP-L01*) eller cathepsiner (E-64), reduserte dette degraderingen av fibronektin. Lysater fra M1- og M2-makrofager viste derimot begrenset kapasitet til å degradere fibronektin, men M2 førte til noe mer spaltning av fibronektin enn M1 (Fig. 4.5.; brønn 1 versus 5). M2-spaltningen av fibronektin ble delvis motvirket ved tilstedeværelse av proteaseinhibitorer som MP-L01* (brønn 6), cystatin E/M (brønn 7) eller E-64 (brønn 8). Størst hemmende effekt på spaltningen av fibronektin ble observert av den irreversible og spesifikke legumainhemmeren MP-L01* [41]. 32


, M1 cellelysat (brønn 1), M1 cellelysat med MP-L01* (10 µm, brønn 2), M1 cellelysat med cystatin E/M (5 µl")
, M2 cellelysat med E- 64 (10 µm, brønn 8), bovint legumain (10 µg/ml, bleg,),")
46 A FN M b L E G M 3 8 L M 3 8 L + M P -L 0 1 * M 3 8 L + E 6 4 FN M 2 b L E G M 3 8 L M 3 8 L + M P -L 0 1 * M 3 8 L + E 6 4 B FN FN Figur 4.5.: Immunoblotting av fibronektin inkubert med lysater fra M1-eller M2- makrofager, samt M38L lysat eller bovint legumain. Fibronektin (FN, 5 µl; 100 µg/ml) ble inkubert med cellelysater (10 µl) eller renset bovint legumain (bleg, 10 µl, 10 µg/ml) med eller uten proteaseinhibitor. Etter 24 timer ved 37 C ble elektroforeseprøver av inkubatene applisert på gel, blottet og inkubert med antistoff mot fibronektin. FN (fibronektin kontroll), M1 cellelysat (brønn 1), M1 cellelysat med MP-L01* (10 µm, brønn 2), M1 cellelysat med cystatin E/M (5 µl M4C-kondisjonert medium, brønn 3), M1 cellelysat med E-64 (10 µm, brønn 4), M2 cellelysat (brønn 5), M2 cellelysat med MP-L01* (10 µm, brønn 6), M2 cellelysat med cystatin E/M (5 µl M4C-kondisjonert medium, brønn 7), M2 cellelysat med E- 64 (10 µm, brønn 8), bovint legumain (10 µg/ml, bleg,), M38L cellelysat (M38L), M38L cellelysat med MP-L01* (10 µm, M38L+MP-L01*), M38L cellelysat med E-64 (10 µm, M38L+E-64). A, immunoblot; B, densitometrisk scanning av A. 33
47 4.1.6 Inkubering av M1- og M2-lysater med aktivitetsbasert legumainprobe THP-celler ble differensiert til M1- og M2-makrofager (metode 2) og cellelysatene ble inkubert med en biotinylert irreversibel spesifikk aktivitetsbasert probe for legumain, MP- L01 (1 µm) [41]. Etter gelelektroforese og immunoblotting av inkubatene, ble legumain analysert ved hjelp av spesifikt antistoff og proben analysert ved hjelp av streptavidin ved laserscanning på Odyssey CLx Near-Infrared Imaging System. Også her ble M38L-lysat og renset bovint legumain brukt som positive kontroller. M38L-lysat og renset bovint legumain ble brukt som kontroller på binding av probe til legumain. I M38L-lysat ble det sett binding av legumainantistoff i form av bånd på 56 kda, 46 kda og 36 kda som tilsvarte proform, mellomform og moden form av legumain (Figur 4.6.). Proben ga to fluorescerende bånd som ved samtidig deteksjon med legumainantistoff ga kolokalisering til både mellomform og moden form av legumain i dette lysatet. Renset bovint legumain inneholdt mye moden legumain, men også en 25 kda-form. Det ble sett binding av probe til begge disse formene av legumain. Blottet ble overeksponert på lav fortynningsgrad av legumainantistoff (1:200) da kontrollene var på samme blott som makrofaglysatene, og det var denne antistoffkonsentrasjonen som ble brukt for disse. Etter inkubering av probe med makrofaglysater, ble det kun sett binding av antistoff mot legumain i M2-makrofager i form av et proteinbånd ved ca. 75 kda (Figur 4.6.). For proben ble det sett bånd i M2- og kontrollmakrofager ved 25 kda, som ikke ble detektert i M1- makrofager, men dette båndet ble ikke detektert med legumainantistoffet. I alle makrofaglysatene ble det sett et sterkt bånd ved ca. 75 kda og et svakere bånd rett under 150 kda av endogene biotinylerte proteiner etter blotting med antistoff mot streptavidin. 34
 fra kontrollmakrofager, M1-makrofager, M2-makrofager og M38L celler, samt renset bovint legumain (36 µm) ble inkubert i paralleller uten eller med biotinylert aktivitetsbasert")
 eller 10 µl (M38L og renset bovint legumain) ble applisert i på gel.")
48 M P -L 0 1 (1 µ M ) Figur 4.6.: Inkubering av M1- og M2-lysater med aktivitetsbasert probe. Cellelysat (19 µl) fra kontrollmakrofager, M1-makrofager, M2-makrofager og M38L celler, samt renset bovint legumain (36 µm) ble inkubert i paralleller uten eller med biotinylert aktivitetsbasert legumainprobe MP-L01 (1 µm) i 30 minutter ved 30 ºC før tilsetning av sample buffer (7,7 µl) og 0,5 M DTT (3,1 µl). Prøvene ble blandet og varmet i 10 minutter ved 70 ºC før 20 µl (kontrollmakrofager, M1 og M2) eller 10 µl (M38L og renset bovint legumain) ble applisert i på gel. Immunoblotting ble detektert på Odyssey CLx Near-Infrared Imaging System og viser legumain (rødt, øverste panel) og probe (grønt, øverste panel) og samlokalisering ( interplay ; gult, øverste panel). 35
49 4.2 Effekt av atorvastatin på M1- og M2-makrofager I en studie gjort av Wang et al. ble det vist at pasienter behandlet med atorvastatin hadde redusert legumain mrna i sirkulerende monocytter [38]. I en annen studie har det også blitt observert signifikant økning av mrna for M2-markører (CD206, IL-10 og CCL18) i sirkulerende monocytter etter behandling med atorvastatin [61]. I denne samme studien ble også sett en doseavhengig økning av CD206 i primære monocytter som ble differensiert til M2-makrofager in vitro og behandlet med atorvastatin. Det var derfor ønskelig å behandle THP-1 celler med atorvastatin under differensieringen til M1- og M2-makrofager for å undersøke om uttrykk, aktivitet og sekresjon av legumain endret seg Økende konsentrasjoner atorvastatin hemmer adherens av M1- og M2-makrofager THP-celler ble behandlet med atorvastatin (0, 1, 5, 10, 25 og 50 μm) under differensieringen. Figur 4.7. viser M1- og M2-makrofager behandlet med av atorvastatin (0, 5 og 25 μm) under differensiering med metode 2. Både M1-makrofager (Figur 4.7.A, C og D) og M2- makrofager (Figur 4.7.B, D og E) ble mindre adherente ved økende konsentrasjoner av atorvastatin, og dette var mest uttalt for M1-makrofager. Etter behandling med 50 μm (ikke vist) ble det observert stort antall av løse celler av både M1- og M2-typen. Ved differensiering uten tilstedeværelse av atorvastatin (Figur 4.2., 4.7. A og B) utviste disse makrofagsubtypene ulik morfologi, som beskrevet tidligere (delkapittel 4.1.2). Denne forskjellen i morfologi var også synlig mellom M1- og M2-makrofager behandlet med 0-25 μm atorvastatin. Imidlertid var det markant færre M1-makrofager som utviste en forgrenet form etter behandling med 25 μm atorvastatin. M1-makrofager behandlet med 50 μm atorvastatin (ikke vist) var nesten utelukkende sirkulære. 36

. E, M1-makrofager (25 μm atorvastatin). F, M2-makrofager (25 μm atorvastatin).")

.")
 så ut til være likt opp til behandling med 25 μm atorvastatin (Figur 4.7.A).")

50 Figur 4.7.: M1- og M2-makrofager behandlet med og uten atorvastatin under differensiering har ulik karakteristikk. M1- og M2-makrofager ble differensiert etter metode 2 og behandlet med ulike konsentrasjoner av atorvastatin under differensieringen. A, M1- makrofager (0 μm atorvastatin). B, M2-makrofager (0 μm atorvastatin). C,M1-makrofager (5 μm atorvastatin). D, M2-makrofager (5 μm atorvastatin). E, M1-makrofager (25 μm atorvastatin). F, M2-makrofager (25 μm atorvastatin). Bilder av celler er tatt rett før høsting med 10 X forstørrelse i lysmikroskop Atorvastatin førte til endret aktivitet, prosessering og sekresjon av legumain i M1- og M2-makrofager Figur 4.8A og B viser uttrykk av legumain, CD80 og CD163 etter behandling med ulike konsentrasjoner av atorvastatin (0-50 μm) under differensiering til henholdsvis M1- og M2- makrofager (metode 2). I M1-differensierte makrofager ble ikke uttrykket av moden legumain (36 kda) endret etter behandling med 0-5 μm atorvastatin, mens uttrykket av prolegumain (56 kda) så ut til være likt opp til behandling med 25 μm atorvastatin (Figur 4.7.A). Housekeeping kontrollen (GAPDH) viste jevn proteinmengde i M1- og M2-makrofager behandlet med 0-25 μm atorvastatin, mens det var markant lavere protein i M1-makrofager og litt lavere i M2- makrofager behandlet med 50 μm atorvastatin. I M2-differensierte makrofager ble det også sett at mengde moden legumain og prolegumain var uendret etter behandling med lave konsentrasjoner atorvastatin (0-5 μm) (Figur 4.8.B). I motsetning til M1, så ble det derimot sett samtidig økning i mengde prolegumain og nedgang 37
51 i mengde moden legumain ved økende konsentrasjoner av atorvastatin (10-50 μm) og etter behandling med høyeste konsentrasjon atorvastatin ble det ikke sett moden legumain. Det ble observert et proteinbånd detektert med legumainantistoff ved omtrent 75 kda i både ubehandlede M2-differensiert makrofager og etter behandling med 1-10 μm atorvastatin. Dette båndet var svakere etter behandling med 10 μm atorvastatin enn 1-5 µm og forsvant etter behandling med 25 og 50 μm atorvastatin. I M1 makrofager (Figur 4.8.A) ble det ikke sett et slikt 75 kda-bånd. Tilstedeværelse av dette 75 kda proteinet i M2-makrofager ble også sett tidligere (Figur 4.2.A). I både M1- og M2-makrofager ble det også sett proteinbånd ved 25 kda etter blotting med antistoff mot legumain. I M1-makrofager var proteinbåndet ved 25 kda relativt like etter behandling med alle konsentrasjoner av atorvastatin, inkludert høyeste konsentrasjon (50 μm). I M2-makrofager, derimot, så ble det sett høyere intensitet av dette båndet etter behandling med 25 og 50 μm atorvastatin. I M1-makrofager var det høyt uttrykk av M1-markøren CD80 (55 kda), som ikke ble observert i M2-makrofager (Figur 4.8.A og B). Etter blotting med antistoff mot M2-markøren CD163 ble det observert et svakt proteinbånd med størrelse < 150 kda, som var den forventede molekylvekten til CD163. Ved måling av enzymaktivitet ble det observert en nedgang i AEP-aktiviteten (df/sek/totalprotein) i både M1- og M2-makrofager etter behandling med økende konsentrasjon av atorvastatin (Figur 4.5.C). I M1-makrofager var det ingen målbar AEPaktivitet etter behandling med konsentrasjoner over 10 μm, mens i M2-makrofager ble aktiviteten kraftig redusert etter μm atorvastatin. Samme undersøkelser ble også gjort med makrofager differensiert etter metode 1. Observerte effekter var tilsvarende som for metode 2, og i tillegg viste behandling av THP-1 celler med 100 og 200 μm atorvastatin at det var enda færre adherente celler og lavere totalproteinkonsentrasjon (μg/ml) i cellelysater (data ikke vist). 38

 0 1 5 1 0 2 5 5 0 37 2 E M 2 k D a 37 0 0")

 THP-1 celler (1 mill.")
, IL-4 (25 ng/ml; M2).")
 Cellelysat ble oppkonsentrert til gelelektroforese ved acetonfelling og lik mengde")
52 A E P -a k tiv ite t (d F /s e k )/ to ta ltp r o te in (µ g /m l) A M 1 k D a 75 B M 2 k D a 75 L e g u m a in L e g u m a in C D C D C D C D G A P D H 37 G A P D H 37 A (µ M ) A (µ M ) C 6 M 1 M 2 D M 1 C a th e p s in -L k D a G A P D H A ( µ M ) E M 2 k D a A to rv a s ta tin, (µ M ) C a th e p s in -L G A P D H A ( µ M ) Figur 4.8.: Endret uttrykk av legumain i THP-1 celler behandlet med økende konsentrasjoner av atorvastatin under M1- og M2-differensiering. PMA-stimulerte (62 ng/ml, 6 timer) THP-1 celler (1 mill./2 ml; 6-brønners brett) ble vasket og inkubert videre i 72 timer ved 37 C med enten LPS (10 ng/ml) og INF (5 ng/ml; M1), IL-4 (25 ng/ml; M2). Under differensieringen ble THP-1 cellene behandlet med atorvastatin (A;0-50 μm). (A, B) Cellelysat ble oppkonsentrert til gelelektroforese ved acetonfelling og lik mengde totalprotein (15 μg/brønn) ble applisert. Uttrykk av legumain, CD8, CD163 og cathepsin L ble analysert etter immunoblotting og fremkalling med spesifikke antistoffer. GAPDH ble brukt som 39
53 housekeeping kontroll. C, AEP-aktiviteten (df/sek) i cellelysat ved bruk av et spesifikt fluorescerende peptidsubstrat (Z-Ala-Ala-Asn-AMC) korrigert for totalprotein f(df/sek/totalprotein) (n=3) Atorvastatin hemmer prosessering av cathepsin L i makrofager Cathepsin L er assosiert med plakkdestabilisering og det er vist at legumain spiller en rolle i prosessering og aktivering av denne cysteinproteasen [18, 62]. Derfor ble uttrykk av cathepsin L undersøkt i atorvastatinbehandlede M1- og M2-makrofager ved immunoblotting. Figur 4.8.D og E viser uttrykk av cathepsin L i atorvastatinbehandlede M1- og M2- makrofager. Det ble sett et relativt stabilt uttrykk av procathepsin L (43 kda) i M1- makrofager behandlet med økende konsentrasjoner atorvastatin, mens behandling 10 μm atorvastatin førte til mer procathepsin L i M2-makrofager. Uttrykket av procathepsin L (43 kda) og two-chain cathepsin L (25 kda) så ut til å være høyere i M1-makrofager enn i M2- makrofager, og i begge celletyper ble det sett nedgang i både single-chain og two-chain, etter behandling med 25 μm atorvastatin. Denne nedgangen i single-chain - og twochain -cathepsin L sammenfalt med nedgang i uttrykk av moden legumain i M1- og M2- makrofager (Figur 4.8. A og B) Atorvastatin hemmer sekresjon av legumain fra M1-makrofager Det ble gjort ELISA-målinger av totalmengden legumain i kondisjonerte medier fra M1- og M2-makrofager som ble behandlet med atorvastatin under differensieringen (0-50 μm). Etter behandling med 0-25 μm atorvastatin var legumainkonsentrasjon (pg/ml) i kondisjonerte medier fra M1-makrofager signifikant høyere enn legumainkonsentrasjonen (pg/ml) i kondisjonerte medier fra M2-makrofager (Figur 4.9.). I medier fra M2-makrofager var legumainkonsentrasjonen lav, og relativt uendret både med og uten behandling med atorvastatin (0-50 μm). Derimot så minket mengden av legumain skilt ut fra M1-makrofager kraftig etter behandling med 1-10 μm atorvastatin. Etter behandling med 50 μm atorvastatin var det skilt ut like lite legumain fra M1-makrofager som M2-makrofager. 40
54 L e g u m a in s e k re s jo n (p g /m l) A M 1 M * A to rv a s ta tin, (µ M ) Figur 4.9.: Legumainkonsentrasjon i kondisjonerte medier fra makrofager behandlet med atorvastatin under differensieringen. Legumain ble målt ved ELISA i kondisjonerte medier fra M1- og M2-makrofager som ble behandlet med atorvastatin (0-50 μm) under differensiering (metode 2). Legumainkonsentrasjonen (pg/ml) er angitt som gjennomsnittlige verdier ± SEM (n=3). *p<0,05 for 10 µm atorvastatin M1 vs. 0 µm atorvastatin M1, uparet t-test) AEP-aktivitet versus legumainaktivitet i M1- og M2-makrofager behandlet med atorvastatin Ved immunoblotting ble det observert at M1-makrofager uttrykte mer moden legumain (36 kda) enn M2-makrofager (Figur 4.3.A), mens det ble sett høyere AEP-aktivitet i M2- makrofager enn i M1-makrofager (Figur 4.3.C). Av denne grunn var det interessant å undersøke hvordan AEP-aktiviteten i atorvastatinbehandlede M1- og M2-makrofager ble påvirket etter tilsetning av E-64, som er et epoksid som hemmer generelt cysteincathepsiner [15, 40], men ikke legumain [5, 14]. Etter tilsetning av E-64 (sluttkonsentrasjon 1 μm) til cellelysater fra M1- og M2-makrofager som var behandlet med 0-50 μm atorvastatin ble lysatene inkubert ved 37 C og to enkle målinger av fluorescens ble gjort; etter 90 minutter og 6 timer. Ut ifra disse målingene ble det AEP-aktivitet (df/sek/totalprotein) beregnet. AEP-aktivitet uten tilsetning av E-64 ble antatt å være den totale aktiviteten, mens aktivitet etter tilsetning av E-64 (restaktivitet) ble antatt å 41
55 være faktisk legumainaktivitet. Differansen mellom total AEP- og restaktivitet (legumain) ved tilstedeværelse av E-64 var AEP-aktivitet som skyldtes et eller flere cysteincathepsiner. Figur 4.10.A (M1) og B (M2) viser AEP-aktiviteten (df/sek/totalprotein) i cellelysater. Det totale arealet av hver søyle representerer den totale AEP-aktiviteten i cellelysatet, mens de røde områdene representerer legumainaktiviteten. Svart eller grå del av søylene representerer AEP-aktivitet av cysteincathepsin som ble hemmet av E-64. Den totale AEP-aktiviteten var lavere i M1-makrofager (Figur 4.10.A) enn i M2-makrofager (Figur 4.10.B) og ble kraftig redusert etter behandling med 25 og 50 μm atorvastatin i begge makrofagtypene. Figur 4.10.C (M1) og D (M2) viser legumainaktivitet (rødt) som prosentandel av den totale AEPaktiviteten. Andelen legumainaktivitet i ubehandlede makrofager (0 μm atorvastatin) var høyere i M1-makrofager (37 %) enn M2-makrofager (17 %). Denne relative forskjellen i legumainaktivitet mellom M1- og M2-makrofager forble uendret med økende doser atorvastatin. 42
56 % A E P -a k tiv ite t (d F /s e k )/ to ta ltp r o te in (µ g /m l) % A E P -a k tiv ite t (d F /s e k )/ to ta ltp r o te in (µ g /m l) A E P -a k tiv ite t (d F /s e k )/ to ta ltp r o te in (µ g /m l) A E P -a k tiv ite t (d F /s e k )/ to ta ltp r o te in (µ g /m l) A B 5 M 1 5 M A to rv a s ta tin (µ M ) A to rv a s ta tin (µ M ) C M 1 D M A to rv a s ta tin (µ M ) A to rv a s ta tin (µ M ) Figur 4.10.: AEP-aktivitet i M1- og M2-makrofager etter tilsetning av cathepsinhemmeren E-64. THP-1 celler ble under differensiering til M1- og M2-makrofager (metode 2) behandlet med atorvastatin (0-50 μm) i 72 timer. For måling av AEP-aktivitet (df/sek) med og uten E- 64 ble 20 μl cellelysat tilsatt en 96-brønners mikrotiterplate i duplikater. To og to brønner ble så tilsatt 17 μl vann eller 17 μl E4 (10 μm) til en sluttkonsentrasjon på 1 μm i hver brønn og inkubert ved 37 C. Fluorescensmålinger ble gjort etter 90 minutter og 6 timer, og AEPaktivitet ble beregnet basert på målt fluorescens ved disse tidspunktene. A, B, AEP-aktivitet (df/sek) korrigert for totalprotein (df/sek/totalprotein) i henholdsvis M1- og M2- cellelysater. Summen av de sorte og røde segmentene i hver søyle viser AEP-aktivitet uten E- 64 til stede. De røde segmentene viser AEP-aktiviteten etter tilsetning av E-64 (restaktivitet). C, D, Restaktivitet i henholdsvis M1- og M2-makrofager (rødt) som prosentandel av AEPaktivitet uten E-64 til stede. 43
57 5 Diskusjon I denne oppgaven ble den humane monocyttiske cellelinjen THP-1 brukt som cellemodell og differensiert til M1- og M2-makrofager ved først å behandle cellene med forbolesteren PMA, og deretter stimulere cellene med henholdsvis LPS/IFNγ (M1) eller IL-4 (M2). For å kontrollere differensieringen til M1- og M2-makrofager ble det benyttet markører som i litteraturen er angitt å være M1- og M2-markører; CD80 som M1-markør og CD163 som M2-markør. I dette arbeidet ble det benyttet to forskjellige metoder for differensiering til M1- og M2-makrofager. Det ble observert at behandling med høy (62 ng/ml) PMA-konsentrasjon (metode 2) ga mer adherente celler enn behandling med lav (5 ng/ml) PMA-konsentrasjon (metode 1), men at lav konsentrasjon av PMA over lengre tid økte differensieringen til makrofager vurdert ut ifra sterkere proteinuttrykk av CD80. Både uttrykk, aktivitet og sekresjon av legumain i M1- og M2-makrofager ble undersøkt. Uttrykket av legumain i M1- og M2-makrofagene var ulikt. Immunoblotting viste at M1- makrofager har et høyt uttrykk av prolegumain, sammenlignet med M2-makrofager. I M2- makrofager differensiert etter metode 2 ble det observert en legumainform på omtrent 75 kda, som ikke ble observert i M1-makrofager. I denne oppgaven ble det observert at både M1- og M2--makrofager sekrerer legumain, og at sekresjonen var signifikant høyere fra M1- makrofager (omtrent 10 ganger høyere) enn fra M2-makrofager. Det ble også observert uttrykk av en 25 kda-form av legumain i både M1- og M2-makrofager. Ved inkubering av M1- og M2-lysater med den legumainaktivitetsbaserte proben MP-L01 ble det sett binding av probe til denne 25 kda-formen. Studier av makrofager har vist at det et stort mangfold i deres form og funksjon, som sannsynligvis reflekterer ulik differensiering i deres mikromiljø [42, 63]. Makrofager kan igangsettes til å produsere M1-fenotype etter stimulering med bakterier, bakterielle celleveggskomponenter og cytokiner som IFNγ, GM-CSF og TNFα. Makrofager av M1- typen kjennetegnes blant annet ved at de sekrerer inflammatoriske cytokiner og produserer nitrogenmono-oksid (NO) [46]. Makrofager av M2-typen igangsettes av parasitter, apoptopiske celler og cytokiner som IL-4, IL-10 og M-CSF. M2-makrofager har høy fagocytterende kapasistet, produserer kjemotaktiske signalstoffer og danner ekstracellulære maktrikskomponenter [46, 47]. Inndeling av makrofagfenotyper i M1- eller M2-type har i 44
58 senere tid mottatt kritikk for å være en forenkling, og betegnes av flere som et paradigme som ikke tar høyde for den store heterogeniteten som er sett blant makrofagfenotyper [42, 46, 64]. I litteraturen finnes ulike eksempler på protokoller for differensiering av makrofager til henholdsvis M1 og M2. Slike forskjeller kan variere fra cellemodellen som er brukt, stimuli for differensiering og tidslengde for differensiering. I studier av M1- og M2-makrofager benyttes både museceller og humane celler, men i denne oppgaven er kun protokoller for studier av humane celler diskutert. I studier av M1/M2-differensiering av primære monocytter benyttes ofte GM-CSF og M-CSF som overlevelsesfaktorer for å forhindre apoptose [65] eller i kombinasjon med andre forbindelser som kostimuli [50]. For eksempel så brukes IFNγ og LPS sammen med GM-CSF for å gi opphav til M1, mens M-CSF i kombinasjon med IL-4, IL-10, IL-13 eller en blanding av disse for å gi opphav til M2 [50, 51, 66]. I studier hvor THP-1 monocytter benyttes til M1/M2-differensiering har ulike stoffer blitt benyttet til å differensiere disse til en makrofaglignende fenotype, deriblant 25- dihydroksyvitamin D3, M-CSF og PMA. Det er rapportert at PMA er best egnet til å oppnå THP-1 celler med høyest likhet til makrofager avledet fra primære monocytter [66]. Dette ble bedømt ut ifra høyere fagocyttisk aktivitet og produksjon av IL-1β og TNFα etter stimulering med PMA. I litteraturen finnes det eksempler på at THP-1 monocytter, etter behandling med PMA, har blitt differensiert til M1- og M2-makrofager med henholdsvis LPS og IFNγ eller IL-4 alene eller sammen med IL-13 [52, 60, 67]. Ulike PMA-konsentrasjoner (5-93 ng/ml) er blitt benyttet til M1/M2-differensiering av THP-1 celler. Park et al. [52] stimulerte THP-1 celler med ulike konsentrasjoner av PMA i 48 timer med hensikt om å undersøke hvilken PMA-konsentrasjon som ga optimal respons til svake sekundære stimuli. I den studien var behandling med 10 ng/ml LPS et av de sekundære stimuli som ble undersøkt. Funnene tydet på at 5 ng/ml PMA var nok for å gi THP-1 makrofager som responderte på 10 ng/ml LPS [52]. God respons ble i studien bedømt ut ifra adherens av celler (et tegn på makrofagdifferensiering av monocytter), uttrykk av makrofagmarkøren CD14, og sekresjon av TNFα og IL-8. Sammenlignet med normale somatiske celler (f. eks, primære monocytter) så har THP-1 monocytter en homogen genetisk bakgrunn [66]. Disse cellene er isolert fra blodet til en ett år 45
59 gammel gutt med akutt monocyttisk leukemi [53]. I de tilfeller der man ønsker å studere effekter som er genotypiske, vil primære monocytter være mer egnet da det finnes stor individuell genetisk variasjon. En fordel med THP-1 celler i dette henseende er at den homogene genetiske bakgrunnen vil gjøre det lettere å reprodusere funn, og donorvariabilitet kan elimineres [66, 68]. I denne oppgaven ble det observert at morfologien til THP-1 cellene behandlet med enten LPS og IFNγ eller IL-4 var ulik. Det ble sett at THP-1 celler behandlet med LPS og IFNγ hadde en mer forgrenet form, mens de IL-4-behandlede cellene var mer sirkulære og morfologisk like kontrollmakrofagene som kun var behandlet med PMA. For å skille mellom ulike makrofagfenotyper studeres uttrykk av markører og cytokinproduksjon i tillegg morfologi [50]. Den observerte forskjellen i morfologien til makrofagene i dette arbeidet er i overensstemmelse med funn gjort av andre grupper, der primære humane monocytter har blitt stimulert med ulike stoffer for differensiering, deriblant LPS og IFNγ eller IL-4 [69, 70]. I litteraturen er det foreslått en rekke markører for identifisering av M1- og M2-makrofager, som CD40, CD64, CD80 og CXCL11 for M1 og CD163, CD180, CD206 og TREM2 for M2 [46, 54, 71-73]. Markører som ble benyttet i arbeidet presentert her var CD80 for M1 og CD163 for M2 i cellelysater. Det er stor variasjon i protokoller som benyttes for differensiering av THP-1 og primære humane monocytter, og det er vist at uttrykk av markører og cytokiner in vitro er sterkt avhengig av differensieringsmetoden som brukes [50]. I arbeidet under denne oppgaven så CD80 ut til å være en god markør, da uttrykk av denne markøren utelukkende ble sett i M1-lysater. På den andre siden ga CD163 ikke klare resultater, ettersom denne markøren både ga svake bånd og ble sett i lysat både fra M1 og kontrollmakrofager kun behandlet med PMA. I tillegg kan sekrerte markører (f. eks. scd80[74] og scd163[75]) måles i kondisjonert medium ved ELISA og dette kunne vært en fordel i dette arbeidet for å styrke bevisene om M1- og M2-identifisering. Produksjon av cytokinene TNFα og IL-6 eller TGF-β1 og IL-10 kan også brukes som markører for henholdsvis M1 eller M2 [76]. Måling av disse cytokinene i kondisjonerte medier ville muligens ha gitt en ytterligere bekreftelse på om M1- og M2-differensiering var oppnådd. Måling av mrna av M1/M2-markører kunne også vært et alternativ for å undersøke grad av differensiering, og eventuelt gi økt styrke til M1- og M2-identifisering, men på grunn av lite materiale (lysater) ville dette ha krevd ytterligere forsøk. 46
60 I oppgaven er det benyttet to metoder for differensiering til M1- og M2-makrofager, hvor metode 1 er basert på protokoll fra Park et al. [52] og Chanput et al. [55], mens metode 2 er basert på metode brukt av Halvorsen et al. [60]. Hovedforskjellene mellom disse metodene er konsentrasjon og tid for PMA-stimulering (5 ng/ml i 48 timer versus 62 ng/ml i 6 timer), samt konsentrasjon av LPS (1 μg/ml i metode 1 versus 10 ng/ml i metode 2). Med utgangspunkt i Park et al. sin studie [52] kunne det vært hensiktsmessig å forsøke å stimulere THP-1 celler med lav dose PMA og 10 ng/ml fremfor 1 μg/ml LPS, ettersom deres studie indikerte at en så lav konsentrasjon av sekundært stimuli (i.e. LPS) var tilstrekkelig for differensiering. Etter at makrofager er stimulert med LPS så aktiveres transkripsjonsfaktoren nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), et proteinkompleks som kontrollerer transkripsjon av en rekke gener som regulerer inflammasjon, celleproliferasjon, og -overlevelse [77]. Det har blitt vist av Takashibi et al. at THP-1 celler forbehandlet med PMA før stimulering med LPS hadde en høyere sekresjon av TNFα, sammenlignet med celler som ikke var forbehandlet med PMA [78]. I studien ble det målt økte nivåer av NF-κB i cytoplasma etter forbehandling. PMA aktiverer protein kinase C signaleringsveien og fører til inhibering av celleproliferasjon og forårsaker differensiering til makrofager [79]. Det ble foreslått av Takashibi et al. at PMA gir en akkumulering av NF-κB i cytoplasma, og dermed en økt følsomhet for LPS. PMA-konsentrasjonen som ble benyttet var 124 ng/ml i 24 timer. CD80 er i denne oppgaven blitt brukt som M1-markør og transkripsjon av CD80-genet igangsettes av NF-κB [80]. I denne oppgaven ble det sett høyere uttrykk av CD80 i THP-1 makrofager differensiert etter metode 1, sammenlignet med tilsvarende celler differensiert etter metode 2. Det skal også bemerkes at det var lavere mengde totalprotein som ble applisert per brønn etter differensiering med metode 1 (10 μg) enn metode 2 (15 μg), da metode 1 ga færre adherente celler enn metode 2. Det kan derfor antas at den virkelige forskjellen i uttrykk av CD80 er enda høyere enn det som ble observert ved immunoblotting når man tar til høyde for forskjell i mengde applisert totalprotein. Uttrykket av M2-markøren CD163 var derimot ikke like avklarende. Etter differensiering med metode 1 var uttrykket av CD163 meget lavt, sett ved et svakt bånd etter immunoblotting. Etter differensiering med metode 2 på den andre siden var uttrykket av CD163 fraværende i de antatte M2-makrofagene etter behandling med 25 ng/ml IL-4 i 72 timer, mens uttrykk ble sett både etter 18 og 72 timer i kontrollmakrofager samt i M1-47
61 makrofager. Overraskende nok så var det høyeste uttrykket av CD163 å se i kontrollmakrofager etter 72 timer. Selv om CD163 betegnes og brukes som en M2-markør er det av andre rapportert at IL-4 ikke påvirker uttrykket av eller til og med undertrykker CD163 i primære humane monocytter in vitro [48, 61]. Halvorsen et al. [60] benyttet IL-13 i tillegg til IL-4 for å differensiere THP-1 celler til M2- makrofager, mens kun IL-4 har blitt brukt i denne oppgaven. Det har vært antatt at begge disse cytokinene har lignende funksjoner, siden IL-4 og IL-13 har en felles reseptorsignaleringsvei (IL-4Rα/IL13Rα1/STAT6) [81]. IL-13 kan i tillegg også igangsette signalering via reseptoren IL13Rα2, som utelukkende binder IL-13. Aktivering av denne reseptoren fører til økt TGF-β1 produksjon i makrofager, og leder til økt deponering av kollagen [82]. I studier av low-density lipoprotein receptor knock-out (LDLR -/- ) mus gjort av Fichtner-Feigl et al. ble det vist at injeksjon av IL-13 ga økt kollageninnhold i aterosklerotiske lesjoner, lavere makrofaginnhold, samt økt M2:M1 ratio i lesjonen [83]. Denne masteroppgaven er kun basert på in vitro eksperimenter og analysemetodene som er brukt i denne oppgaven gir ingen informasjon om kollagensyntese i M1- og M2-makrofager, likevel så indikerer funnene til Fichtner-Feigl et al. at det vil være fysiologisk mer nærliggende å differensiere THP-1 celler til M2 ved kostimulering med IL-4 og IL-13, med hensyn til å studere makrofagers pro- og antiinflammatoriske effekt på progresjon av aterosklerose lesjoner. Det er et generelt ønske ved utforming av in vitro modeller at de skal speile in vivo betingelser i den grad det lar seg gjøre, for å gi best mulig forklaring til faktiske in vivo hendelser. Med dette argumentet kunne det vært en fordel at IL-13 ble benyttet i tillegg til IL-4 ved M2-differensiering av THP-1 monocytter i denne masteroppgaven. I dette arbeidet ble det observert at M1-makrofager hadde litt høyere uttrykk av moden form av legumain (36 kda) enn M2-makrofager sett ved immunoblotting. Samtidig viste enzymaktivitetsmålingene med peptidsubstratet Z-Ala-Ala-Asn-AMC at AEP-aktiviteten var høyere i M2-lysater. Inkubering av makrofaglysater (M1 og M2) med E-64 som er en generell hemmer av cysteincathepsiner viste at en stor del av AEP-aktiviteten i både M1- og M2-lysater ble hemmet av E-64. Solberg et al. (2015) har nylig vist at omtrent 70 % av AEPaktiviteten i PMA- og TNFα-stimulerte THP-1 monocytter ble hemmet av E-64 [51]. Solberg et al. foreslo at det kan være andre cysteincathepsin(er) til stede i disse cellene, i tillegg til legumain, som kan spalte Z-Ala-Ala-Asn-AMC [51]. At AEP-aktivitet i THP-1 makrofager reduseres etter tilsetning av E-64 ble også observert av tidligere masterstudent i farmasi Inass 48
62 Elyouncha (2015). Det at PMA-behandlede THP-1 makrofager har en AEP-aktivitet som ikke skyldes legumain har i dette arbeidet blitt bekreftet. Interessant nok, ble det i den første legumain knock-out (KO)-studien fra 2003 [18] målt lav AEP-aktivitet i vevshomogenat fra nyre med peptidsubstratet Z-Ala-Ala-Asn-AMC i både villtype og KO-mus, og AEPaktiviteten i KO-mus ble vist å være < 10 % av legumain +/+ aktiviteten [18]. For å få et inntrykk av hvor mye ekte legumainaktivitet som det faktisk er i M1 og M2 ble lysater tilsatt E-64 som hemmer cysteincathepsiner, men ikke legumain [15] før måling av AEP-aktivitet. Resultatene viste at det en var større andel ukjent AEP i M2 enn i M1. Dette kan være en forklaring på hvorfor det ble sett høyere uttrykk av moden legumain (36 kda) i M1, mens AEP-aktiviteten var høyere i M2. Tilstedeværelse av andre cysteincathepsin(er) utfordrer den antatte spesifisiteten til peptidsubstratet i metoden for måling av legumainaktivitet [57]. Samtidig bør grad av endogen hemming av legumain i M1-lysater sammenlignes med M2-lysater og tas med i vurderingen. Potensielt kan mer endogen hemming i M1-lysater sammenlignet med M2-lysater være årsaken til lavere legumainaktivitet i M1- enn M2-lysater etter tilsetning av E-64. Uttrykk at den mest potente endogene legumainhemmeren cystatin E/M er undersøkt i PMA-stimulerte THP-1 celler [51], men uttrykk av cystatin E/M i M1- versus M2-makrofager er ukjent. Det ble observert høyere uttrykk av prolegumain (56 kda) i både M1- og M2-makrofager differensiert ved begge metodene brukt i denne oppgaven. ELISA-målinger av sekrert legumain i kondisjonerte medier støttet det som ble sett ved immunoblotting. Kondisjonerte medier fra M1 hadde en betraktelig høyere konsentrasjon av legumain enn medier fra M2. Lignende observasjon ble gjort av Solberg et al. i kondisjonerte medier fra M1- og M2- differensierte primære humane monocytter stimulert med henholdsvis GM-CSF og IFNγ (M1) eller M-CSF (M2) [51]. En form for legumain på 75 kda ble observert i M2-makrofager differensiert etter metode 2. Denne 75 kda-formen er en ukjent legumainform som ikke er beskrevet tidligere. Zhao et al. har demonstrert at legumain kan ha en ligaseaktivitet, i tillegg til en proteaseaktivitet [84]. En mulig forklaring på tilstedeværelsen av denne 75 kda-formen kan være at den er dannet som følge av ligering og/eller homodimerisering av moden legumain (36 kda). De reduserende betingelsene under gelelektroforesen vil derimot bryte opp en ikke-kovalent homodimer legumain, og ligaseaktivitet som Zhao et al. observerte ble sett ved bruk av renset 49
63 rekombinant legumain, og det er ikke kjent om ligaseaktiviteten eksisterer i levende celler eller in vivo. Av disse grunner så er trolig ikke 75-kDa formen av legumain en homodimer av moden legumain. I tillegg til en 75 kda legumainform, ble det også sett en form med molekylvekt noe større enn prolegumain ( 56 kda). Denne formen ble sett kraftigst uttrykt i M2-makrofager, men den ble også observert i M1-makrofager og kontrollmakrofager. Human prolegumain har et signalpeptid som frigjøres under cellulær transport [7] og det er mulig at denne legumainformen var prolegumain med signaleringspeptidet. Legumain har et Arg-Gly-Asp 120 (RGD) motiv som kan binde til αvβ3- integrinreseptoren på cellemembranen, og ved binding til denne reseptoren øker legumain sitt ph-optimum fra 5,5 til 6, og det er i tillegg kjent at prolegumain er stabil ved nøytral ph [13]. Det kan være fristende å foreslå at sekrert prolegumain kan binde til αvβ3- integrinreseptoren, hvor proteasen aktiveres ekstracellulært og potensielt kløyver fibronektin, aktiverer prommp-2 og prosesserer cysteincathepsiner (cathepsin L og cathepsin B) in vivo [85]. Fibronektin er en av hovedbestanddelene i ekstracellulær matriks (ECM)[24] og det er vist at legumain er i stand til å spalte fibronektin in vitro [26]. Spalting av fibronektin ved legumain er bekreftet i denne oppgaven. Det har også nylig blitt vist kolokalisering av legumain og fibronektin i ECM i tumorvev fra mus xenotransplantert med OCI-Ly3 (human cellemodell for diffus storcellet B-celle lymfom) [86]. Samlet kan dette tyde på at legumain kan være aktiv og spalte fibronektin ekstracellulært. I denne oppgaven ble det sett at fibronektin gjennomgår omfattende spaltning og degradering etter inkubering med renset bovint legumain eller lysat fra en stabiltransfektert HEK293 cellelinje som overuttrykker legumain (M38L). M1- og M2-lysater på den annen side viste begrenset spaltning av fibronektin, men M2-lysatene ga tydeligst degradering av fibronektin, sett ved densitometrisk scanning av immunoblott. Det skal her gjøres oppmerksom på at like volumer av M1- og M2-lysater ble brukt ved gelelektroforesen, uten at forskjell i totalproteinkonsentrasjon ble tatt hensyn til. Dette kom av at inhibitorene som ble benyttet kun binder til aktive enzymer, noe som ikke tillot acetonfelling for oppkonsentrering av prøvene og korrigering for forskjell i totalproteinkonsentrasjon. Forskjellen mellom M1 og M2 kan derfor skyldes ulik mengde totalprotein ved elektroforese. 50
64 Det har her blitt vist at både legumain og cysteincathepsiner er aktive i M38L-lysater og degraderer fibronektin, ettersom degradering av fibronektin ble hemmet etter tilsetning av spesifikk inhibitor av legumain (MP-L01*) eller cysteincathepsininhibitor (E-64). Det ble observert omfattende degradering av fibronektin i M38L-lysat tilsatt legumaininhibitoren (MP-L01*) i forhold til makrofaglysatene. Dette kan komme av at legumain kan ha aktivert andre enzymer som kan delta i degradering av fibronektin. Immunoblottene viste også unike degraderingsprodukter av fibronektin som skyldes aktiv legumain, som var fraværende ved hemming med MP-L01*, men ikke E-64. I denne oppgaven ble det i både M1- og M2-makrofager sett uttrykk av en 25 kda legumainform ved deteksjon med kjemiluminescens (ChemiDoc XRS + ). Denne 25 kdaformen ble også observert med renset bovint legumain i kompleks med legumainselektiv probe (MP-L01) og deteksjon med infrarød fluorescens (Odyssey CLx Near-Infrared Imaging System). I M2-makrofager og kontrollmakrofager ble det observert binding av proben til et 25 kda protein, mens antistoff mot legumain ikke ga detekterbare signaler ved denne metoden. Denne tilsynelatende uoverensstemmelse kan forklares med at det er benyttet forskjellige sekundære antistoffer ved de to fremkallingsmetodene. I tillegg så var det samme volumer av lysater, og ikke proteinmengder, som ble tilsatt brønnene på gelen. Dette kom av at inkubatene må inneholde aktiv legumain for å kunne binde MP-L01 [41]. Det var generelt lite totalprotein i makrofaglysatene som ble inkubert med probe, da disse ikke kunne oppkonsentreres ved acetonfelling. Fortynningsgraden av legumainantistoff ved immunoblotting er viktig og kan være et moment som kan forklare uoverensstemmelsen mellom observasjoner gjort ved fremkalling på ChemiDoc XRS + kontra Odyssey CLx Near-Infrared Imaging System. Da det i vårt laboratorium normalt brukes en 1:1000 fortynning av legumainantistoff for deteksjon av legumain i lysater fra celletyper med moderat eller høy legumainuttrykk (f.eks. HEK293 og M38L), kan en 1:200 fortynning som ble benyttet på THP-1 differensierte makrofager anses som relativt høy. Ved fremkalling av immunoblott på Odyssey CLx Near-Infrared Imaging System ble det observert mye uspesifikk binding av antistoff, som kan skyldes den høye antistoffkonsentrasjonen og vanskeliggjorde tolkningen av immunoblottene. MP-L01 proben ble vist å binde til både mellomform (46 kda; M38L-lysat) og moden form (36 kda), i tråd med det som er kjent om aktive former av legumain [7], samt til en 25 kda- 51
65 formen (i både M38L-lysat og renset bovint legumain). Hvorvidt denne 25 kda-formen er aktiv er ukjent, men ettersom proben binder til det aktive setet på legumain [41] er det grunn til å anta at denne legumainformen er aktiv. Det vil også være nærliggende å anta at denne formen er et nedbrytningsprodukt av moden 36 kda-form. Imidlertid er det ukjent hvor den modne formen av legumain eventuelt spaltes for å gi opphav til denne 25 kda-formen, og om denne formen har spesifikke cellulære lokaliseringer og funksjoner. I vårt laboratorium har masterstudent i farmasi Kristina B.B. Larsen (2016) nylig observert internalisering av en 25 kda-form av legumain i HEK293-celler etter inkubering med renset bovint legumain med og uten bundet MP-L01 probe, hvilket bekrefter funnet i denne oppgaven. Andre funn støtter også våre observasjoner om en 25 kda-form av legumain. En form av legumain på 24 kda har tidligere blitt observert i primære makrofager isolert fra beinmarg fra mus etter stimulering med IL-4 og IL-10 [87]. I studien ble differensierte primære makrofager stimulert med interleukiner og samtidig inkubert med en fluorescerende aktivitetsbasert legumainprobe (LE28). Derimot ble ikke 24 kda-formen av legumain observert i lysater fra celler behandlet på samme måte. Forfatterne i den artikkelen foreslo at betingelser ved selve lyseringsprosessen kan ha forstyrret observasjonen av 24 kda-formen. En 26 kda-form av legumain har også tidligere blitt sett av Mattock et al. i stabile og ustabile områder fra karotisplakk [27]. I 2006 ble en studie utført av Papaspyridonos et al. der forskjell i uttrykk av ulike kandidatgener i ustabile og stabile områder fra aterosklerotiske plakk ble undersøkt [33]. Her ble det tatt prøver etter karotis-endarterektomi (CEA) fra 46 pasienter, og det ble funnet ved kvantitativ polymerase chain reaction (PCR) at 18 gener var oppregulert i ustabile områder av plakk, deriblant proteasene legumain, MMP-9 og cathepsin B. Videre viste Mattock et al. at uttrykk og aktivitet av legumain var høyere i ustabile områder av plakk fra CEA, sammenlignet med stabile områder [27]. Dessuten viste Papaspyridonos et al. at legumain mrna var kolokalisert med den generelle makrofagmarkøren CD68 i skulderregionen av aterosklerotiske lesjoner, og det ble foreslått at proteasen kan ha en mulig funksjon i plakkdestabilisering. Andre har vist at M1- og M2-makrofager er ulikt fordelt i humane plakk, med økt M1 i ustabile områder og M2 i mer stabile områder [59], men 52
66 derimot har det til nå ikke blitt demonstrert at legumain uttrykkes hovedsakelig av M1- makrofager in vivo. I denne oppgaven har effekt av atorvastatin under M1- og M2-differensiering blitt undersøkt. Det er tidligere vist gjennom store kliniske studier at statiner har flere gunstige effekter som er uavhengig av deres kolesterolsenkende egenskaper; såkalte pleiotrope effekter [35]. Med hensyn til aterosklerose er det foreslått flere mekanismer for slike pleiotrope effekter, deriblant modulering av endotelfunksjon, reduksjon av ROS, reduksjon av plakkinflammasjon og plakkstabilisering [34]. Wang et al. utførte en randomisert dobbelblind studie med 40 deltagere som ble behandlet med atorvastatin, og det ble sett en reduksjon av legumain mrna i sirkulerende monocytter fra deltagerne [38]. Det har i arbeidet med denne oppgaven blitt observert at M1- og M2-makrofager ble mindre adherente ved behandling med økende konsentrasjoner atorvastatin. Det ble også observert nedgang i uttrykk av moden legumain i både M1 og M2, samt uttalt redusert prosessering av prolegumain i M2. I tråd med observasjoner gjort i dette arbeidet har Solberg et al. rapportert en doseavhengig reduksjon i uttrykk av moden legumain og medfølgende redusert prosessering av prolegumain i M2-differensierte humane primære monocytter [51]. Smith et al. behandlet humane myotuber med simvastatin (0-40 µm) og observerte redusert opptak av glukose og redusert prosessering av prolegumain (simvastatin, 30 µm) [88]. Smith et al. forklarer disse observasjonene med at det reduserte opptaket av glukose kan lede til redusert ATP-produksjon med nedsatt aktivitet av lysosomal H + -ATPase og økt ph i lysosomer som konsekvens. Ettersom legumain autoaktiveres ved sur ph kan økt lysosomal ph lede til redusert prosessering av prolegumain til moden legumain. I denne oppgaven ble det sett at den tidligere nevnte 75 kda-formen for legumain i M2 ble påvirket av atorvastatin, da båndintensiteten på immunoblottet ble kraftig redusert etter behandling med 10 µm atorvastatin og var fullstendig borte etter behandling med 25 µm atorvastatin. Hvordan atorvastatin påvirker uttrykket av denne legumainformen er ukjent. Zhang et al. har vist at primære monocytter isolert fra pasienter behandlet med atorvastatin (20-40 mg daglig) i en måned inneholdt økte nivåer av M2-markører (CD206, IL-10 og CCL18) [61]. Forskerne har også differensiert humane monocytter til M2 med IL-4 og behandlet disse med atorvastatin (5, 10 og 20 µm) i begynnelsen av 53
67 differensieringsprosessen. Her ble det sett at uttrykket av CD206 økte etter IL-4 behandling, og at denne effekten ble forsterket av atorvastatin på en doseavhengig måte. Derimot ble uttrykket av CD163, som også ble undersøkt, redusert med økende doser atorvastatin. Zhang et al. sine resultater med hensyn til CD163 er i tråd med observasjonene som ble gjort av denne angivelige M2-markøren under arbeidet med denne oppgaven. Uttrykk av CD163 ble sett i både M1 og M2 og høyest uttrykk av denne markøren ble gjort i kontrollmakrofager. Mangel på en unik positiv markør for M2 i denne oppgaven, gir svakere grunnlag verifisering av M2-differensiering, enn for M1 nevnt over. Med hensyn til påvirkning av atorvastatin (1-10 µm) på morfologi utviste behandlede M1- makrofager en morfologi som er karakteristisk for denne fenotypen [69, 70]. Derimot så utviste M1-makrofager behandlet med 25 µm og 50 µm (ikke vist) atorvastatin en hovedsakelig sirkulær morfologi, som er typisk for M2-makrofager [69, 70]. På basis av dette kan det være fristende å spekulere om denne endrede morfologien av M1-makrofager behandlet med høye doser atorvastatin (25 µm og 50 µm) kan være en følge av at M1 har blitt dreiet mot en M2-fenotype. I tillegg har andre grupper demonstrert at fullt differensierte M1- og M2-makrofager kan skifte fenotype som respons til ulike stimuli. Makrofager stimulert med IFNγ (M1) ble endret til en M2-fenotype etter behandling med kondisjonert medium fra M2-makrofager stimulert med IL-4, og makrofager stimulert med deksametason (M2) ble endret til en M1-fenotype etter behandling med LPS [70, 89]. Det ble i denne oppgaven sett at legumain er nødvendig for prosessering av procathepsin L fra single-chain til two-chain formen av cathepsin L. Uttrykket av two-chain -cathepsin L ble mindre i både M1- og M2-makrofager etter behandling med 25 μm atorvastatin.. Denne nedgangen sammenfalt med nedgang i uttrykk av aktiv legumain. Disse funnene er i tråd med tidligere publikasjoner [18, 62]. Som nevnt over er det sett redusert opptak av glukose i humane myotuber behandlet med simvastatin [88], og interessant nok er det dessuten rapportert at økt konsentrasjon av glukose i dyrkningsmediet til humane monocytter gir nedregulering av cathepsin B, D, L og S [90]. Konsentrasjoner av atorvastatin (mikromolar) som ble benyttet i denne oppgaven er i samme område som statinkonsentrasjoner brukt i andre in vitro studier [51, 88], men over plasmakonsentrasjonsområdet (nanomolar) som oppnås in vivo. Celleforsøk med atorvastatin in vitro spenner over en kort tidsperiode (dager), sammenlignet men langsiktig terapeutisk 54
68 bruk. Det kan dermed argumenteres for at det er nødvendig med høyere atorvastatinkonsentrasjoner for å kunne detektere cellulære responser. M1-makrofager er assosiert med inflammasjon, produserer inflammatoriske cytokiner som TNFα og IL-6 og øker produksjonen av ROS, som opprettholder utviklingen av aterosklerotiske lesjoner [49, 91]. Videre er M1-makrofager hovedsakelig lokalisert til ustabile områder av aterosklerotiske plakk [59] og det har blitt vist at proteolyse bidrar til utviklingen av aterosklerose ved for eksempel plakkdestabilisering, degradering av ECM og infiltrering av leukocytter i blodåreveggen [92]. I tillegg er det sett at renset prolegumain utviser kjemotaktiske effekter på humane monocytter in vivo, likeledes med denaturert legumain, som indikerer at denne effekten er uavhengig av legumain sin proteaseaktivitet [28]. Disse ovenfor nevnte publiserte funnene nevnes med hensikt om å understreke hvordan proinflammatoriske makrofager (M1) og legumain er implisert i aterosklerose. I denne oppgaven har det blitt etablert metode for M1-differensiering av THP-1 monocytter, og proteinuttrykk, sekresjon og aktivitet av legumain fra disse cellene ble redusert etter behandling med atorvastatin. Disse funnene kan være med på å forklare noe at statiners pleiotrope effekter. 55
69 6 Konklusjon Metode for differensiering av THP-1 monocytter til M1- og M2-makrofager ble etablert THP-1 celler stimulert med høy (62 ng/ml) PMA-konsentrasjon er mer adherente enn celler stimulert med lav (5 ng/ml) PMA-konsentrasjon M1- og M2-makrofager har ulik morfologi M1- og M2-differensierte THP-1 celler har ulikt uttrykk av legumain M2-makrofager uttrykker en 75 kda-form av legumain som ikke sees i M1- makrofager M1-makrofager sekrerer (10 x) mer legumain enn M2-makrofager AEP-aktivitet er høyere i M2-makrofager enn M1-makrofager M2-lysater har større evne til å degradere fibronektin enn M1-lysater Behandling med atorvastatin 10μM under differensiering fører til nedregulering i uttrykk av moden legumain (36 kda) i M1-makrofager Behandling med atorvastatin 10 μm under differensiering fører til redusert prosessering av prolegumain (56 kda) og nedregulering av moden legumain (36 kda) i M2-makrofager M1-makrofager uttrykker mer cathepsin L ( two-chain -form og singlechain -form) enn M2-makrofager ved behandling med atorvastatin under makrofagdifferensieringen? 56
70 Litteraturliste 1. Turk, B., Targeting proteases: successes, failures and future prospects. Nat Rev Drug Discov, (9): p Rawlings, N.D., A.J. Barrett, and R. Finn, Twenty years of the MEROPS database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res, (D1): p. D Shutov, A.D., N.L. Do, and I.A. Vaintraub, [Purification and partial characterization of protease B from germinating vetch seeds]. Biokhimiia, (5): p Ruppel, A., et al., Sera of Schistosoma japonicum-infected patients cross-react with diagnostic 31/32 kd proteins of S. mansoni. Clin Exp Immunol, (2): p Chen, J.M., et al., Cloning, isolation, and characterization of mammalian legumain, an asparaginyl endopeptidase. J Biol Chem, (12): p Chen, J.M., et al., Identification of the active site of legumain links it to caspases, clostripain and gingipains in a new clan of cysteine endopeptidases. FEBS Lett, (3): p Dall, E. and H. Brandstetter, Structure and function of legumain in health and disease. Biochimie, : p Halfon, S., et al., Autocatalytic activation of human legumain at aspartic acid residues. FEBS Lett, (1-2): p Yamane, T., et al., Legumain from bovine kidney: its purification, molecular cloning, immunohistochemical localization and degradation of annexin II and vitamin D- binding protein. Biochim Biophys Acta, (1): p Haugen, M.H., et al., Nuclear legumain activity in colorectal cancer. PLoS One, (1): p. e Smith, R., et al., Intra- and extracellular regulation of activity and processing of legumain by cystatin E/M. Biochimie, (12): p Li, D.N., et al., Multistep autoactivation of asparaginyl endopeptidase in vitro and in vivo. J Biol Chem, (40): p Dall, E. and H. Brandstetter, Mechanistic and structural studies on legumain explain its zymogenicity, distinct activation pathways, and regulation. Proc Natl Acad Sci U S A, (27): p Alvarez-Fernandez, M., et al., Inhibition of mammalian legumain by some cystatins is due to a novel second reactive site. J Biol Chem, (27): p Barrett, A.J., et al., L-trans-Epoxysuccinyl-leucylamido(4-guanidino)butane (E-64) and its analogues as inhibitors of cysteine proteinases including cathepsins B, H and L. Biochem J, (1): p Manoury, B., et al., An asparaginyl endopeptidase processes a microbial antigen for class II MHC presentation. Nature, (6712): p Sepulveda, F.E., et al., Critical role for asparagine endopeptidase in endocytic Tolllike receptor signaling in dendritic cells. Immunity, (5): p Shirahama-Noda, K., et al., Biosynthetic processing of cathepsins and lysosomal degradation are abolished in asparaginyl endopeptidase-deficient mice. J Biol Chem, (35): p Miller, G., et al., Asparagine endopeptidase is required for normal kidney physiology and homeostasis. Faseb j, (5): p
71 20. Turk, V., et al., Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics, (1): p Barrett, A.J. and H. Kirschke, Cathepsin B, Cathepsin H, and cathepsin L. Methods Enzymol, Pt C: p Lutgens, S.P., et al., Cathepsin cysteine proteases in cardiovascular disease. Faseb j, (12): p Chen, J.M., et al., Activation of progelatinase A by mammalian legumain, a recently discovered cysteine proteinase. Biol Chem, (5): p Plow, E.F., et al., Ligand binding to integrins. J Biol Chem, (29): p Pankov, R. and K.M. Yamada, Fibronectin at a glance. J Cell Sci, (Pt 20): p Morita, Y., et al., Legumain/asparaginyl endopeptidase controls extracellular matrix remodeling through the degradation of fibronectin in mouse renal proximal tubular cells. FEBS Lett, (7): p Mattock, K.L., et al., Legumain and cathepsin-l expression in human unstable carotid plaque. Atherosclerosis, (1): p Clerin, V., et al., Expression of the cysteine protease legumain in vascular lesions and functional implications in atherogenesis. Atherosclerosis, (1): p Mohamed, M.M. and B.F. Sloane, Cysteine cathepsins: multifunctional enzymes in cancer. Nat Rev Cancer, (10): p Liu, C., et al., Overexpression of legumain in tumors is significant for invasion/metastasis and a candidate enzymatic target for prodrug therapy. Cancer Res, (11): p Wu, W., et al., Targeting cell-impermeable prodrug activation to tumor microenvironment eradicates multiple drug-resistant neoplasms. Cancer Res, (2): p Luo, Y., et al., Targeting tumor-associated macrophages as a novel strategy against breast cancer. J Clin Invest, (8): p Papaspyridonos, M., et al., Novel candidate genes in unstable areas of human atherosclerotic plaques. Arterioscler Thromb Vasc Biol, (8): p Liao, J.K. and U. Laufs, Pleiotropic effects of statins. Annu Rev Pharmacol Toxicol, : p Liao, J.K., Beyond lipid lowering: the role of statins in vascular protection. Int J Cardiol, (1): p Blum, A. and R. Shamburek, The pleiotropic effects of statins on endothelial function, vascular inflammation, immunomodulation and thrombogenesis. Atherosclerosis, (2): p Dichtl, W., et al., HMG-CoA reductase inhibitors regulate inflammatory transcription factors in human endothelial and vascular smooth muscle cells. Arterioscler Thromb Vasc Biol, (1): p Wang, Z.H., et al., Pleiotropic effects of atorvastatin on monocytes in atherosclerotic patients. J Clin Pharmacol, (3): p Abrahamson, M., Cystatins. Methods Enzymol, : p Varughese, K.I., et al., Crystal structure of a papain-e-64 complex. Biochemistry, (3): p Poreba, M., et al., Counter Selection Substrate Library Strategy for Developing Specific Protease Substrates and Probes. Cell Chem Biol,
72 42. Gordon, S. and P.R. Taylor, Monocyte and macrophage heterogeneity. Nat Rev Immunol, (12): p Das, A., et al., Monocyte and macrophage plasticity in tissue repair and regeneration. Am J Pathol, (10): p Mosser, D.M. and J.P. Edwards, Exploring the full spectrum of macrophage activation. Nat Rev Immunol, (12): p Davies, L.C., et al., Tissue-resident macrophages. Nat Immunol, (10): p Martinez, F.O. and S. Gordon, The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep, : p Shalhoub, J., et al., Innate immunity and monocyte-macrophage activation in atherosclerosis. J Inflamm (Lond), : p Barros, M.H., et al., Macrophage polarisation: an immunohistochemical approach for identifying M1 and M2 macrophages. PLoS One, (11): p. e Gordon, S., Alternative activation of macrophages. Nat Rev Immunol, (1): p Vogel, D.Y., et al., Human macrophage polarization in vitro: maturation and activation methods compared. Immunobiology, (9): p Solberg, R., et al., Legumain expression, activity and secretion are increased during monocyte-to-macrophage differentiation and inhibited by atorvastatin. Biol Chem, (1): p Park, E.K., et al., Optimized THP-1 differentiation is required for the detection of responses to weak stimuli. Inflamm Res, (1): p Tsuchiya, S., et al., Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int J Cancer, (2): p Badylak, S.F., et al., Macrophage phenotype as a determinant of biologic scaffold remodeling. Tissue Eng Part A, (11): p Chanput, W., et al., Characterization of polarized THP-1 macrophages and polarizing ability of LPS and food compounds. Food Funct, (2): p Bradford, M.M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, : p Johansen, H.T., C.G. Knight, and A.J. Barrett, Colorimetric and fluorimetric microplate assays for legumain and a staining reaction for detection of the enzyme after electrophoresis. Anal Biochem, (2): p Towbin, H., T. Staehelin, and J. Gordon, Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A, (9): p Stoger, J.L., et al., Distribution of macrophage polarization markers in human atherosclerosis. Atherosclerosis, (2): p Halvorsen, B., et al., Increased expression of NAMPT in PBMC from patients with acute coronary syndrome and in inflammatory M1 macrophages. Atherosclerosis, (1): p Zhang, O. and J. Zhang, Atorvastatin promotes human monocyte differentiation toward alternative M2 macrophages through p38 mitogen-activated protein kinasedependent peroxisome proliferator-activated receptor gamma activation. Int Immunopharmacol, (1): p Li, W., et al., Cathepsin L is significantly associated with apoptosis and plaque destabilization in human atherosclerosis. Atherosclerosis, (1): p
73 63. Moon, M.L., L.K. McNeil, and G.G. Freund, Macrophages make me sick: how macrophage activation states influence sickness behavior. Psychoneuroendocrinology, (10): p Murray, P.J., et al., Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity, (1): p Kohro, T., et al., A comparison of differences in the gene expression profiles of phorbol 12-myristate 13-acetate differentiated THP-1 cells and human monocytederived macrophage. J Atheroscler Thromb, (2): p Chanput, W., J.J. Mes, and H.J. Wichers, THP-1 cell line: an in vitro cell model for immune modulation approach. Int Immunopharmacol, (1): p Genin, M., et al., M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer, : p Schildberger, A., et al., Monocytes, peripheral blood mononuclear cells, and THP-1 cells exhibit different cytokine expression patterns following stimulation with lipopolysaccharide. Mediators Inflamm, : p Jaguin, M., et al., Polarization profiles of human M-CSF-generated macrophages and comparison of M1-markers in classically activated macrophages from GM-CSF and M-CSF origin. Cell Immunol, (1): p Porcheray, F., et al., Macrophage activation switching: an asset for the resolution of inflammation. Clin Exp Immunol, (3): p Mantovani, A., et al., The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol, (12): p Martinez, F.O., et al., Transcriptional profiling of the human monocyte-tomacrophage differentiation and polarization: new molecules and patterns of gene expression. J Immunol, (10): p Glim, J.E., et al., Platelet derived growth factor-cc secreted by M2 macrophages induces alpha-smooth muscle actin expression by dermal and gingival fibroblasts. Immunobiology, (6): p Kakoulidou, M., et al., Soluble costimulatory factors scd28, scd80, scd86 and scd152 in relation to other markers of immune activation in patients with myasthenia gravis. J Neuroimmunol, (1-2): p Kazankov, K., et al., Soluble CD163, a macrophage activation marker, is independently associated with fibrosis in patients with chronic viral hepatitis B and C. Hepatology, (2): p Durafourt, B.A., et al., Comparison of polarization properties of human adult microglia and blood-derived macrophages. Glia, (5): p Sharif, O., et al., Transcriptional profiling of the LPS induced NF-κB response in macrophages. BMC Immunology, : p Takashiba, S., et al., Differentiation of monocytes to macrophages primes cells for lipopolysaccharide stimulation via accumulation of cytoplasmic nuclear factor kappab. Infect Immun, (11): p Schwende, H., et al., Differences in the state of differentiation of THP-1 cells induced by phorbol ester and 1,25-dihydroxyvitamin D3. J Leukoc Biol, (4): p Niu, H., G. Cattoretti, and R. Dalla-Favera, BCL6 controls the expression of the B7-1/CD80 costimulatory receptor in germinal center B cells. J Exp Med, (2): p Kuperman, D.A. and R.P. Schleimer, Interleukin-4, interleukin-13, signal transducer and activator of transcription factor 6, and allergic asthma. Curr Mol Med, (5): p
74 82. Fichtner-Feigl, S., et al., IL-13 signaling through the IL-13alpha2 receptor is involved in induction of TGF-beta1 production and fibrosis. Nat Med, (1): p Cardilo-Reis, L., et al., Interleukin-13 protects from atherosclerosis and modulates plaque composition by skewing the macrophage phenotype. EMBO Mol Med, (10): p Zhao, L., et al., Structural analysis of asparaginyl endopeptidase reveals the activation mechanism and a reversible intermediate maturation stage. Cell Res, (3): p Smith, R.L., Regulation of legumain in human cells. 2014, (Doctoral thesis) University of Oslo. 86. Shen, L., et al., M2 tumour-associated macrophages contribute to tumour progression via legumain remodelling the extracellular matrix in diffuse large B cell lymphoma. Scientific Reports, : p Edgington, L.E., et al., Functional imaging of legumain in cancer using a new quenched activity-based probe. J Am Chem Soc, (1): p Smith, R., et al., Simvastatin Inhibits Glucose Metabolism and Legumain Activity in Human Myotubes. PLoS ONE, (1): p. e Khallou-Laschet, J., et al., Macrophage plasticity in experimental atherosclerosis. PLoS One, (1): p. e Moheimani, F., et al., Inhibition of lysosomal function in macrophages incubated with elevated glucose concentrations: a potential contributory factor in diabetesassociated atherosclerosis. Atherosclerosis, (1): p Ley, K., Y.I. Miller, and C.C. Hedrick, Monocyte and macrophage dynamics during atherogenesis. Arterioscler Thromb Vasc Biol, (7): p Liu, J., et al., Lysosomal cysteine proteases in atherosclerosis. Arterioscler Thromb Vasc Biol, (8): p
75 Vedlegg A: Oppskrift for løsninger xPBS: 14,4 g Natriumdihydrogenfosfat NaH 2PO 4 H 2O 2,0 g KH 2PO 4 80 g NaCl 2 g KCl Destillert H 2O ad 1 L Justeres til ph 7,2 med HCl Fortynnes til 1:10 med dh 2O før bruk 1.2 Lysisbuffer (ph 5,8): 14,71 g Natriumcitrat Natriumcitrat 0,19 g Dinatrium-EDTA 5 g N-oktyl-β-D-glukopyranosid Justeres til ph 5,8 med HCl Destillert H 2O ad 500 ml 1.3 Legumain-assaybuffer (ph 5,8): 3,30 g Sitronsyre 21,54 g Dinatriumhydrogenfosfat 0,37 g Dinatrium-EDTA 0,1 g CHAPS Destillert H 2O ad 1,0 L 200 µl 200 mm DTT tilsettes 40 ml legumain-assaybuffer rett før bruk 1.4 Legumain-substratoppløsning (34 µm): 200 µl stock -løsning med Ala-Ala-Asn-AMC (2,5 mm i DMSO) 14,5 ml legumain-assaybuffer (ph 5,8) 1.5 Fortynning av E-64 til 10 µm bruksløsning: 10 µl stock -løsning med E-64 (10 mm) fortynnes til 10 µm med destillert H 2O 1.6 Elektroforesebuffer: 40 ml NuPAGE MOPS SDS Running Buffer (20X) Destillert H 2O ad 0,8 L 500 µl NuPAGE Antioxidant tilsettes direkte i elektroforesekaret med elektroforesebuffe 1.7 Blottebuffer: 400 ml 10X Tris/Glycine-buffer (Bio-Rad) 800 ml Metanol (MeOH) Destillert H 2O ad 2,8 L X T-TBS: 48,4 g Tris 220,2 g NaCl 20 ml Tween 20 Justert til ph 7,5 med HCl Destillert H 2O ad 2,0 L Fortynnes 1:10 med destillert H 2O før bruk 62
 Foredrag holdt av masterstudent")


76 Vedlegg B: Foredrag-Norsk Selskap for Farmakologi og Toksikologi (NSFT) Foredrag holdt av masterstudent ved Farmasøytisk institutt, Nuriddin Abdukadir, på Norsk Selskap for Farmakologi og Toksikologi sitt vintermøte 29. januar Vintermøtet ble arrangert på Radisson Blu Resort Beitostølen, Beitostølen, Norge. 63
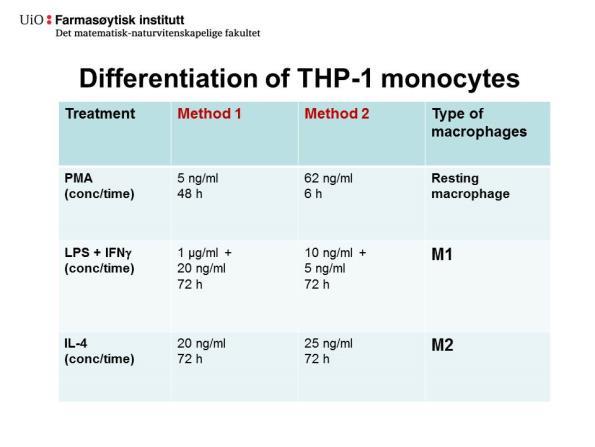
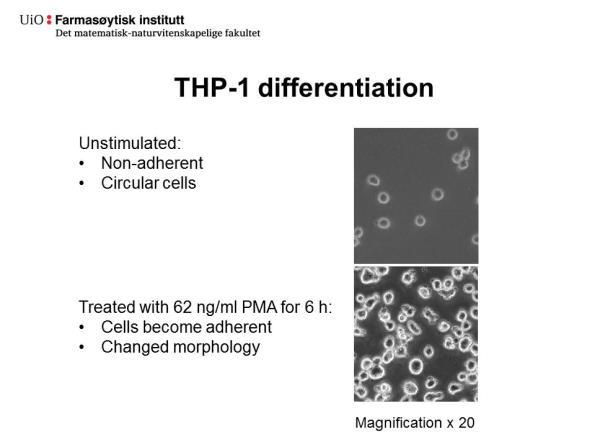
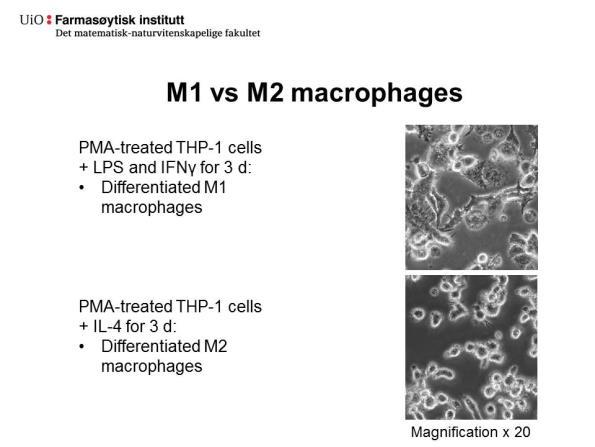
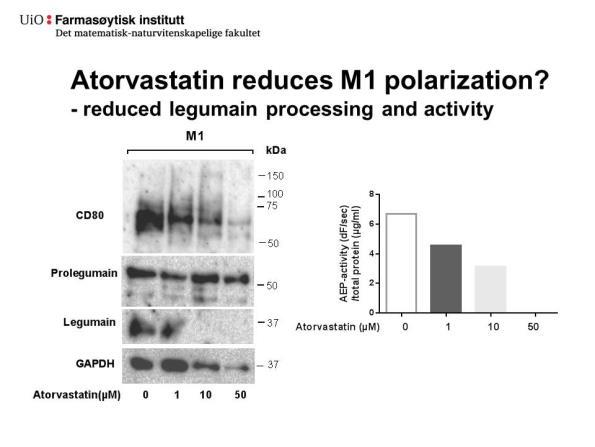
77 64
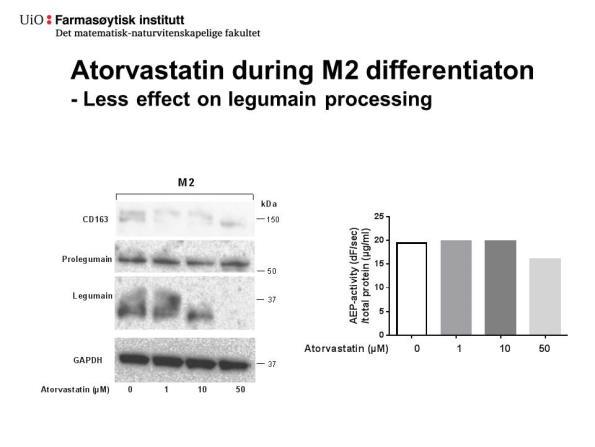
78 65
, Italia.")
79 Vedlegg C: Posterpresentasjon-Gordon Research Seminar Poster presentert av stipendiat Ngoc Nguyen Lunde, Seksjon for farmasøytisk biovitenskap, Farmasøytisk institutt, Universitetet i Oslo. Posteren ble presentert på The Gordon Reasearch Seminar on Proteolytic Enzymes and Their Inhibitors den juni, 2016, i Renaissance Tuscany II Ciocco, Lucca (Barga), Italia. 66
Hypoglykemi reduserer legumainaktivitet i makrofager
 Hypoglykemi reduserer legumainaktivitet i makrofager Økt legumain i plasma fra pasienter med karotisplakk Inass Elyouncha Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk
Hypoglykemi reduserer legumainaktivitet i makrofager Økt legumain i plasma fra pasienter med karotisplakk Inass Elyouncha Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk
Legumain i samspillet mellom tykktarmkreftceller og makrofager
 Legumain i samspillet mellom tykktarmkreftceller og makrofager Kjersti Bergh Ånonsen Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Legumain i samspillet mellom tykktarmkreftceller og makrofager Kjersti Bergh Ånonsen Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Etablering av metode for måling av inhibitorisk aktivitet mot cysteinproteaser og studier av cystatiner i makrofager og kreftceller
 Masteroppgave for graden Master i Farmasi Etablering av metode for måling av inhibitorisk aktivitet mot cysteinproteaser og studier av cystatiner i makrofager og kreftceller Hilde Rakvaag Mai 2007 Institutt
Masteroppgave for graden Master i Farmasi Etablering av metode for måling av inhibitorisk aktivitet mot cysteinproteaser og studier av cystatiner i makrofager og kreftceller Hilde Rakvaag Mai 2007 Institutt
Betydningen av legumain og cystatin E/M for aktivering av caspase-3
 Betydningen av legumain og cystatin E/M for aktivering av caspase-3 Masteroppgave av Dlvin Salem Hame Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematiske-naturvitenskaplige
Betydningen av legumain og cystatin E/M for aktivering av caspase-3 Masteroppgave av Dlvin Salem Hame Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematiske-naturvitenskaplige
Effekt av statiner på de lysosomale cysteinproteasene legumain, cathepsin B og L i monocytter/makrofager
 Effekt av statiner på de lysosomale cysteinproteasene legumain, cathepsin B og L i monocytter/makrofager Eyassu Tewolde Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk
Effekt av statiner på de lysosomale cysteinproteasene legumain, cathepsin B og L i monocytter/makrofager Eyassu Tewolde Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk
Glykosylering er nødvendig for internalisering og prosessering av prolegumain
 Glykosylering er nødvendig for internalisering og prosessering av prolegumain Kan modent legumain tas opp i levende humane celler? Kristina Broe Bodin Larsen Mastergradsoppgave i farmasi Seksjon for farmasøytisk
Glykosylering er nødvendig for internalisering og prosessering av prolegumain Kan modent legumain tas opp i levende humane celler? Kristina Broe Bodin Larsen Mastergradsoppgave i farmasi Seksjon for farmasøytisk
Cysteinproteasene legumain og cathepsin B i monocytter/makrofager og i aterosklerotiske plakk
 Cysteinproteasene legumain og cathepsin B i monocytter/makrofager og i aterosklerotiske plakk Sakina Ben Omar Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt
Cysteinproteasene legumain og cathepsin B i monocytter/makrofager og i aterosklerotiske plakk Sakina Ben Omar Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt
Laboratorieprotokoll for manuell rensing av DNA fra 0,5 ml prøve
 Laboratorieprotokoll for manuell rensing av DNA fra 0,5 ml prøve For rensing av genomisk DNA fra innsamlingssett i seriene Oragene og ORAcollect. Du finner flere språk og protokoller på vårt nettsted,
Laboratorieprotokoll for manuell rensing av DNA fra 0,5 ml prøve For rensing av genomisk DNA fra innsamlingssett i seriene Oragene og ORAcollect. Du finner flere språk og protokoller på vårt nettsted,
NITO Bioingeniørfaglig Institutt kurs i Immunologi The Edge, Tromsø, 11. februar 2015. Generell Immunologi
 NITO Bioingeniørfaglig Institutt kurs i Immunologi The Edge, Tromsø, 11. februar 2015 Generell Immunologi Tor B Stuge Immunologisk forskningsgruppe, IMB Universitetet i Tromsø Innhold: 1. Immunsystemets
NITO Bioingeniørfaglig Institutt kurs i Immunologi The Edge, Tromsø, 11. februar 2015 Generell Immunologi Tor B Stuge Immunologisk forskningsgruppe, IMB Universitetet i Tromsø Innhold: 1. Immunsystemets
Nytte av prokalsitonin og nøytrofil CD64 som markør for postoperativ infeksjon
 Nytte av prokalsitonin og nøytrofil CD64 som markør for postoperativ infeksjon Kurs i hematologi 18. 19. mai 2015 KS møtesenter, Oslo Kristin Husby, Spesialbioingeniør Prosjektet er utført ved Tverrfaglig
Nytte av prokalsitonin og nøytrofil CD64 som markør for postoperativ infeksjon Kurs i hematologi 18. 19. mai 2015 KS møtesenter, Oslo Kristin Husby, Spesialbioingeniør Prosjektet er utført ved Tverrfaglig
Statiner hemmer aktivitet av den lysosomale cysteinproteasen legumain Spiller legumain en rolle i apoptose?
 Statiner hemmer aktivitet av den lysosomale cysteinproteasen legumain Spiller legumain en rolle i apoptose? Islam Mohammed Yaseen Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk
Statiner hemmer aktivitet av den lysosomale cysteinproteasen legumain Spiller legumain en rolle i apoptose? Islam Mohammed Yaseen Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk
Atorvastatin hemmer aktiviteten av cysteinproteaser og stimulerer sekresjonen av MMP-9
 Atorvastatin hemmer aktiviteten av cysteinproteaser og stimulerer sekresjonen av MMP-9 Anny Thi Tran Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematiske-naturvitenskapelige
Atorvastatin hemmer aktiviteten av cysteinproteaser og stimulerer sekresjonen av MMP-9 Anny Thi Tran Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematiske-naturvitenskapelige
Hvordan holde styr på proteasene?? Harald; 31.01.08
 Hvordan holde styr på proteasene?? Harald; 31.01.08 Protease? Proteinase? Spalter proteiner Peptidase? Spalter peptider? Spalter peptidbindinger Exo-peptidaser Endo-peptidaser Endo Ekso- Proteaser er enzymer!
Hvordan holde styr på proteasene?? Harald; 31.01.08 Protease? Proteinase? Spalter proteiner Peptidase? Spalter peptider? Spalter peptidbindinger Exo-peptidaser Endo-peptidaser Endo Ekso- Proteaser er enzymer!
Forelesninger i BI Cellebiologi. Protein struktur og funksjon - Kap. 3
 Forelesninger i BI 212 - Cellebiologi Protein struktur og funksjon - Kap. 3 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio.ntnu.no Tlf. 73 59
Forelesninger i BI 212 - Cellebiologi Protein struktur og funksjon - Kap. 3 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio.ntnu.no Tlf. 73 59
Cellulære effekter av et kolkisin-prodrug som spaltes av legumain
 Cellulære effekter av et kolkisin-prodrug som spaltes av legumain Tina Elvestrand Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap, Farmasøytisk institutt Det matematisk-naturvitenskapelige
Cellulære effekter av et kolkisin-prodrug som spaltes av legumain Tina Elvestrand Mastergradsoppgave i farmasi Avdeling for farmasøytisk biovitenskap, Farmasøytisk institutt Det matematisk-naturvitenskapelige
Selv. Trygg Ignorere. Farlig. Fremmed. Immunsystemet: Generelt, og om allergi spesielt. Immunsystemets utfordring. Medfødte og ervervete immunsystemet
 U N I V E R S I T E T E T I B E R G E N Immunsystemet: Generelt, og om allergi spesielt Silke Appel Broegelmanns Forskningslaboratorium Klinisk Institutt 2 Universitetet i Bergen silke.appel@ Immunsystemets
U N I V E R S I T E T E T I B E R G E N Immunsystemet: Generelt, og om allergi spesielt Silke Appel Broegelmanns Forskningslaboratorium Klinisk Institutt 2 Universitetet i Bergen silke.appel@ Immunsystemets
Forelesninger i BI Cellebiologi Proteinrensing - Væskekromatografi. Figure 3-43 b
 Proteinrensing - Væskekromatografi Figure 3-43 b Proteinrensing - Væskekromatografi Ved affinitets-kromatografi brukes en søyle med kuler som er dekket med ligander (f.eks. et enzym-substrat eller et annet
Proteinrensing - Væskekromatografi Figure 3-43 b Proteinrensing - Væskekromatografi Ved affinitets-kromatografi brukes en søyle med kuler som er dekket med ligander (f.eks. et enzym-substrat eller et annet
1 Bakgrunn Metode og gjennomføring Belegg Biofilmdannelse Resultater Biofilmdannelse Diskusjon...
 Innholdsfortegnelse Bakgrunn... 3 2 Metode og gjennomføring... 4 2. Belegg... 4 2.2 Biofilmdannelse... 4 3 Resultater... 5 3. Biofilmdannelse... 5 4 Diskusjon... 6 5 Foreløpige konklusjoner... 7 60205-2
Innholdsfortegnelse Bakgrunn... 3 2 Metode og gjennomføring... 4 2. Belegg... 4 2.2 Biofilmdannelse... 4 3 Resultater... 5 3. Biofilmdannelse... 5 4 Diskusjon... 6 5 Foreløpige konklusjoner... 7 60205-2
Forløp av ikke-adaptiv og adaptiv immunrespons. Mononukleære celler, metylfiolett farging
 Forløp av ikke-adaptiv og adaptiv immunrespons Mononukleære celler, metylfiolett farging 1 Nøytrofile granulocytter Gjenkjennelsesprinsipper medfødt vs. adaptiv immunitet Toll Like Receptors Mikroorganismer
Forløp av ikke-adaptiv og adaptiv immunrespons Mononukleære celler, metylfiolett farging 1 Nøytrofile granulocytter Gjenkjennelsesprinsipper medfødt vs. adaptiv immunitet Toll Like Receptors Mikroorganismer
Oppgave 16.2 fortsetter
 Oppgave 16.2 fortsetter Vask av presipiteringsgeler for å fjerne ikke-presipitert protein (bakgrunn): Vask 3 x 15 min + 10 min press i fysiologisk saltvann /evt. over natt + 10 min. press. eretter 1 x
Oppgave 16.2 fortsetter Vask av presipiteringsgeler for å fjerne ikke-presipitert protein (bakgrunn): Vask 3 x 15 min + 10 min press i fysiologisk saltvann /evt. over natt + 10 min. press. eretter 1 x
N-bundet glykosylering: Betydning for sekresjon og autoaktivering av legumain i humane celler
 N-bundet glykosylering: Betydning for sekresjon og autoaktivering av legumain i humane celler Roya Kasem Mastergradsoppgave i farmasi Seksjon for farmasøytisk biovitenskap, Farmasøytisk institutt Det matematisk-naturvitenskapelige
N-bundet glykosylering: Betydning for sekresjon og autoaktivering av legumain i humane celler Roya Kasem Mastergradsoppgave i farmasi Seksjon for farmasøytisk biovitenskap, Farmasøytisk institutt Det matematisk-naturvitenskapelige
BIOS 2 Biologi
 . Figurer kapittel 2: Energi Figur s. 48 Solenergi Økosystem CO 2 + 2 O Fotosyntese i kloroplaster Organiske molekyler + O 2 Celleånding i mitokondrier Energi til arbeid Varme rodusentene i økosystemet
. Figurer kapittel 2: Energi Figur s. 48 Solenergi Økosystem CO 2 + 2 O Fotosyntese i kloroplaster Organiske molekyler + O 2 Celleånding i mitokondrier Energi til arbeid Varme rodusentene i økosystemet
4. Antigenpresentasjon til T celler. MHC molekyler.
 Immunologi 3. semester, V2015 4. Antigenpresentasjon til T celler. MHC molekyler. Karl Schenck Institutt for oral biologi TcR og MHC molekyler Noen hovedgrupper T celler som forlater thymus T celler med
Immunologi 3. semester, V2015 4. Antigenpresentasjon til T celler. MHC molekyler. Karl Schenck Institutt for oral biologi TcR og MHC molekyler Noen hovedgrupper T celler som forlater thymus T celler med
1. Medfødt og ervervet immunitet. Karl Schenck, V2015
 1. Medfødt og ervervet immunitet Karl Schenck, V2015 Medfødt og ervervet immunforsvar Antimicrobial peptides «Alltid beredt!» Relativt uspesifikt Må aktiveres Spesifikt Komponenter av medfødt immunitet
1. Medfødt og ervervet immunitet Karl Schenck, V2015 Medfødt og ervervet immunforsvar Antimicrobial peptides «Alltid beredt!» Relativt uspesifikt Må aktiveres Spesifikt Komponenter av medfødt immunitet
Oncogenic Mutations Affecting Cell Proliferation
 Oncogenic Mutations Affecting Cell Proliferation Fra RTK til Nucleus (Boka s.1070-74) Normalt kreves et vekst stimulerende signal ( growth factor eks. PDGF, EGF, NGF) for at celler skal gå inn i celledeling,
Oncogenic Mutations Affecting Cell Proliferation Fra RTK til Nucleus (Boka s.1070-74) Normalt kreves et vekst stimulerende signal ( growth factor eks. PDGF, EGF, NGF) for at celler skal gå inn i celledeling,
Antitrombin i laboratoriet
 Antitrombin i laboratoriet Carola Henriksson, Seksjonsoverlege, hematologiseksjonen, avdeling for medisinsk biokjemi, Oslo Universitetssykehus, Ullevål Hurdalsjøen, 8. juni Oslo Universitetssykehus (OUS)
Antitrombin i laboratoriet Carola Henriksson, Seksjonsoverlege, hematologiseksjonen, avdeling for medisinsk biokjemi, Oslo Universitetssykehus, Ullevål Hurdalsjøen, 8. juni Oslo Universitetssykehus (OUS)
5. T cellers effektorfunksjoner
 Immunologi 3. semester, V2015 5. T cellers effektorfunksjoner Karl Schenck Institutt for oral biologi Noen typer effektor T celler 3 3 IFNg IL-4 IL-17 IL-10 TGFb TGFb IL-4 Secretory IgA Enhance mast cell
Immunologi 3. semester, V2015 5. T cellers effektorfunksjoner Karl Schenck Institutt for oral biologi Noen typer effektor T celler 3 3 IFNg IL-4 IL-17 IL-10 TGFb TGFb IL-4 Secretory IgA Enhance mast cell
Membran-proteiner (Del 3.4)
") Membran-proteiner (Del 3.4) Poriner adskiller seg dramatisk fra andre integral proteiner. Finnes bl.a. i ytre membranen hos E.coli (se Figure 1-7). Poriner er med å beskytte bakterien mot toksiske forbindelser
Membran-proteiner (Del 3.4) Poriner adskiller seg dramatisk fra andre integral proteiner. Finnes bl.a. i ytre membranen hos E.coli (se Figure 1-7). Poriner er med å beskytte bakterien mot toksiske forbindelser
Enzymes make the world go around. Enzymer i dagliglivet
 Enzymes make the world go around Enzymer i dagliglivet Innledning Enzymer er i de fleste tilfellene proteiner som øker reaksjonshastigheten til biologiske prosesser. Derfor blir enzymer ofte kalt biologiske
Enzymes make the world go around Enzymer i dagliglivet Innledning Enzymer er i de fleste tilfellene proteiner som øker reaksjonshastigheten til biologiske prosesser. Derfor blir enzymer ofte kalt biologiske
T celle aktivering og HLA Trond S. S. Halstensen
 T celle aktivering og HLA Trond S. S. Halstensen T-celler og Thymus T cellens identifisering av antigener Human Leukocyt Antigen (HLA) restriksjon, CD4 og CD8 Antigen prosessering: cytosol- og endocytisk
T celle aktivering og HLA Trond S. S. Halstensen T-celler og Thymus T cellens identifisering av antigener Human Leukocyt Antigen (HLA) restriksjon, CD4 og CD8 Antigen prosessering: cytosol- og endocytisk
Trombotisk Trombocytopenisk Purpura og ADAMTS13
 Trombotisk Trombocytopenisk Purpura og ADAMTS13 Bergen 14. september 2017 Bioingeniør Siri Lie Størkersen Seksjon spesiell biokjemi Avdeling for medisinsk biokjemi St Olavs hospital Analysering av ADAMTS13-aktivitet
Trombotisk Trombocytopenisk Purpura og ADAMTS13 Bergen 14. september 2017 Bioingeniør Siri Lie Størkersen Seksjon spesiell biokjemi Avdeling for medisinsk biokjemi St Olavs hospital Analysering av ADAMTS13-aktivitet
IMMUNOLOGISKE METODER Linn Silje Hansen. Bioingeniør ved avdeling for immunologi og transfusjonsmedisin, seksjon immunologi og cytometri
 Bilde fra: https://bioprocessintl.com/sponsored-content/keeping-host-cell-protein-elisas-covered/ (26.11.18) IMMUNOLOGISKE METODER Linn Silje Hansen Bioingeniør ved avdeling for immunologi og transfusjonsmedisin,
Bilde fra: https://bioprocessintl.com/sponsored-content/keeping-host-cell-protein-elisas-covered/ (26.11.18) IMMUNOLOGISKE METODER Linn Silje Hansen Bioingeniør ved avdeling for immunologi og transfusjonsmedisin,
Cellesignalisering II: Reseptor tyrosin kinaser, cytosoliske kinaser
 Cellesignalisering II: Reseptor tyrosin kinaser, cytosoliske kinaser! Introduksjon! Definisjon og klassifisering! Kinasefamilier: Receptor/cytosol! Receptor Tyrosin kinase-mediert signalisering! MAP kinase
Cellesignalisering II: Reseptor tyrosin kinaser, cytosoliske kinaser! Introduksjon! Definisjon og klassifisering! Kinasefamilier: Receptor/cytosol! Receptor Tyrosin kinase-mediert signalisering! MAP kinase
Er cysteinproteaser involvert i statinindusert myotoksisitet?
 Er cysteinproteaser involvert i statinindusert myotoksisitet? Anette Larsen Voreland Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Er cysteinproteaser involvert i statinindusert myotoksisitet? Anette Larsen Voreland Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Grunnleggende immunologi
 Grunnleggende immunologi Anne Storset anne.storset@nmbu.no Institutt for Mattrygghet og infeksjonsbiologi Norges miljø- og biovitenskapelige universitet 1 Sykdom Ervervet immunitet Medfødt immunitet Fysiske
Grunnleggende immunologi Anne Storset anne.storset@nmbu.no Institutt for Mattrygghet og infeksjonsbiologi Norges miljø- og biovitenskapelige universitet 1 Sykdom Ervervet immunitet Medfødt immunitet Fysiske
Nr. 9/60 EØS-tillegget til Den europeiske unions tidende KOMMISJONSVEDTAK. av 21. februar 2002
 Nr. 9/60 EØS-tillegget til Den europeiske unions tidende 24.2.2005 KOMMISJONSVEDTAK 2005/EØS/9/09 av 21. februar 2002 om endring av vedlegg D til rådsdirektiv 90/426/EØF med hensyn til diagnostiske prøver
Nr. 9/60 EØS-tillegget til Den europeiske unions tidende 24.2.2005 KOMMISJONSVEDTAK 2005/EØS/9/09 av 21. februar 2002 om endring av vedlegg D til rådsdirektiv 90/426/EØF med hensyn til diagnostiske prøver
Inflammasjonens betydning for endotel dysfunksjon og atherosklerose
 Inflammasjonens betydning for endotel dysfunksjon og atherosklerose Ingebjørg Seljeflot Senter for klinisk hjerteforskning Kardiologisk avdeling Oslo universitetssykehus Ullevål http:/ous-research.no/clinicalheartresearch/
Inflammasjonens betydning for endotel dysfunksjon og atherosklerose Ingebjørg Seljeflot Senter for klinisk hjerteforskning Kardiologisk avdeling Oslo universitetssykehus Ullevål http:/ous-research.no/clinicalheartresearch/
NYHETSAVIS NR. 2/2000 Oktober 2000
 NYHETSAVIS NR. 2/2000 Oktober 2000 INNHOLD: Analysenytt Endret metode for påvisning av thyreoidea autoantistoffer (TRAS, anti-tpo og anti-tg) Endret referanseområde for hcg hos gravide Bestemmelse av jod
NYHETSAVIS NR. 2/2000 Oktober 2000 INNHOLD: Analysenytt Endret metode for påvisning av thyreoidea autoantistoffer (TRAS, anti-tpo og anti-tg) Endret referanseområde for hcg hos gravide Bestemmelse av jod
Immunstimulanter for potensiering av torskens naturlige immunsystem
 Store programmer HAVBRUK - En næring i vekst Faktaark www.forskningsradet.no/havbruk Immunstimulanter for potensiering av torskens naturlige immunsystem Prosjekt: Immunostimulation of Atlantic cod (Gadus
Store programmer HAVBRUK - En næring i vekst Faktaark www.forskningsradet.no/havbruk Immunstimulanter for potensiering av torskens naturlige immunsystem Prosjekt: Immunostimulation of Atlantic cod (Gadus
Bruksanvisning IVD Matrix HCCA-portioned
 8290200 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til matriksassistert laserdesorpsjons-ionisasjon flytid for massespektrometri (MALDI-TOF-MS). CARE- produkter er utformet for å støtte
8290200 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til matriksassistert laserdesorpsjons-ionisasjon flytid for massespektrometri (MALDI-TOF-MS). CARE- produkter er utformet for å støtte
Colostrum FAQ. Hyppig stilte spørsmål om LRs Colostrum-produkter
 Colostrum FAQ Hyppig stilte spørsmål om LRs Colostrum-produkter Innholdsfortegnelse I. Hva er colostrum? S 3 II. Hvilket dyr kommer colostrum fra? S 3 III. Hva skjer med kalvene? S 3 IV. Hvorfor er colostrum
Colostrum FAQ Hyppig stilte spørsmål om LRs Colostrum-produkter Innholdsfortegnelse I. Hva er colostrum? S 3 II. Hvilket dyr kommer colostrum fra? S 3 III. Hva skjer med kalvene? S 3 IV. Hvorfor er colostrum
Hensikten med forsøket er å isolere eget DNA fra kinnceller, se hvordan det ser ut og hva det kan brukes til videre.
 DNA HALSKJEDE Hensikt Hensikten med forsøket er å isolere eget DNA fra kinnceller, se hvordan det ser ut og hva det kan brukes til videre. Bakgrunn Det humane genomet består av omtrent 2.9 milliarder basepar.
DNA HALSKJEDE Hensikt Hensikten med forsøket er å isolere eget DNA fra kinnceller, se hvordan det ser ut og hva det kan brukes til videre. Bakgrunn Det humane genomet består av omtrent 2.9 milliarder basepar.
Kreftforskning.no/myklebost. Eva Wessel Pedersen. Cancer Stem Cell Innovation Centre
 Stam Celler og Kreft Eva Wessel Pedersen Avdeling for Tumorbiologi,, Radium Hospitalet Cancer Stem Cell Innovation Centre Oversikt Stamceller generelt Hvorfor vi forsker på stamceller Kreft-stamceller
Stam Celler og Kreft Eva Wessel Pedersen Avdeling for Tumorbiologi,, Radium Hospitalet Cancer Stem Cell Innovation Centre Oversikt Stamceller generelt Hvorfor vi forsker på stamceller Kreft-stamceller
Oppgave: MED1100-3_OPPGAVE2_V18_ORD
 Side 15 av 46 Oppgave: MED1100-3_OPPGAVE2_V18_ORD Del 1: Hvilke av de følgende celler uttrykker normalt (i hvilende tilstand) HLA klasse II molekyler hos mennesket? Angi de tre riktigste svarene. Fibroblaster
Side 15 av 46 Oppgave: MED1100-3_OPPGAVE2_V18_ORD Del 1: Hvilke av de følgende celler uttrykker normalt (i hvilende tilstand) HLA klasse II molekyler hos mennesket? Angi de tre riktigste svarene. Fibroblaster
Tuberkulose.
 Tuberkulose behov for mer effektiv behandling og nye vaksiner behov for å forstå vert-mikrobe forholdet bedre Persontilpasset behandling: immunterapi + antibiotika www.ntnu.edu/cemir Forskning på tuberkulose
Tuberkulose behov for mer effektiv behandling og nye vaksiner behov for å forstå vert-mikrobe forholdet bedre Persontilpasset behandling: immunterapi + antibiotika www.ntnu.edu/cemir Forskning på tuberkulose
TNF/BiO og dermatologiske utfordringer. Øystein Sandanger, MD, PhD Seksjon for hudsykdommer, OUS Institutt for indremedisinsk forskning, OUS
 TNF/BiO og dermatologiske utfordringer Øystein Sandanger, MD, PhD Seksjon for hudsykdommer, OUS Institutt for indremedisinsk forskning, OUS Viktige dermatologiske lidelser som behandles med monoklonale
TNF/BiO og dermatologiske utfordringer Øystein Sandanger, MD, PhD Seksjon for hudsykdommer, OUS Institutt for indremedisinsk forskning, OUS Viktige dermatologiske lidelser som behandles med monoklonale
Nr. 46/108 EØS-tillegget til De Europeiske Fellesskaps Tidende KOMMISJONSDIREKTIV 1999/76/EF. av 23. juli 1999
 Nr. 46/108 EØS-tillegget til De Europeiske Fellesskaps Tidende KOMMISJONEN FOR DE EUROPEISKE FELLESSKAP HAR under henvisning til traktaten om opprettelse av Det europeiske fellesskap, under henvisning
Nr. 46/108 EØS-tillegget til De Europeiske Fellesskaps Tidende KOMMISJONEN FOR DE EUROPEISKE FELLESSKAP HAR under henvisning til traktaten om opprettelse av Det europeiske fellesskap, under henvisning
Tissue_LC_200_V7_DSP og Tissue_HC_200_V7_DSP (brukervalidert for QIAsymphony DSP DNA Mini-sett)
") August 2015 QIAsymphony SP-protokollark Tissue_LC_200_V7_DSP og Tissue_HC_200_V7_DSP (brukervalidert for QIAsymphony DSP DNA Mini-sett) Dette dokumentet er Tissue_LC_200_V7_DSP og Tissue_HC_200_V7_DSP
August 2015 QIAsymphony SP-protokollark Tissue_LC_200_V7_DSP og Tissue_HC_200_V7_DSP (brukervalidert for QIAsymphony DSP DNA Mini-sett) Dette dokumentet er Tissue_LC_200_V7_DSP og Tissue_HC_200_V7_DSP
Cellebiologiske prosesser som respons på virusinfeksjon
 Cellebiologiske prosesser som respons på virusinfeksjon PBM 336 2005 Siri Mjaaland Infeksjoner - immunresponser 1 Figure 2-49 Interferoner Uspesifikk immunitet viral infeksjon stimulerer direkte produksjon
Cellebiologiske prosesser som respons på virusinfeksjon PBM 336 2005 Siri Mjaaland Infeksjoner - immunresponser 1 Figure 2-49 Interferoner Uspesifikk immunitet viral infeksjon stimulerer direkte produksjon
BI 212- Protein Sorting - Kap. 17 Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.)
") Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.) Veiene for opptak fra cytosol av kloroplast-proteiner Opptak av proteiner fra cytosol til kloroplaster ligner mye på mitokondrie-importen
Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.) Veiene for opptak fra cytosol av kloroplast-proteiner Opptak av proteiner fra cytosol til kloroplaster ligner mye på mitokondrie-importen
Obligatorisk oppgave 2 MBV1030 Høst 2005
 Obligatorisk oppgave 2 MBV1030 Høst 2005 Levert av (navn): Første del: Flervalgsspørsmål. Angi det svaralternativet (ett) du mener er korrekt. I-1: Ved anaerob glykolyse dannes det laktat. Dersom glukosen
Obligatorisk oppgave 2 MBV1030 Høst 2005 Levert av (navn): Første del: Flervalgsspørsmål. Angi det svaralternativet (ett) du mener er korrekt. I-1: Ved anaerob glykolyse dannes det laktat. Dersom glukosen
DNA isolering Elektroforese av DNA og protein
 DNA isolering Elektroforese av DNA og protein Et teoritisk labkurs Medisin IA v/ Øyvind Halaas 1 Molekylær biologi er i hovedsak studiet av informasjonsbærerne DNA og proteiner De mest brukte metodene
DNA isolering Elektroforese av DNA og protein Et teoritisk labkurs Medisin IA v/ Øyvind Halaas 1 Molekylær biologi er i hovedsak studiet av informasjonsbærerne DNA og proteiner De mest brukte metodene
QIAsymphony SP protokollark
 QIAsymphony SP protokollark DNA_Blood_400_V6_DSP-protokoll Generell informasjon Til in vitro-diagnostisk bruk. Denne protokollen brukes til rensing av totalt genomisk og mitokondrielt DNA fra friskt eller
QIAsymphony SP protokollark DNA_Blood_400_V6_DSP-protokoll Generell informasjon Til in vitro-diagnostisk bruk. Denne protokollen brukes til rensing av totalt genomisk og mitokondrielt DNA fra friskt eller
Påvisning av anti dsdna men forskjellige metoder og teknikker. Innhold. Årsaksmekanismer. Metoder. Forekomst 13.02.2015
 Påvisning av anti dsdna men forskjellige metoder og teknikker Randi Karlsen Fagbioingeniør Seksjon for medisinsk immunologi Oslo Universitetssykehus Ullevål sykehus Februar 2015 Innhold Årsaksmekanismer?
Påvisning av anti dsdna men forskjellige metoder og teknikker Randi Karlsen Fagbioingeniør Seksjon for medisinsk immunologi Oslo Universitetssykehus Ullevål sykehus Februar 2015 Innhold Årsaksmekanismer?
Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu
,Botanisk institutt,ntnu") Forelesninger i BI 212 - Cellebiologi Biomembraner og subcellular organisering av eukaryote celler - Kap. 5 - vår 2002 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-
Forelesninger i BI 212 - Cellebiologi Biomembraner og subcellular organisering av eukaryote celler - Kap. 5 - vår 2002 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-
Institutt for biologi Faglig kontaktperson(er) under eksamen: Diem Hong Tran mobil tlf. nr Sensurdato: 3 uker fra eksamen + 10 dager
 under eksamen: Diem Hong Tran mobil tlf. nr Sensurdato: 3 uker fra eksamen + 10 dager") Side 1 av 7 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson(er) under eksamen: Diem Hong Tran mobil tlf. nr. 906 30 861
Side 1 av 7 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson(er) under eksamen: Diem Hong Tran mobil tlf. nr. 906 30 861
Forelesninger i BI Cellebiologi. Denaturering og renaturering. Figure 3-13
 Figure 3.9 Denaturering og renaturering Figure 3-13 Denaturering og renaturering Figure 3-14 Viser tre trinn i refolding av et protein som har vært denaturert. Molten globule -formen er en intermediær
Figure 3.9 Denaturering og renaturering Figure 3-13 Denaturering og renaturering Figure 3-14 Viser tre trinn i refolding av et protein som har vært denaturert. Molten globule -formen er en intermediær
Oppgave: MED1100-3_OPPGAVE2_H16_KONT
 Side 10 av 35 Oppgave: MED1100-3_OPPGAVE2_H16_KONT Del 1: Ola har en arvelig betinget kombinert immundefekt med mangel på både T-celler og B-celler. Ola får derfor gjentatte Hvorfor er Ola beskyttet mot
Side 10 av 35 Oppgave: MED1100-3_OPPGAVE2_H16_KONT Del 1: Ola har en arvelig betinget kombinert immundefekt med mangel på både T-celler og B-celler. Ola får derfor gjentatte Hvorfor er Ola beskyttet mot
Idrett og energiomsetning
 1 Medisin stadium IA, Tonje S. Steigedal 2 ATP er den eneste forbindelsen som kan drive kontraksjon av musklene. ATPnivået i muskelcellene er imidlertid begrenset, og må etterfylles kontinuerlig. Ved ulike
1 Medisin stadium IA, Tonje S. Steigedal 2 ATP er den eneste forbindelsen som kan drive kontraksjon av musklene. ATPnivået i muskelcellene er imidlertid begrenset, og må etterfylles kontinuerlig. Ved ulike
NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK. EKSAMEN I FAG CELLEBIOLOGI 1 august 1997 Tid: kl
 NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK Faglig kontakt under eksamen: Navn: Professor Tore Lindmo Tlf.:93432 EKSAMEN I FAG 74618 CELLEBIOLOGI 1 august 1997 Tid: kl
NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK Faglig kontakt under eksamen: Navn: Professor Tore Lindmo Tlf.:93432 EKSAMEN I FAG 74618 CELLEBIOLOGI 1 august 1997 Tid: kl
Identifisering av PAX- og MMP-uttrykk i tre prostatakreft-cellelinjer
 HOVEDOPPGAVE I FARMAKOLOGI FOR MASTERGRADEN I FARMASI Identifisering av PAX- og MMP-uttrykk i tre prostatakreft-cellelinjer Av Elisabeth Weibust 2009 Utført ved Avdeling for farmakologi, Institutt for
HOVEDOPPGAVE I FARMAKOLOGI FOR MASTERGRADEN I FARMASI Identifisering av PAX- og MMP-uttrykk i tre prostatakreft-cellelinjer Av Elisabeth Weibust 2009 Utført ved Avdeling for farmakologi, Institutt for
FLERVALGSOPPGAVER ANALYSE
 FLERVALGSOPPGAVER ANALYSE Hjelpemidler: Periodesystem (og kalkulator der det er angitt) Hvert spørsmål har et riktig svaralternativ. Når ikke noe annet er oppgitt kan du anta STP (standard trykk og temperatur).
FLERVALGSOPPGAVER ANALYSE Hjelpemidler: Periodesystem (og kalkulator der det er angitt) Hvert spørsmål har et riktig svaralternativ. Når ikke noe annet er oppgitt kan du anta STP (standard trykk og temperatur).
Fastere filet - Aktivitet 2
 Fastere filet - Aktivitet 2 MRI, protease og protein analyser Iciar Martinez 1,2, Rasa Slizyté 1, Pål Anders Wang 2,Stine W. Dahle 1, Michiaki Yamashita 3, Ragnar Olsen 2, Emil Veliuylin 1 og Ulf Erikson
Fastere filet - Aktivitet 2 MRI, protease og protein analyser Iciar Martinez 1,2, Rasa Slizyté 1, Pål Anders Wang 2,Stine W. Dahle 1, Michiaki Yamashita 3, Ragnar Olsen 2, Emil Veliuylin 1 og Ulf Erikson
Bruksanvisning IVD Matrix HCCA-portioned
 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
Preanalyse. Kurs i Molekylærpatologi Oslo, juni 2017
 Preanalyse Kurs i Molekylærpatologi Oslo, 07.-08. juni 2017 Heidemarie Svendsen Bioingeniør Enhet for molekylærpatologi Avdeling for Patologi Oslo Universitetssykehus Innhold Prøvemateriale Preparering
Preanalyse Kurs i Molekylærpatologi Oslo, 07.-08. juni 2017 Heidemarie Svendsen Bioingeniør Enhet for molekylærpatologi Avdeling for Patologi Oslo Universitetssykehus Innhold Prøvemateriale Preparering
Klinisk molekylærmedisin (5): Eksempler på funksjonelle analyser
: Eksempler på funksjonelle analyser") Pediatrisk Endokrinologi 2003;17: 64-69 Klinisk molekylærmedisin (5): Eksempler på funksjonelle analyser Pål Rasmus Njølstad 1,2,3, Lise Bjørkhaug 1 1 Seksjon for pediatri, Institutt for klinisk medisin
Pediatrisk Endokrinologi 2003;17: 64-69 Klinisk molekylærmedisin (5): Eksempler på funksjonelle analyser Pål Rasmus Njølstad 1,2,3, Lise Bjørkhaug 1 1 Seksjon for pediatri, Institutt for klinisk medisin
EKSAMENSOPPGAVE I BI2014 MOLEKYLÆRBIOLOGI
 Norges teknisk-naturvitenskapelige universitet Institutt for biologi EKSAMENSOPPGAVE I BI014 MOLEKYLÆRBIOLOGI Faglig kontakt under eksamen: Ralph Kissen Tlf.: 41344134 (mobil) - Eksamensdato: 11. desember
Norges teknisk-naturvitenskapelige universitet Institutt for biologi EKSAMENSOPPGAVE I BI014 MOLEKYLÆRBIOLOGI Faglig kontakt under eksamen: Ralph Kissen Tlf.: 41344134 (mobil) - Eksamensdato: 11. desember
Molekylære metoder i medisinsk mikrobiologi.
 - 1 - Molekylære metoder i medisinsk mikrobiologi. Laboratorieøvelser onsdag 18.mars 2014. Øvelser: 5. Gelelektroforese på PCR produkt fra meca og nuc PCR fra dag 1 6. Mycobakterium genus FRET realtime
- 1 - Molekylære metoder i medisinsk mikrobiologi. Laboratorieøvelser onsdag 18.mars 2014. Øvelser: 5. Gelelektroforese på PCR produkt fra meca og nuc PCR fra dag 1 6. Mycobakterium genus FRET realtime
Turbidimetri og nefelometri. Olav Klingenberg Overlege, dr.med Avdeling for medisinsk biokjemi OUS-Rikshospitalet
 Turbidimetri og nefelometri Olav Klingenberg Overlege, dr.med Avdeling for medisinsk biokjemi OUS-Rikshospitalet Nefelometri Turbidimetri Partikler Hva slags partikler sprer lys? Molekyler Gassmolekyler
Turbidimetri og nefelometri Olav Klingenberg Overlege, dr.med Avdeling for medisinsk biokjemi OUS-Rikshospitalet Nefelometri Turbidimetri Partikler Hva slags partikler sprer lys? Molekyler Gassmolekyler
Generell immunologi og immunforsvaret på 45 minutt
 Generell immunologi og immunforsvaret på 45 minutt NITO BIOINGENIØRFAGLIG INSTITUTT, Immunologi og immunologiske metoder, etterutdanningskurs 27. 28.november 2018 Kirsti Hokland bioingeniør og universitetslektor
Generell immunologi og immunforsvaret på 45 minutt NITO BIOINGENIØRFAGLIG INSTITUTT, Immunologi og immunologiske metoder, etterutdanningskurs 27. 28.november 2018 Kirsti Hokland bioingeniør og universitetslektor
Amplifikasjonsteknikker - andre metoder
 Amplifikasjonsteknikker - andre metoder Svein Arne Nordbø TH-28973 17.03.15 Alternative amplifikasjonsmetoder Templat-amplifikasjons metoder Signal-amplifikasjonsmetoder Templat-amplifikasjons metoder
Amplifikasjonsteknikker - andre metoder Svein Arne Nordbø TH-28973 17.03.15 Alternative amplifikasjonsmetoder Templat-amplifikasjons metoder Signal-amplifikasjonsmetoder Templat-amplifikasjons metoder
Leucosep-rør LTK.615 PAKNINGSVEDLEGG. Brukes til in vitro-diagnostikk PI-LT.615-NO-V3
 Leucosep-rør LTK.615 PAKNINGSVEDLEGG Brukes til in vitro-diagnostikk PI-LT.615-NO-V3 Bruksanvisning Bruksområde Leucosep-rørene er beregnet til bruk ved oppsamling og separasjon av mononukleære celler
Leucosep-rør LTK.615 PAKNINGSVEDLEGG Brukes til in vitro-diagnostikk PI-LT.615-NO-V3 Bruksanvisning Bruksområde Leucosep-rørene er beregnet til bruk ved oppsamling og separasjon av mononukleære celler
Institutt for biologi Faglig kontaktperson(er) under eksamen: Diem Hong Tran, mobil tlf. nr
 under eksamen: Diem Hong Tran, mobil tlf. nr") Side 1 av 6 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson(er) under eksamen: Diem Hong Tran, mobil tlf. nr. 906 30 861
Side 1 av 6 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson(er) under eksamen: Diem Hong Tran, mobil tlf. nr. 906 30 861
QIAsymphony SP protokollark
 Februar 2017 QIAsymphony SP protokollark circdna_2000_dsp_v1 og circdna_4000_dsp_v1 Dette dokumentet er QIAsymphony circdna_2000_dsp_v1 og circdna_4000_dsp_v1- protokollark, versjon 1, R1 Sample to Insight
Februar 2017 QIAsymphony SP protokollark circdna_2000_dsp_v1 og circdna_4000_dsp_v1 Dette dokumentet er QIAsymphony circdna_2000_dsp_v1 og circdna_4000_dsp_v1- protokollark, versjon 1, R1 Sample to Insight
Dei lysosomale cysteinproteasane legumain og cathepsin B i apoptose
 Dei lysosomale cysteinproteasane legumain og cathepsin B i apoptose Eilen Tungland Mastergradsoppgåve i farmasi Avdeling for farmasøytisk biovitskap Farmasøytisk institutt Det matematiske-naturvitskaplege
Dei lysosomale cysteinproteasane legumain og cathepsin B i apoptose Eilen Tungland Mastergradsoppgåve i farmasi Avdeling for farmasøytisk biovitskap Farmasøytisk institutt Det matematiske-naturvitskaplege
ARBEIDSBESKRIVELSE Institutt for husdyr-og akvakulturvitenskap, NMBU
 EIDSBESKRIVELSE Institutt for husdyr-og akvakulturvitenskap, NMBU Metodenavn: Vannløselig karbohydrater (WSC) BIOVIT-nr.: Arb1014 1. Prinsipp Prøvene ekstraheres i acetatbuffer i romtemperatur over natt,
EIDSBESKRIVELSE Institutt for husdyr-og akvakulturvitenskap, NMBU Metodenavn: Vannløselig karbohydrater (WSC) BIOVIT-nr.: Arb1014 1. Prinsipp Prøvene ekstraheres i acetatbuffer i romtemperatur over natt,
BIOS 1 Biologi
 . Figurer kapittel 10: Menneskets immunsystem Figur s. 281 En oversikt over immunsystemet og viktige celletyper.> Immunsystemet Uspesifikt immunforsvar Spesifikt immunforsvar Ytre forsvar: hindrer mikroorganismer
. Figurer kapittel 10: Menneskets immunsystem Figur s. 281 En oversikt over immunsystemet og viktige celletyper.> Immunsystemet Uspesifikt immunforsvar Spesifikt immunforsvar Ytre forsvar: hindrer mikroorganismer
Bakgrunn Kvinner føder barn på et senere tidspunkt i livet enn tidligere Føder færre barn enn tidligere. Alder og fertilitet. Årsaker: Eggkvantitet
 Alder og fertilitet Mette Haug Stensen Senior klinisk embryolog Laboratoriesjef @ Bakgrunn Kvinner føder barn på et senere tidspunkt i livet enn tidligere Føder færre barn enn tidligere 23.05.14 Fakta:
Alder og fertilitet Mette Haug Stensen Senior klinisk embryolog Laboratoriesjef @ Bakgrunn Kvinner føder barn på et senere tidspunkt i livet enn tidligere Føder færre barn enn tidligere 23.05.14 Fakta:
Oppgave: MED2200-2_OPPGAVE1_H16_ORD
 Side 2 av 38 Oppgave: MED2200-2_OPPGAVE1_H16_ORD Del 1: OM RUS OG RUSS Jeanette er 19 år og russ. Hun og flere i hennes familie har familiær hyperkolesterolemi, en sykdom som gjør at pasienten har høyt
Side 2 av 38 Oppgave: MED2200-2_OPPGAVE1_H16_ORD Del 1: OM RUS OG RUSS Jeanette er 19 år og russ. Hun og flere i hennes familie har familiær hyperkolesterolemi, en sykdom som gjør at pasienten har høyt
NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR FYSIKK EKSAMEN I EMNE TFY4260 CELLEBIOLOGI OG CELLULÆR BIOFYSIKK
 Side av 1 av5 NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR FYSIKK EKSAMEN I EMNE TFY4260 CELLEBIOLOGI OG CELLULÆR BIOFYSIKK Faglig kontakt under eksamen: Catharina Davies Tel 73593688 eller
Side av 1 av5 NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR FYSIKK EKSAMEN I EMNE TFY4260 CELLEBIOLOGI OG CELLULÆR BIOFYSIKK Faglig kontakt under eksamen: Catharina Davies Tel 73593688 eller
Bruksanvisning IVD Matrix HCCA-portioned
 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
Sikkerhetsrisiko:lav. fare for øyeskade. HMS ruoner
 Reaksjonskinetikk. jodklokka Risiko fare Oltak Sikkerhetsrisiko:lav fare for øyeskade HMS ruoner Figur 1 :risikovurdering Innledning Hastigheten til en kjemisk reaksjon avhenger av flere faktorer: Reaksjonsmekanisme,
Reaksjonskinetikk. jodklokka Risiko fare Oltak Sikkerhetsrisiko:lav fare for øyeskade HMS ruoner Figur 1 :risikovurdering Innledning Hastigheten til en kjemisk reaksjon avhenger av flere faktorer: Reaksjonsmekanisme,
När och hur kan man mäta koagulationsfaktor III?
 När och hur kan man mäta koagulationsfaktor III? Carola Elisabeth Henriksson seksjonsleder, overlege Seksjon for hemostase og trombose Avdeling for medisinsk biokjemi Oslo Universitetssykehus, Rikshospitalet,
När och hur kan man mäta koagulationsfaktor III? Carola Elisabeth Henriksson seksjonsleder, overlege Seksjon for hemostase og trombose Avdeling for medisinsk biokjemi Oslo Universitetssykehus, Rikshospitalet,
Stamceller og blodcelledannelse
 Stamceller og blodcelledannelse Haakon Breien Benestad Avd. Fysiologi, IMB, UiO h.b.benestad@medisin.uio.no Dagens første tema Blodcelledannelse fra stamceller Noen reguleringsmekanismer for blodcelledannelsen
Stamceller og blodcelledannelse Haakon Breien Benestad Avd. Fysiologi, IMB, UiO h.b.benestad@medisin.uio.no Dagens første tema Blodcelledannelse fra stamceller Noen reguleringsmekanismer for blodcelledannelsen
Fritt kalsium i serum ph-korrigeres eller ikke?
 Foto: Morten Rakke Photography Fritt kalsium i serum ph-korrigeres eller ikke? Helle Borgstrøm Hager Avdelingsoverlege Sentrallaboratoriet Sykehuset i Vestfold Vårmøte i medisinsk biokjemi 2017 Fordeling
Foto: Morten Rakke Photography Fritt kalsium i serum ph-korrigeres eller ikke? Helle Borgstrøm Hager Avdelingsoverlege Sentrallaboratoriet Sykehuset i Vestfold Vårmøte i medisinsk biokjemi 2017 Fordeling
HVEM HAR ETTERLATT DNA?
 HVEM HAR ETTERLATT DNA? Hensikt Hensikten med forsøket er å utvikle en grunnleggende forståelse for DNA fingeravtrykksanalyser basert på restriksjonsenzymer og deres bruk i rettsmedisinske undersøkelser.
HVEM HAR ETTERLATT DNA? Hensikt Hensikten med forsøket er å utvikle en grunnleggende forståelse for DNA fingeravtrykksanalyser basert på restriksjonsenzymer og deres bruk i rettsmedisinske undersøkelser.
Nasjonal holdbarhetsdatabase
 Nasjonal holdbarhetsdatabase hvordan kan vi finne ut hvor lenge prøvene er holdbare? Preanalyse 10. 11.mai 2017 Nasjonalt prosjekt for standardisering av holdbarhetsforsøk Protokoll med beskrivelse av
Nasjonal holdbarhetsdatabase hvordan kan vi finne ut hvor lenge prøvene er holdbare? Preanalyse 10. 11.mai 2017 Nasjonalt prosjekt for standardisering av holdbarhetsforsøk Protokoll med beskrivelse av
Kan laboratoriene stole på sine kontrollmaterialer?
 Kan laboratoriene stole på sine kontrollmaterialer? Bioingeniørkongressen 2. juni 2016 Mari Solhus Produktspesialist/bioingeniør SERO AS Utfordringer med bruk av kvalitetskontroller Konsekvenser for laboratoriedriften
Kan laboratoriene stole på sine kontrollmaterialer? Bioingeniørkongressen 2. juni 2016 Mari Solhus Produktspesialist/bioingeniør SERO AS Utfordringer med bruk av kvalitetskontroller Konsekvenser for laboratoriedriften
NYHETSAVIS NR. 1/2000 Mai 2000
 NYHETSAVIS NR. 1/2000 Mai 2000 INNHOLD: Analysenytt Nye analyser akkreditert Endret referanseområde for serum testosteron og SHBG hos menn Prøvetaking til PTH og Ca 2+ Kommentarer Prioritering ved for
NYHETSAVIS NR. 1/2000 Mai 2000 INNHOLD: Analysenytt Nye analyser akkreditert Endret referanseområde for serum testosteron og SHBG hos menn Prøvetaking til PTH og Ca 2+ Kommentarer Prioritering ved for
Molekylære mekanismer ved Alzheimers sykdom (AD)
") Medisin stadium 1C, Geir Slupphaug, IKM Molekylære mekanismer ved Alzheimers sykdom (AD) De underliggende molekylære årsakene til AD er fremdeles mangelfullt kartlagt, men det synes nå klart at AD tilhører
Medisin stadium 1C, Geir Slupphaug, IKM Molekylære mekanismer ved Alzheimers sykdom (AD) De underliggende molekylære årsakene til AD er fremdeles mangelfullt kartlagt, men det synes nå klart at AD tilhører
Stiftelsen KG Jebsen Senter for B-cellekreft
 Stiftelsen KG Jebsen Senter for B-cellekreft Professor Ludvig Munthe Senterleder ludvig@uio.no B cellekreft Lymfekreft (Lymfomer) NHL, HL Blodkreft (Leukemier): B-ALL CLL Benmargskreft (myelomatose) Livstidsrisiko
Stiftelsen KG Jebsen Senter for B-cellekreft Professor Ludvig Munthe Senterleder ludvig@uio.no B cellekreft Lymfekreft (Lymfomer) NHL, HL Blodkreft (Leukemier): B-ALL CLL Benmargskreft (myelomatose) Livstidsrisiko
Bakgunn/historie. Memory loss, loss of. incontinence, confined to bed. Nevropatologi bestående av nevrofibrillære floker og
 Bakgunn/historie Nevropatologi bestående av nevrofibrillære floker og amyloide lid plakk lkk Alois Alzheimer Über eine eigenartige Erkrankung der Hirnrinde, 1907: Memory loss, loss of orientation, incontinence,
Bakgunn/historie Nevropatologi bestående av nevrofibrillære floker og amyloide lid plakk lkk Alois Alzheimer Über eine eigenartige Erkrankung der Hirnrinde, 1907: Memory loss, loss of orientation, incontinence,
KROPPENS FORSVARSMEKANISMER
 KROPPENS FORSVARSMEKANISMER Immunitet = fri for, uberørt av Immunologi Immunsystemet Uspesifikke forsvarsmekanismer Ytre- og indre forsvarslinje Spesifikke forsvarsmekanismer Cellulære forsvar (T-celler)
KROPPENS FORSVARSMEKANISMER Immunitet = fri for, uberørt av Immunologi Immunsystemet Uspesifikke forsvarsmekanismer Ytre- og indre forsvarslinje Spesifikke forsvarsmekanismer Cellulære forsvar (T-celler)
ROLLEN TIL UROKINASE PLASMINOGEN AKTIVATOR RESEPTOREN (upar) FOR UTTRYKK AV MOLEKYLER INVOLVERT I CELLE ADHESJON, MIGRASJON OG INVASJON.
 FOR UTTRYKK AV MOLEKYLER INVOLVERT I CELLE ADHESJON, MIGRASJON OG INVASJON.") ROLLEN TIL UROKINASE PLASMINOGEN AKTIVATOR RESEPTOREN (upar) FOR UTTRYKK AV MOLEKYLER INVOLVERT I CELLE ADHESJON, MIGRASJON OG INVASJON. Av: Stine Haustreis Mai 2010 Veiledere: Førsteamanuensis Gunbjørg
ROLLEN TIL UROKINASE PLASMINOGEN AKTIVATOR RESEPTOREN (upar) FOR UTTRYKK AV MOLEKYLER INVOLVERT I CELLE ADHESJON, MIGRASJON OG INVASJON. Av: Stine Haustreis Mai 2010 Veiledere: Førsteamanuensis Gunbjørg
TBT4135 Biopolymerkjemi Laboratorieoppgave 4: Analyse av løselighet og utfelling Gruppe 5
 TBT4135 Biopolymerkjemi Laboratorieoppgave 4: Analyse av løselighet og utfelling Gruppe 5 Hilde M. Vaage hildemva@stud.ntnu.no Malin Å. Driveklepp malinad@stud.ntnu.no Oda H. Ramberg odahera@stud.ntnu.no
TBT4135 Biopolymerkjemi Laboratorieoppgave 4: Analyse av løselighet og utfelling Gruppe 5 Hilde M. Vaage hildemva@stud.ntnu.no Malin Å. Driveklepp malinad@stud.ntnu.no Oda H. Ramberg odahera@stud.ntnu.no
Lab forelesning. C-vitamin. Enzymer i hverdagen
 Lab forelesning C-vitamin Enzymer i hverdagen C-vitamin eller askorbinsyre Finnes i svært mange frukter og grønnsaker Viktige kilder: appelsin paprika poteter C-vitamin Har mange viktige funksjoner i kroppen
Lab forelesning C-vitamin Enzymer i hverdagen C-vitamin eller askorbinsyre Finnes i svært mange frukter og grønnsaker Viktige kilder: appelsin paprika poteter C-vitamin Har mange viktige funksjoner i kroppen
QIAsymphony SP protokollark
 QIAsymphony SP protokollark DNA_Buffy_Coat_200_V7_DSP-protokoll Generell informasjon Til in vitro-diagnostisk bruk. Denne protokollen brukes til rensing av totalt genomisk og mitokondrielt DNA fra frisk
QIAsymphony SP protokollark DNA_Buffy_Coat_200_V7_DSP-protokoll Generell informasjon Til in vitro-diagnostisk bruk. Denne protokollen brukes til rensing av totalt genomisk og mitokondrielt DNA fra frisk
Medikamentell Behandling
 www.printo.it/pediatric-rheumatology/no/intro Medikamentell Behandling Versjon av 2016 13. Biologiske legemidler Gjennom bruk av biologiske legemidler har nye behandlingsprinsipper mot revmatisk sykdom
www.printo.it/pediatric-rheumatology/no/intro Medikamentell Behandling Versjon av 2016 13. Biologiske legemidler Gjennom bruk av biologiske legemidler har nye behandlingsprinsipper mot revmatisk sykdom
Emne:HSB1209 Biomedisinsk analyse Lærer/telefon: Gerd 11.Breivik /
 Høgskoleni Østfold 1 Avdelingfor ingeniørfag EKSAMENSOPPGAVE Emne:HSB1209 Biomedisinsk analyse Lærer/telefon: Gerd 11.Breivik 69303052/99706354 Grupper: 13B10/13BIOY Dato: 05.06.14 Tid: 09.00 13.00 Antall
Høgskoleni Østfold 1 Avdelingfor ingeniørfag EKSAMENSOPPGAVE Emne:HSB1209 Biomedisinsk analyse Lærer/telefon: Gerd 11.Breivik 69303052/99706354 Grupper: 13B10/13BIOY Dato: 05.06.14 Tid: 09.00 13.00 Antall