Silika nanopartikler endrer morfologi og øker glutamat cystein ligase - promotoraktiviteten i PC12-celler
|
|
|
- Vegar Simensen
- 7 år siden
- Visninger:
Transkript


1 Silika nanopartikler endrer morfologi og øker glutamat cystein ligase - promotoraktiviteten i PC12-celler Taqwa Farajalla Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige fakultet UNIVERSITETET I OSLO Juni 2015 I
2 II
3 Masteroppgave for graden Master i farmasi Silika nanopartikler endrer morfologi og øker glutamat cystein ligase - promotoraktiviteten i PC12-celler Taqwa Farajalla Oppgaven er utført ved Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige fakultet Universitet i Oslo Interne veiledere: Professor Ragnhild Elisabeth Paulsen Postdoktor Kirsten Eline Rakkestad Ekstern veileder: Professor II Marit Låg III
4 Copyright Taqwa Farajalla 2015 Silika nanopartikler endrer morfologi og øker glutamat cystein ligase-promotoraktiviteten i PC12-celler Taqwa Farajalla Trykk: Reprosentralen, Universitetet i Oslo IV
5 Forord Det er alltid noen personer som står bak en suksess. Først vil jeg takke min mor, far og ektemannen for all støtten jeg fikk i studieårene. Dette har jeg satt stor pris på. Jeg takker Professor Ragnhild Elisabeth Paulsen og Postdoktor Kirsten Eline Rakkestad for den gode veiledningen og støtten jeg fikk i gjennomføring av denne oppgaven. Jeg må si at jeg aldri kommer til å glemme deres hjelp. I tillegg vil jeg takke stipendiat Lars Peter Austdal og avdelingsingeniøren Mona Gaarder. Dere har vært til god hjelp på labben. Oslo, juni 2015 Taqwa Farajalla V
6 VI
7 Sammendrag Effektiv levering av legemidler til hjernen er utfordrende på grunn av beskyttende barrierer. Blood-hjernebarrieren danner den største hindringen for levering av legemidler til sentralnervesystemet. Flere metoder for mer effektiv legemiddellevering er derfor i stadig utvikling. Nanopartikler har på grunn av sine unike egenskaper som modifiserbare partikkeloverflater, stort overflateareal i forhold til volum, ulike strukturer og størrelser og optiske egenskaper evne til å bære ulike legemidler og sørge for effektiv legemiddellevering til hjernen. Silika nanopartikler er biodegraderbare nanopartikler med som syntetiseres lett, er kostnadseffektive, har høyt overflateareal og lett modifiserbar overflate, er reaktive og har høy legemiddelbæreevne. Disse nanopartiklene er formulert for flere medisinske formål inkludert cellulær og ikke-virusbasert genlevering, legemiddellevering, diagnostisering og avbildning i CNS. For at silika nanopartikler skal kunne brukes effektivt må deres neurotoksisitetsprofil klarlegges og vurderes nøye. I denne masteroppgaven ble PC12-celler benyttet som modell for å studere eventuelle toksiske effekter av 10 nm og 50 nm silika nanopartikler. Silika nanopartikler har vært involvert i morfologiske forandringer og økt celledød. PC12-celler eksponert for silika nanopartikler ble derfor analysert morfologisk og det ble undersøkt om nanopartiklene ville føre til økt celledød i PC12-celler. I tillegg ble det undersøkt om silika nanopartikler påvirket glutamat cystein ligase-promotoraktiviteten siden de i tidligere studier har vist induksjon av oksidativ stress. Ettersom silika nanopartikler har vært involvert i morfologiske forandringer ble MMP9, Gli og P53-promotoraktiviteten også studert. Noen nanopartikkeltyper har vist å kunne aktivere østrogen kjernereseptorer. Studier av ERE-promotoraktiviteten ble derfor også inkludert. Metoden luciferase reporte assay ble benyttet for promotoraktivitetsstudier. Både 10 nm og 50 nm silika nanopartikler førte til morfologiske forandringer i form av avlange, trådlignende PC12-celler og begge førte til økt celledød. 10 nm silikananopartikler i lav konsentrasjon økte GCS-promotoraktiviteten og oksidativ stress indusert av silika nanopartiklene ble foreslått som mekanismen bak dette. Nanopartiklene viste også østrogenlignende effekter og førte til økt aktivitet av østrogenreseptorer. VII
8 Resultater fra denne studien viser en del toksiske effekter mediert av silika nanopartikler. Dette i tillegg til toksiske effekter rapportert fra andre studier gjør at videre forskning for klargjøring av neurotoksisitetsprofilen er nødvendig. VIII
9 Forkortelser ANOVA BSA DMEM DMSO DTT ER ERE FBS HS MQ NAC NaCl NGF NP P PBS PEG PS RLU ROS SE Si Si10 Si50 z-devd-cmk Variansanalyse (Analysis of variance) Bovint serum albumin Dulbecco s Modified Eagle s medium Dimetylsulfoksid Dithiothreitol Østrogen kjernereseptorer Østrogen responselement Føtalt kalveserum Hesteserum Milli Q N-Acetyl-L-cysteine Natriumklorid Nervevekstfaktor Nanopartikler Natriumpyruvat Fosfatbufret saltvann Polyethylene glykol Penicillin-Streptomycin Relativ luminescensenhet Reaktive oksygenspesier Standardfeil Silika nanopartikler Silika nanopartikler med diameter 10 nm Silika nanopartikkel med diameter 50 nm z-aspartat-glutamat-valin-aspartatklormetylketon IX
10 X
11 Innholdsfortegnelse 1 INNLEDNING Sentral nervesystemet Barrierer i sentralnervesystemet Strategier for levering av legemidler til sentral nervesystemet Nanopartikler Nanopartikkelbasert legemiddellevering Unike egenskaper ved nanopartikler Silika nanopartikler Toksisitet av silika nanopartikler Oksidativt stress og GCL MMP Gli P ER Målet med studien Materialer og metoder Oversikt over kjemikalier, biologiske produkter, nanopartikler og utstyr PC12-celler PC12-celler Splitting av PC12-celler til flaske Splitting av PC12-celler til skåler Eksponering av PC12-celler Transfeksjon Transfeksjon av PC12-celler Luciferase reporter assay Ildflueluciferase Høsting av celler til luciferasemåling Måling av ildflueluciferaseaktivitet Renillaluciferase Måling av renillaluciferaseaktivitet Morfometrisk analyse i mikroskop Deteksjon av celledød Prosedyre for farging med trypanblå Statistiske analyser Resultater Morfometrisk analyse Celledød Celledød ved bruk av batch 1 nanopartikler Celledød ved bruk av batch 2 nanopartikler Promotoraktivitet studier GCS-promotoraktivitet i silika nanopartikkel eksponerte PC12-celler Effekt av silika nanopartikler på MMP9-promotoraktivitet i PC12-celler Effekt av silika nanopartikler på Gli-promotoraktivitet i PC12-celler med og uten NAC P53-promotoraktivitet i silika nanopartikkel eksponerte PC12-celler XI
12 3.3.5 Effekt av silika nanopartikler på ERE-promotoraktivitet i PC12-celler Effekt av silika nanopartikler på ERE-promotoraktivitet i PC12-celler Celledød Celledød ved bruk av batch 1 nanopartikler Celledød ved bruk av batch 2 nanopartikler Diskusjon Diskusjon av metoder Valg av PC12-celler som model Valg av eksponeringer Luciferaseassay som metode for å studere promotoraktivitet Valg av kvalitativ morfometrisk analyse Deteksjon av celledød som endepunkt Diskusjon av biologiske funn Morfologisk forandring i silika nanopartikkel eksponerte PC12-celler Økt GCS-promotoraktivitet i silika nanopartikkel eksponerte PC12-celler Silika nanopartiklers påvirkning på Gli-promotoraktivitet Silika nanopartikler viste tendens til å redusere p53-promotoraktiviteten Økt ER aktivitet Konklusjon Litteraturliste Vedlegg / Appendiks XII
13
14
15 1 INNLEDNING Bruk av nanopartikler i medisin, spesielt som legemiddelbærere er i stekt økende grad [1]. Silika nanopartiker har på grunn av sine unike egenskaper potensial for bruk i medisin og aktiv forskning er på gang med tanke på dem som ideelle bærere av legemidler [2]. 1.1 Sentral nervesystemet Hjernen er en av de mest sensitive organene i kroppen. Den er godt beskyttet med flere barrierer som danner strenge begrensninger i passasje av skadelige toksiske stoffer. Til tross for nytten disse barrierene innebærer, er de også et hinder for effektiv levering av legemidler til hjernen. Effektiv behandling av lidelser i sentralnervesystemet (CNS) er derfor utfordrende [3]. Selv om det finnes effektive legemidler for behandling av en del CNS lidelser, er det begrensninger i levering av disse legemidlene til målstedet i hjernen med en konsentrasjon nødvendig for deres optimale effekt, som gjør at behandling av en del CNS lidelse ikke er under kontroll [4]. Flere metoder for mer effektiv levering av legemidler via systemisk sirkulasjon er derfor i stadig utvikling [3] Barrierer i sentralnervesystemet Blod-hjernebarrieren, blod- cerebrospinalvæskebarrieren og blod- tumorbarrieren er barrierene som begrenser levering av legemidler og effektiv behandling av lidelser i CNS [3]. Blod-hjernebarrieren danner størst hindring for levering av legemidler til CNS [5]. Denne barrieren separerer blodet fra ekstracellulær matriks i hjernen. Tight junctions mellom endotelcellene i blodkapillærer danner grunnlaget for denne barrieren som fører til begrensninger i passasje av substanser. I utgangspunktet kan bare små molekyler (maksimum 400 Da), små hydrofile substanser (maksimum 150 Da) og lipofile substanser (maksimum 400 Da) diffundere passivt gjennom blod-hjernebarrieren, men det er i tillegg flere andre faktorer som begrenser passasjen. Større molekyler og hydrofile substanser kan bare passere blod-hjernebarrieren via aktive transportsystemer, dersom de har affinitet til disse. Flere 1

16 effluks pumper som P-glykoprotein pumpen (Pgp), Multidrug resistens assosierte proteiner (MRPs) og brystkreft resistens proteiner (BCRP), i endotel celler begrenser passasje av legemidler til hjernen. Forskjellige protein degraderende enzymer i endotel cellene er også en begrensende faktor [6] Strategier for levering av legemidler til sentral nervesystemet To hovedstrategier for levering av legemidler til CNS eksisterer: invasiv og ikke invasiv levering. Oversikt over metoder innen hver strategi er vist på figur1.1. Figur 1.1 oversikt over strategier og metoder innen hver strategi, for levering av legemidler til CNS. Hovedstrategier deles i invasive og ikke invasive. Invasive metoder er kirurgi, modulering av blod-hjernebarriere permeabilitet og direkte injeksjon av legemiddel til målstedet i hjernen. Ikke invasive metoder er biologisk metode (bruk av legemiddel bærer, nanopartikler), kjemisk modifisering av legemiddel og levering av legemiddel via luktenervene i nesen. Figur er hentet fra [5]. Invasive metoder innebærer fysisk legemiddel levering via kirurgi, direkte injeksjon av legemidler til målstedet eller metoder som modulerer blod-hjernebarriere permeabiliteten slik at legemidler kan passere gjennom barrieren. Disse invasive metodene er risiko fulle og uegnet til flergangsbehandling av lidelser med behov for flere behandlinger. Ikke invasive metoder innebærer passasje av legemidler gjennom blod-hjernebarrieren uten modulering av barrieren eller transport av legemidler utenom passasje av barrieren, via luktenervene i neseslimhinnen. legemidler med kjemiske egenskaper som gjør at de ikke er i stand til å passere blodhjernebarrieren, kan moduleres kjemisk eller syntetiseres i prodrug form som er egnet til å 2
![passerer barrieren i en inaktiv form av legemidlet og bli omdannet til aktivt legemiddel i hjernen [7].](/docs-images/62/47202653/images/17-0.jpg "En annen ikke invasiv metode utviklet i det siste er levering av legemidler til CNS ved bruk av nanoteknologi, mer spesifikk nanopartikler (NP) [8]. 1.")
17 passerer barrieren i en inaktiv form av legemidlet og bli omdannet til aktivt legemiddel i hjernen [7]. En annen ikke invasiv metode utviklet i det siste er levering av legemidler til CNS ved bruk av nanoteknologi, mer spesifikk nanopartikler (NP) [8]. 1.2 Nanopartikler Nanopartikler karakteriseres som partikler med diameter dimensjon som ligger mellom 1 og 100 nanometer størrelse. Dette har stor betydning for NP egenskaper i forhold til større partikler. Figur 1.2 demonstrerer størrelsesorden for en del nanopartikler [9]. Figur 1.2. Sammenligning av størrelsen på en del nanopartikler med kjente stoffer i naturen. [1] liposom, [2] solid lipid NP, [3] polymer micelle, [4] dendrimer, [5] quantum dot (Qdot) og [6] jernoksid NP i forhold til (fra venstre) vannmolekyl, DNA, bakteriofag, virus, bakterie og nervecelle. Figuren er hentet fra [9]. 3
18 1.2.1 Nanopartikkelbasert legemiddellevering Målet med formulering av nanopartikler for medisinske formål er ideelle legemiddelbærere med høy sikkerhet som har høy bæreevne, kan effektiv passere fysiologiske barrierer for en spesifikk levering og effektiv behandling av lidelser [10]. På grunn av sine unike egenskaper har NP-er evne til å bære ulike legemidler og sørge for effektiv passasje gjennom blod-hjernebarrieren og levering til hjernen. Denne metoden for transport av legemidler gjennom blod-hjernebarrieren er avhengig av optimalisert funksjonalisering av NP, ikke modifisering av legemidlers kjemiske egenskaper [8]. Med passende overflatefunksjonalisering kan et legemiddel rettes mot bestemte steder i CNS [6]. Dette er av spesiell betydning for behandling av aggressive hjernetumorer [11]. Legemidler kan assosieres med NP-er, via kovalent binding til NP eller uten kovalent binding (figur 1.3). I assosiasjon uten kovalent binding, kan legemidler enten innkapsles i NP eller sitte stabilt på overflaten. Innkapslet legemiddel frigjøres ved en forandring i Ph, temperatur eller en annen ekstern stimuli, som gjør NP-et permeabel for legemiddel frigjøring. Dersom legemidlet er stabilisert på overflaten kan en forandring i miljøet i kroppen frigjøre legemidlet, for eksempel et hydrofobt legemiddel frigjøres ved cellemembraner som er hydrofob. Fordeler med transport av legemiddel uten kovalent binding med NP er: 1. det er ikke behov for modifisering av legemiddel, noe som bevarer effektiviteten 2. en optimalisert assosiasjon kan brukes for andre lignende legemidler med samme egenskaper 3. det er ikke behov for en ekstern stimulus for brytning av kovalent binding. Når legemidlet er kovalent bundet til NP må en ekstern stimulus ha ansvaret for å bryte bindingen, men dette innebærer også en fordel da stedsuspesifikk levering hindres [12]. 4
 faktorer som kan brukes for kontrollert frisetting. Grøn indikerer legemiddel. Gult indikerer NP. Figur hentet fra [12]")
19 Figur 1.3. Nanopartikkelbasert legemiddellevering. A) kovalent eller ikke kovalent binding. B) Stabilisering av legemiddel på overflaten eller innkapsling. C) faktorer som kan brukes for kontrollert frisetting. Grøn indikerer legemiddel. Gult indikerer NP. Figur hentet fra [12] Unike egenskaper ved nanopartikler NP-er har flere unike egenskaper som er ansvarlig for den store oppmerksomheten på deres invaderende rolle i medisin [12]. Modifiserbar partikkeloverflate: Nanopartikler har modifiserbare overflater som gjør det mulig å designe dem med flere funksjoner for ulike medisinske formål (figur 1.4). Som beskrevet tidligere kan legemidler både innkapsles eller konjugeres på overflaten. Hydrofile surfaktanter som polyetylenglykol 5

20 (PEG) kan konjugeres på overflaten for å øke partiklenes halveringstid og stabilitet i blodbanen. Dette begrenser adsorpsjon av proteiner i systemisk sirkulasjon til overflaten, som kan føre til at partiklene oppdages som fremmede legemer og fjernes av retikuloendoteliale systemet, noe som gir partiklene kort halveringstid. Med kovalent binding av ulike antistoffer, peptider, aptamer (små RNA/DNA molekyler som kan bindes til spesifikke proteiner eller andre målsteder i celler) og kationiske molekyler på NP overflaten kan et legemiddel virke på et spesifikt sted i hjernen. Man kan i tillegg binde fluorescerende prober på overflaten og på denne måten kunne spore legemidlet i hjernen[13] [8]. Figur 1.4 Multifunksjonalisering av nanopartikler. Legemidler kan både innkapsles eller bindes på overflaten av NP. Binding av hydrofile surfaktanter som polyetylenglykol (PEG) øker NP halveringstid. Kovalent binding av antistoffer, peptider, aptamer og kationiske molekyler muliggjør stedspesifikk virkning av legemidler i hjernen. Med binding av fluorescerende prober kan legemidler spores i hjernen. Figur er hentet fra [8]. Stort overflateareal i forhold til volum: Store partikler har liten overflateareal i forhold til volumet, i tillegg er det en maksimum grense i antall partikler i et bestemt volum. NP-er derimot har stort overflateareal i forhold til volum og de kan syntetiseres i forskjellige størrelser, slik at antall NP-er i et bestemt volum kan justeres ved justering av partikkelstørrelse. Dette har stor betydning for optimalisering av NP-er brukt som legemiddelbærere [12]. Ulik struktur og størrelse: Nanopartikler kan syntetiseres i forskjellig form og størrelse, egenskaper som har betydning for deres biologiske funksjon. Ulike form på NP-er påvirker opptaket i cellene og ulik 6
21 størrelse bestemmer deres akkumulering og virkningssted. For eksempel setter NP-er med en viss størrelse seg på ujevne blodårer i tumorer og virker spesifikk på stedet, mens mindre NPer passerer disse blodårene [12]. Optiske egenskaper: NP har også unike optiske egenskaper som gjør at de er egnet for avbildning i kroppen [12]. Egenskaper på en ideell nanopartikkel som legemiddelbærer: Biokompatibilitet er en sentral egenskap Høy bære eller innkapslings evne Målstyring og legemiddellevering til spesifikk celle type/vev. Null prematur legemiddel frisetting Evne til kontrollert legemiddelfrisetting og riktig frisettingshastighet for å oppnå riktig konsentrasjon [14] 1.3 Silika nanopartikler Silika (SiO 2 ) nanopartikler (SiNP) er biodegraderbare metaloksid nanopartikler med spesielle fysisk og kjemiske egenskaper som gjør dem egnet for flere formål. De er små nanoskala partikler som syntetiseres lett, er kostnadseffektive, har høyt overflateareal og lett modifiserbar overflate, er høy reaktive og har høy legemiddel bæreevne. SiNP har blitt utviklet for bruk i blant annet kosmetikk, mekanisk polering, mat og for flere medisinske formål som molekylær avbildning, diagnose, målrettet legemiddel/gen/protein levering, kontrollert legemiddel frigjøring og kreftterapi [10, 15, 16]. For bruk i CNS er SiNP formulert for cellulær og ikke-virusbasert gen levering, legemiddel levering, diagnostisering og avbildning, fordi SiNP er vurdert å ha høyere biokompatibilitet grad enn andre NPer, for eksempel quantom dot som brukes for avbildning og kan inneholde toksiske metaller som kadmium og kvikksølv [17, 18]. For at SiNP skal kunne brukes effektivt må sikkerhet og toksisitetsprofilen klarlegges og vurderes nøye [10]. Disse NP er generelt vurdert som ikke toksiske, men deres neurotoksisitet 7
22 profil er uklart [16, 17]. Flere in vivo og in vitro studier har rapportert toksiske effekter av SiNP hvor forandring av cellulær morfologi, økt celledød, induksjon av oksidativ stress, aktivering av P53-signalveien og forstyrret cellesyklus er noen av dem [15, 16, 19]. 1.4 Toksisitet av silika nanopartikler Oksidativt stress og GCL Hjernen er et sensitivt organ mot oksidativ stress og økt nivå av reaktive oksygen radikaler kan føre til nevrodegenerasjon og dermed nevrodegenerative sykdommer som Alzheimer [18]. Rikelighet med umettede fettsyrer i hjernen gjør organet mer utsatt for lipid peroksidering. I tillegg er det høyt forbruk av oksygen i hjernen og redusert aktivitet av glutation peroksidase [2]. Oksidativ stress i celler oppstår ved ubalanse i produksjon av reaktive oksygen spesies (ROS) og kroppens antioksidant system. En viktig endogen antioksidant er glutation (GSH). Proteinet syntetiseres i cellene i to trinn der det hastighetsbestemmende enzymet glutamat cystein ligase (GCL) regulerer første trinn i syntesen. Enzymet kalles også gammaglutamylcystein syntetase (GCS). Forandring i GCS gen uttrykket fører til forandring i GSH nivået, slik at ved oppregulering av GCS gen øker glutaion syntesen og ved nedregulering reduseres glutation syntesen [20]. Silika nanopartikler har i flere studier vist å kunne føre til oksidativ stress i flere celle typer inkludert SK-N-SH humane neuroblastoma celler, N2a muse neuroblastoma celler, primære mikroglia celler fra rotte og PC12 celler [2, 17, 18, 21]. I PC12-celler har eksponering med SiNP ført til økt produksjon av reaktive oksygenspesier, redusert GSH nivå, økt lipid peroksidering og økt celledød, der lipidperoksidering av celle membraner ble foreslått som mekanismen for oksidativ stress indusert celledød [2]. I en in vivo studie der silika nanopartikler viste størst akkumulering i striatum via intranasalt administrasjon i rotter, førte partiklene til oksidativ stress i striatum [16]. 8
23 1.4.2 MMP9 Matrix metaloprotease-9 (MMP-9) er et medlem i en undergruppe, gelatinaser, av MMP familie av proteolytiske enzymer [22]. Disse proteasene er involvert i remodellering av ekstracellulær matriks både ved normale fysiologiske funksjoner og patologiske tilstander [23]. Under utvikling av CNS er disse enzymene sammen med en del andre enzymer ansvarlig for degradering av ekstracellulær matriks for å muliggjøre neuronal migrasjon og nevrittutvekst. I tillegg ved skade i perifere nevroner induseres uttrykket av MMP9 i schwann celler og makrofager som muliggjør gjenopprettelse av nevroner og aksoner [24]. I en studie hvor PC12 celler ble brukt som model for å studere MMP9 rolle i nevrittutvekst, ble akkumulering av MMP9 i skadete nevroner påvist og studien konkluderte med at MMP9 har en sentral rolle i nevritt utvekst. [25]. Flere studier har vist økt MMP9 nivå i metastaserende tumorer. Degradering av ekstra cellulær matriks rundt tumorer gir dem mulighet til å spre seg [26]. MMP-9 er ikke konstitutivt uttrykk. Promotorregionen i denne proteasen inneholder aktivator protein-1 sekvens (AP-1) [23]. AP-1 er en transkripsjonsfaktor som består av heterodimer proteiner fra jun og fos protein familien. Komplekset bindes til AP-1 bindingssete og regulerer gen transkripsjonen. Flere faktorer kan indusere jun og fos genuttrykket, for eksempel inflammatoriske cytokiner og onkogene produkter [23, 27]. Silika NP har i en studie vist å kunne føre til morfologiske forandringer i PC12 celler og påvirke nevritt utvekst. I studien hadde NGF+SiNP eksponerte PC12 celler mistet evnen til å utvikle modne nevritter i forhold til celler bare eksponert for NGF [2] Gli Gli er en familie av tre transkripsjonsfaktorer involvert i sonic hedgehog signalveien, en nødvendig signalvei for riktig utvikling av organer, inkludert CNS, under organogenesen [28]. I fullt utviklet CNS har signalveien sentral rolle for vedlikehold av neuronal stamceller [29]. 9
24 På cellenivå er signalveien viktig for celle proliferasjon og forstyrrelser i signalveien kan føre til ukontrollert celle proliferasjon som kan ende i tumorutvikling [30]. Sonic hedgehog signalveien har også vist å fremme tumor metastasering ved å redusere uttrykket av E- cadherin (transmembran proteiner som fører til binding av naboceller ved at de bindes til hverandre) og øke uttrykket av snail protein (transkripsjonsfaktor som hemmer E-cadherin uttryk) [30-32]. Signalveien aktiveres ved at sonic hedgehog proteiner setter seg på PTCH1 transmembran reseptorer som da aktiverer smoothened reseptorer. Aktiverte smoothened reseptorer fører til fosforylering av Gli som translokkeres til celle kjerne, bindes til promotor region av gener og regulerer transkripsjonen [28]. Gli 1 og Gli 2 virker som transkripsjonsaktivatorer, mens Gli 3 virker som transkripsjonssupressor. De fleste gener regulert av sonic hedgehog signalveien har gli1 sekvens og reguleres av Gli1 som er hovedfaktoren i signalveien. Gli1 er involvert i regulering av celle proliferasjon og celleorganisering [30] P53 P53 er en transkripsjonsfaktor som ved oksidativ stress i celler og DNAskade har en beskyttelses rolle på genomet [33]. P53 blir også referert til som et tumorsuppressorgen. I omtrent 50% av krefttilfellene har man sett mutasjoner i p53 genet, derfor har p53 aktivitet fått et særplass i flere studier [34]. Aktivering av p53 ved oksidativt stress eller DNA skader fører via flere mekanismer til enten apoptose eller reversibel arrestering av cellesyklus, reparasjon og celle proliferasjon igjen, viktige prosesser for hindring av tumor utvikling. Flere gener ansvarlige for apoptose eller arrestering av cellesyklus er responsive til p53 regulering og kan både nedreguleres eller oppreguleres [33]. Aktivert p53 aktiverer uttrykk av Bax, et proapoptotisk protein og medlem i Bcl-2 familien, og nedregulerer utrykk av antiapoptotisk Bcl-2. Dett fører til frigjøring av cytokrom c og medfølgende caspase-3- aktivering som ender i apoptose [19]. Ved store oksidative skader oppreguleres apoptotiske gener, mens ved moderate skader på DNA oppreguleres gener ansvarlige for arrestering av cellesyklus. I CNS har p53 i tillegg til regulering av nevronal apoptose og DNA reparasjon, en sentral rolle i nevritt utvekst og aksonal regenerasjon. Transkripsjonsfaktoren p53 regulerer utrykk av aktinbindende proteinet coronin 1b og GTPasen Rab 13, to nødvendige proteiner for nevrittutvekst. I en studie hvor PC12 celler ble brukt som model viste celler eksponert for 10
25 nervevekstfaktor (NGF) ved fravær av p53 ikke utvikling i nevritt utvekst i forhold til NGF eksponerte celler med tilstedværelse av p53. Transkripsjonsfaktoren er derfor en nødvendig faktor for NGF mediert nevrittutvekst i PC12 celler. Samme studie viser nødvendigheten av p53 for nevtittutvekst og modning av embryonale kortikal nevroner fra rotte [35]. Silika NP har vist å kunne aktivere p53 og en del andre signalveier via oksidativ stress. I en studie har eksponering av endotelceller med SiNP, med tanke på at partiklene har en direkte kontakt med endotel celler i sirkulasjonssystemet, vist å kunne indusere produksjon av reaktive oksygen radikaler med medfølgende aktivering av p53, endotelcelle dysfunksjon og apoptose [19]. I en annen studie har SiNP via induksjon av oksidativ stress i PC12 celler, vist et doseavhengig uttrykk av p21 og Gadd45 proteinene som aktiveres av p53 og regulerer arrestering av celle syklus. Det var i tillegg økt uttrykk av bax og redusert uttrykk av bcl-2 som da fører til apoptose [16] ER Hormonet østrogen er et viktig hormon for riktig funksjon av flere organer, inkludert CNS. Hormonet medierer sin effekt via østrogen kjernereseptorer (ER), viktige transkripsjonsfaktorer som regulerer transkripsjon av flere gener. Disse reseptorene kan aktiveres av hormonene 17α- østradiol, 17β-østradiol og en del andre faktorer involvert i andre signalveier. 17β-østradiol er det mest aktive østrogenet som har sterkere affinitet for østrogen reseptorer. Dette hormonet syntetiseres i flere organer inkludert hjerne [36]. 17βøstradiol medierer sin effekt via binding til østrogen reseptorer i cytoplasma, som da fører til aktivering av reseptoren som translokeres til kjernen, binder seg til østrogen responselement (ERE) i promotorregion og regulerer transkripsjon av flere gener [37]. En del nanopartikler og miljøtoksiner er vist å kunne aktivere østrogen reseptorer på samme måte som østrogener [38, 39]. En studie viste at nanopartikler fra dieseleksos kunne aktivere ER i MCF-7 celler (en brystkreft cellelinje) og føre til stimulering av celleproliferasjon på samme måte som 17β-østradiol [39]. 11
26 1.5 Målet med studien Hovedmål: Undersøke videre uønskede effekter av silika nanopartikler i PC12-celler. Delmål: 1. Undersøke eventuelle forandringer i PC12-cellers morfologi ved eksponering for silika nanopartikler. 2. Undersøke om silika nanopartikler fører til økt celledød i PC12-celler og videre prøve å finne ut ansvarlig signalvei for celledød. 3. Undersøke om silika nanopartikler påvirker GCS, MMP9, Gli, P53 og EREpromotoraktiviteten. 12
27 2 Materialer og metoder 2.1 Oversikt over kjemikalier, biologiske produkter, nanopartikler og utstyr Tabell 2.1 Kjemikalier og biologiske produkter Produkt L-Ascorbic acid Bovint serumalbumin (BSA) (A9418) Dithiothreitol (DTT) DMSO Dulbecco s Modified Eagle s medium (DMEM) Dual Luciferase Reporter Assay System Etanol (absolutt alkohol prima) Føtalt kalveserum (FBS) Glutamin Hesteserum (HS) D-luciferin Metafectene Pro N-Acetyl-L-cysteine (NAC) Natriumklorid (NaCl) Natriumpyruvat NGF (nervevekstfaktor) Penicillin-Streptomycin ( U/ml- 10 mg/ml) Raloxifene SB (P38-hemmer) Triton Trypanblå Leverandør Sigma- Aldrich, St.Louis, USA Sigma- Aldrich, St.Louis, USA Sigma- Aldrich, St.Louis, USA Sigma- Aldrich, St.Louis, USA Gibco, Life Technologies, Carlsbad, USA Promega Corporation, USA Kemetyl Norge AS, Vestby, Norge Lonza, Walkersville, USA Sigma- Aldrich, St.Louis, USA Gibco, Life Technologies, Carlsbad, USA Duchefa Biochemistry, Nederland Biontex laboratories, Tyskland Sigma-adrich, Merck KGaA Darmstad, Tyskland Gibco, Life Technologies, Carlsbad, USA Sigma- Aldrich, St.Louis, USA Gibco, Life Technologies, Carlsbad, USA Tocris, Bristol, England Sigma-Aldrich, St. Louis, USA Sigma-Aldrich, St. Louis, USA Sigma-Aldrich, St. Louis, USA 13
28 (±)-α-tocopherol z-devd-cmk (Caspase-3-hemmer) Sigma-Aldrich, St.Louis, USA Bachem AG, Bubendorf, Sveits Tabell 2.2 Nanopartikler Nanopartikkel Si10 Si50 Leverandør Kisker Biotech GmbH & Co. KG, Steinfort, Germany Kisker Biotech GmbH & Co. KG, Steinfort, Germany Tabell 2.3 Utstyr Utstyr Cellekulturflaske 75 cm 2 Cellekulturskåler 8,8 cm 2 og 21,5 cm 2 Celleskrape CO 2 -inkubatorskap Gassbrenner Tellekammer Kamera til lysmikroskop Kjølesentrifuge (Hergeus Fresco 21) Varmeapparat 37 C LAF-benk (Holten LaminAir, modell 1.2) Luminometer (Lumat LB 9507) Lysmikroskop (TMS 2.2.1) Magnetrører Pasteurpipette Pipetboy Pipetter Leverandør Nunc, Danmark Nunc, Danmark Corning Incorporated, USA Thermo Scientific Rockford, USA Fireboy eco, Integra Biosciences Zizers, Sveits neubauer Spot, USA Thermo Scientific, Rockford, USA Grant, England Eco Holten AS, Danmark EG&G, Tyskland Nikon, Japan RCT basic, IKA Labortechnic, Tyskland Thermo Scientific, Rockford, USA Integra Bioscience, Zizers, Sveits Corning Incorporated, Mexico 14
29 Pipettespiser Rør til å sette i luminometer Sterilfilter 0,2 μm Sonikator Vannbad (Sub Aqua 12) Vekt Whirlmixer (M52 Minishaker) Sartoorius Biohit, Helsinki, Finland Sarstedt, Tyskland Whatman, Tyskland Sonics & Materials, Inc. Newtown, USA Grant, England Sartorius, Goettingen, Tyskland Tamro Lab. AS, Sverige 2.2 PC12-celler PC12-celler PC12-celler er en klonet nevronal cellelinje som stammer fra et feokromocytom i rottebinyre [40]. Disse cellene uttrykker reseptorer for en del vekstfaktorer inkludert NGF og kan i nærvær av dette differensiere og få nevronliknende egenskaper. Denne effekten er reversibel ved fjerning av NGF [41] Splitting av PC12-celler til flaske PC12-celler splittes normalt 2 ganger i uken, når de har nådd konfluent tetthet. LAF-benk brukes for å utføre splitting. Cellene dyrkes i celleflaske med PC12-cellekulturmedium (tabell 2.4) og inkuberes i inkubatorskap ved 37 C og 5% CO 2. Prosedyre for splitting av PC12-celler til flaske: 1. Cellene observeres i lysmikroskop for å se om de har nådd konfluent tetthet. 2. Cellemedium helles av flasken ml nytt PC12-cellemedium tilsettes cellene. For at cellene skal løsnes fra flaskebunnen og blandes med mediumet, bankes det godt på sidene og i bunnen av celleflasken. En annen måte som gjør det lettere å løsne cellene er å tilsette 5 ml nytt PC12-medium først og tilsette resten av mediumet som er 5ml etter at cellene har 15
30 løsnet. Slik får man mindre volum av cellesuspensjonen under splitting som gjør prosessen lettere og raskere. 4. Både lang pipette og pasteurpipette kan brukes for å løsne cellene. Det pipetteres først 10 ganger opp og ned med langpipette og deretter 10 ganger ved bruk av pasteurpipette med innskrenket åpning. 5. Cellene inspiseres under lysmikroskop for å se om celleaggregater er løst opp og jevn homogen cellesuspensjon er dannet. Prosessen gjentas ved observasjon av celleaggregater. 6. Hver ny celleflaske tilsettes 1,5 ml cellesuspensjon og 20 ml PC12-medium. Celleflaskene merkes med antall pasasjer Splitting av PC12-celler til skåler Prosedyre for splitting av PC12-celler til skåler: 1. Cellene behandles som beskrevet i trinnene 1-5 over. 2. Celletettheten må bestemmes ved splitting til skåler. Derfor tas en liten mengde cellesuspensjon og settes på et haemocytometer (Neubauer). 3. Lysmikroskop brukes for telling av antall celler i 2 ruter. Gjennomsnittet av antall celler i rutene registreres som antall celler talt. Ønsket celletetthet for både små og medium skåler er 7,0 x 10 4 celler/ml. 4. Mengde cellesuspensjon som trenges til skålene overføres til en kolbe og blandes med nytt PC12 medium: Formlene under anvendes for å beregne mengde cellesuspensjon og mengde PC12-medium: Mengde cellesuspensjon (ml) = antall ml per skål x antall skåler x (7,0x10 4 celler/ml) antall celler i tellekammer x
31 Mengde PC12-medium (ml) = (antall ml/skål x antall skåler) - antall ml cellesuspensjon 5. PC12-medium og cellesuspensjonen blandes godt for å få en jevn fordeling av cellene. Deretter overføres passende mengde til sterile cellekultur skåler. Mengde cellesuspensjon som trenges for skåler med forskjellige størrelser er angitt i tabell 2.6. Små skåler (8,8 cm 2 ) ble brukt til dyrkning av celler til promotorstudier og morfometrisk analyse og medium skåler (21,5 cm 2 ) ble brukt til deteksjon av celledød. 6. Dyrkede celler inkuberes ved 37 C og 5 % CO2. PC12-medium med serum Alle komponentene som står i tabell 2.4 blandes i en 500 ml Dulbecco s Modified Eagle s medium (DMEM) flaske. Natriumpyruvat (P) og Penicillin-Streptomycin (PS) sterilfiltreres ved hjelp av sprøyte og sterilfilter, før blanding. Arbeidet utføres i LAF-benk. Mediet oppbevares ved 2-8 C. Tabell 2.4 Oversikt over komponenter brukt for produksjon av PC12-medium med serum Komponent Mengde Konsentrasjon Dulbecco s Modified Eagle s medium (DMEM) 500 ml Føtalt kalveserum (FBS) 50 ml 10% Hesteserum (HS) 25 ml 5% Glutamin 10 ml 2 mm Natriumpyruvat (P) 5 ml 1 mm Penicillin-Streptomycin (PS) ( U/ml-10 mg/ml ) 5 ml 100 U/ml-0,1 mg/ml PC12 medium uten serum produseres uten FBS og HS 17
32 Tabell 2.5 Oversikt over serum brukt for produksjon av PC12-medium Serum Lot-nr Katalognr. Leverandør Hesteserum Gibco, USA Føtalt kalveserum OSB017 DE14-801F Lonza, USA Tabell 2.6 Mengde cellesuspensjon per skål Skål Størrelse Mengde (ml) Små 8,8 cm 2 1 ml Medium 21,5 cm 2 3,3 ml Eksponering av PC12-celler PC12-celler ble eksponert for silika nanopartikler på to størrelser, 10 nm (Si10) og 50 nm (Si50) og ulike faktorer (tabell ). Eksponering av PC12-celler til promotoraktivitetstudier ble utført 24 timer etter transfeksjon av cellene som tilsvarer 48 timer etter utplating. PC12-celler brukt til telling av celledød og morfometrisk analyse ble eksponert 24 timer etter utplating. Serumfritt PC12-medium ble brukt sammen med silika nanopartikler og som kontroll i ueksponerte skåler (tabell 2.4). For å få en jevn silika nanopartikkel suspensjon før eksponering ble suspensjonen rystet godt ved bruk av wirlmikser. Eksponeringsløsningene ble tilsatt forsiktig på kanten av skålene for å ikke skylle vekk cellene. Eksponering ble gjort i LAF-benk og eksponerte celler sto i inkubatorskap ved 37 C og 5 % CO2. Inkubasjonstid for eksponerte skåler til promotoraktivitet studier var 24 timer og for eksponerte skåler til deteksjon av celledød 48 timer. Eksponerte skåler til morfometrisk analyse ble inkubert i 24, 48 og 72 timer. Preparering av silika nanopartikkelløsning Si10 og Si50 nanopartikkel stamløsninger benyttet i denne masteroppgaven ble levert av leverandøren i konsentrasjon 25 mg/ml. 920 μl stamløsning ble fortynnet med 9,08 ml sterilt 18
33 H 2 O til konsentrasjon 2,3 mg/ml. Denne fortynningen og rekkefølgen i prepareringsprosedyren er optimalisert av [42]. Sluttkonsentrasjon av SiNP var 2 mg/ml med 0,15 % BSA og 1 X PBS. Prosedyre for preparering av nanopartikkelløsning : 1. 1 ml fortynnet nanopartikkelløsning (2,3 mg/ml) sonikeres i sonikator med 420 kj/ml og amplitude ,5 μl BSA (50 mg/ml) tilsettes partikkelløsningen. 3. Deretter tilsettes 115 μl 10 X PBS. 4. Partikkelløsning blandes i wirlmikser. Tabell 2.7 Oversikt over eksponeringer av PC12-celler til promotoraktivitet studier Medium PC12 medium med serum Eksponering av PC12-celler Ueksponert Ueksponert + NAC (250 μm) Ueksponert + Raloxifene (0,1 μm) PC12 medium uten serum Ueksponert Ueksponert + NAC (250 μm) Ueksponert + Raloxifene (0,1 μm) 17beta-østradiol (10 μm ) 17beta-østradiol (10 μm ) + Raloxifene (0,1 μm) Si10 (12,5 μg/ml ) Si10 (25 μg/ml) Si10 (50 μg/ml ) Si10 (25 μg/ml) + NAC (250 μm) Si10 (25 μg/ml) + Raloxifene (0,1 μm) Si50 (25 μg/ml) Si50 (50 μg/ml) Si50 (100 μg/ml) 19
34 Si50 (50 μg/ml) + NAC (250 μm) Si50 (50 μg/ml) + Raloxifene (0,1 μm) Tabell 2.8 Oversikt over eksponeringer av PC12-celler til morfometrisk analyse Medium PC12 medium med serum PC12 medium uten serum Eksponering av PC12-celler Ueksponert Ueksponert + NGF Ueksponert Ueksponert + NGF Si10 (25 μg/ml) Si50 (50 μg/ml) Tabell 2.9 Oversikt over eksponeringer av PC12-celler til deteksjon av celledød Medium PC12 medium med serum PC12 medium uten serum Ekospnering av PC12-celler Ueksponert Ueksponert + NAC (250 μm) Ueksponert + SB (20 μm) Ueksponert + Raloxifene (0,1 μm) Ueksponert + z-devd-cmk (1 μm) Ueksponert + Vitamin C (2 μm) og Vitamin E (10 μm) Ueksponert Ueksponert + NAC (250 μm) Ueksponert + SB (20 μm) Ueksponert + Raloxifene (0,1 μm) Ueksponert + z-devd-cmk (1 μm) Ueksponert + Vitamin C (2 μm) og Vitamin E (10 μm) Si10 (12,5 μg/ml) Si10 (25 μg/ml) Si10 (50 μg/ml) Si10 (100 μg/ml) 20
35 Si10 (25 μg/ml) + NAC (250 μm) Si10 (25 μg/ml) + SB (20 μm) Si10 (25 μg/ml) + Raloxifene (0,1 μm) Si10 (25 μg/ml) + z-devd-cmk (1 μm) Si10 (25 μg/ml) + Vitamin C (2 μm) og Vitamin E (10 μm) Si10 (100 μg/ml) + NAC (250 μm) Si10 (100 μg/ml) + SB (20 μm) Si10 (100 μg/ml) + Raloxifene (0,1 μm) Si50 (25 μg/ml) Si50 (50 μg/ml) Si50 (100 μg/ml) Si50 (200 μg/ml) Si50 (50 μg/ml) + NAC (250 μm) Si50 (100 μg/ml) + NAC (250 μm) Si50 (100 μg/ml) + SB (20 μm) Si50 (100 μg/ml) + Raloxifene (0,1 μm) Si50 (100 μg/ml) + z-devd-cmk (1 μm) Si50 (100 μg/ml) + Vitamin C (2 μm) og Vitamin E (10 μm) 2.3 Transfeksjon For transfeksjon innkapsles DNA i liposom og tilsettes cellene. Liposomet setter seg på cellemembraner og smelter sammen med dem. Dette fører til levering av genetisk materiale til cytoplasma og deretter translokeres til kjernen og kan bli uttrykt (figur 2.1). I dette studiet ble Metafectene Pro brukt som transfeksjonreagens. Metafectene Pro er liposomer som består av kationiske og nøytrale lipider [43]. 21
36 Figur 2.1 Transfeksjon. DNA innkapsles i liposom og tilsettes cellene. Liposomet setter seg på cellemembraner og smelter sammen med dem. Genetisk materiale leveres til cytoplasma og deretter translokeres til kjernen og blir uttrykt. Figur er hentet fra [44] Transfeksjon av PC12-celler Transfeksjon av PC12-celler med Metafectene Pro ble gjennomført 24 timer etter utplating av celler i små skåler. Hver skål tilsettes 1,1 μg DNA hvorav 0,1 μg er internstandarden renillaluciferase under kontroll av thymidinkinasepromotor (rl-tk), og 2 μl Metafectene Pro. Prosessen ble gjennomført i LAF-benk. Skålene sto i inkubatorskap ved 37 C og 5 % CO2 etter transfeksjon. Prosedyre for transfeksjon av PC12-celler med Metafectene Pro: 22
37 1. Mengde serumfritt PC12-medium og DNA som trenges for alle skålene beregnes med formlene under. Mengde serumfritt PC12-medium = 100 μl serumfritt PC12-medium x antall skåler Mengde ildflueluciferase plasmid = 1 μg ildflueluciferase x antall skåler Mengde rl-tk = 0,1 μg rl-tk x antall skåler 2. Halvparten av mediumet tilsettes forsiktig Metafectene Pro. Blandingen beveges forsiktig opp og ned 2 ganger. 3. Halvparten av mediumet tilsettes DNA og blandes godt ved bruk av wirlmikser. Mengde DNA som trenges beregnes med formlene under. 4. Metafectene Pro løsning og DNA løsning blandes forsiktig sammen ved å bevege blandingen opp og ned forsiktig 1 gang. 5. Løsningen inkuberes i 20 minutter ved romtemperatur. 6. Overfører 100 μl løsning til hver skål med medium. For å få jevn fordeling av løsning i skålene dryppes forsiktig 50 μl løsning til alle skålene først og deretter gjentas prosessen med 50 μl løsning. 7. Cellene legges til inkubasjon i inkubatorskap ved 37 C og 5 % CO2. 8. Transfeksjonsløsning erstattes med PC12-medium med serum etter 5 timer. 23
38 Tabell 2.10 Plasmider brukt for transfeksjon av PC12-celler Plasmid Ildflueluciferase under kontroll av GCS-promotor Plasmidets forkortelse Konsentrasjon per skål Leverandør GCS-luc 1 μg / ml Gave fra R.Blomhoff, UiO Ildflueluciferase under kontroll av P53-response element P53 1 μg /ml Addgene Ildflueluciferase under kontroll av MMP9-promotor MMP9-luc 1 μg /ml Gave fra Mikola - UIT Ildflueluciferase under kontroll av Gli-response element Gli-luc 1 μg /ml Riken, Japan Ildflueluciferase under kontroll av østrogen response element ERE-luc 0,5 μg /ml Gave fra Milbrandt, Washington Universitet, St. Louis, USA Østrogen reseptor ER 0,5 μg /ml Gave fra Milbrandt, Washington Universitet, St. Louis, USA Renillaluciferase under kontroll av thymidinkinasepromotor rl-tk 0,1 μg /ml Promega Corporation, USA 24
39 2.4 Luciferase reporter assay Ildflueluciferase Ildflueluciferase er et enzym som katalyserer oksidasjon av luciferin i nærvær av ATP, Mg 2 og O 2. Reaksjonen resulterer i emisjon av fotoner som detekteres i form av lysproduksjon. Lysintensiteten måles i et luminometer. I luciferase reporter assay er reporter gen for ildflueluciferase koblet til promotor som man ønsker å studere aktiviteten på. Graden av reporter genuttrykket som måles i form av lysintensitet, har en direkte sammenheng med promotoraktiviteten [45]. Metoden gir mulighet for å studere aktiviteten av ulike promotorer i ulike celletyper in vitro Høsting av celler til luciferasemåling Transfekterte og eksponerte celler høstes 24 timer etter eksponering som tilsvarer 48 timer etter transfeksjon. For høsting av celler er det behov for 0,1% LUC med 1mM DTT (tabell 2.11 ) og PBS (tabell 2.12). Løsningene må settes på is. Prosedyre for høsting av PC12-celler til luciferasemåling : 1. Vekstmediet i skålene tømmes av. 2. Skålene vaskes med 1 ml PBS x 2. PBS rester fjernes med pipette etter andre vask. 3. En løsning av 0,1% LUC med 1 mm DTT lages.135 μl av løsningen tilsettes midt på skålen. 4. Skålene settes til inkubasjon ved romtemperatur i 5 minutter. 5. Cellene skrapes med en celleskraper i to retninger vinkelrett på hverandre. Celleskraper vaskes med PBS etter skraping av hver skål for å unngå påvirkning mellom skålene. 6. Cellesuspensjonen overføres til eppendorfrør. 7. Eppendorfrørene sentrifugeres i en kjølesentrifuge ved 4 C og rpm (revolutions per minute) x g i 5 minutter. 25
40 Tabell 2.11 Oversikt over innholdsstoffer for løsninger brukt ved høsting av celler til luciferasemåling Løsning Innholdsstoffer Mengde 0,1% LUC 1 M Tris 1 M MES ph 7,8 Triton X-100 MQ-vann (romtemperert) 5 ml 0,1 ml 95 ml Oppbevares ved 2-8 C 1 M DTT Dithiothreitol MQ-vann 154,4 mg 1,0 ml Oppbevares ved -20 C Tabell 2.12 PBS løsning Løsning Innholdsstoffer Mendge Sluttkonsentrasjon PBS (fosfatbufret saltvann) NaCl KCl KH2PO4 NaHPO4x2H2O MQ-vann 32,0 g 0,8 g 1,08 g 7,12 g ad 4000 ml 138 mm 2,7 mm 2 mm 10 mm Justeres til ph 7,4 med HCl/NaOH og autoklaveres Oppbevares kjølig Måling av ildflueluciferaseaktivitet For måling av ildflueluciferaseaktivitet er det behov for 1mM luciferin (tabell 2.13) og LUCcoctail (tabell 2.14 ). Løsningene må settes på is. Prosedyre for måling av ildflueluciferaseaktivitet: 1. Supernatanten overføres til nye eppendorfrør. 26
41 2. 50 μl av cellesuspensjonen blandes med 150 μl LUC-coctail. Spesielle rør som kan settes i luminometeret brukes. 3. Blandingen settes i luminometer som måler lysintensiteten på 2 sekunder. Luciferinløsningen overføres til blandingen via en dispenser. Tabell mM luciferin løsning brukt ved måling av ildflueluciferaseaktivitet Løsning Innholdsstoffer Mengde 1mM luciferin D-luciferin MQ-vann 11 mg 36,7 ml Beskyttes mot lys ved bruk av folie Oppbevares i kjøleskap Tabell 2.14 LUC-coctail løsning brukt ved måling av ildflueluciferaseaktivitet Løsning Innholdsstoffer Mengde LUC-coctail Adenosin 5-trifosfat (ATP) Magnesiumacetat 1 M 1 M Tris 1M MES ph 7,8 MQ-vann 240 mg 2 ml 8,3 ml 89,7 ml Oppbevares ved -20 C Renillaluciferase For korrigering av variasjonen som kan skje i de ulike cellebehandlingstrinnene, er det viktig å bruke internstandard. I denne masteroppgaven er renillaluciferase under kontroll av thymidinkinasepromotor (rl-tk) benyttet som intern standard i luciferase reporter assay metoden. Reportergen for enzymet renillaluciferase er koblet til en konstitutiv thymidinkinasepromotor som ikke lar seg forstyrres av variasjon som følge av behandling av cellene. Antall celler og transfeksjonseffektivitet er eksempler på parametere som kan variere [45]. Enzymet er en katalysator av coelenterazine (renilla luciferin) oksidasjon som utføres i 27
42 nærvær av O 2 og resulterer i lysproduksjon [46]. Lysintensitet detekteres av luminometer og benyttes som mål for promotoraktivitet Måling av renillaluciferaseaktivitet Prosedyre for måling av renillaluciferaseaktivitet : 1. 9,9 ml MQ-vann blandes med 100 μl Stop&Glo -reagens (tabell 2.15). Wirlmikser benyttes for blanding og blandingen beskyttes mot lys ved bruk av folie. 2. Luminometer innstilles på programmet til måling av renillaluciferaseaktivitet. Lysintensiteten måles på 10 sekunder μl fortynnet Stop&Glo -reagens blandes med supernatant fra cellehøsting. Spesielle rør til å sette i luminometer benyttes. Passende volum for testing av renillaluciferaseaktivitet bestemmes ved å finne volumet som ligger i det lineære området. Dette gjøres ved å teste for eksempel 2,5 μl, 5,0 μl og 7,5 μl supernatant fra cellehøstingen og se etter dobling av renillaluciferaseaktivitet ved dobling av volum. 4. Hver prøve måles flere ganger til man finner minst tre verdier med maksimum 10% forskjell. Gjennomsnittet av disse verdiene benyttes som resultat. Tabell 2.15 Stop&Glo -reagens benyttet til måling av renillaluciferaseaktivitet Løsning Innholdsstoffer Mengde Stop&Glo -reagens Stop&Glo -substrat Stop&Glo -buffer 20 μl 1 ml Oppbevares ved -20 C 28
43 2.5 Morfometrisk analyse i mikroskop PC12-celler ble dyrket i små skåler (8,8 cm 2 ) og inkubert ved 37 C og 5 % CO2 i 24 timer. Celler ble eksponert for Si10 og Si50 nanopartikler. Serumfritt PC12-medium ble benyttet for eksponerte celler og ubehandlede kontrollceller. PC12-medium med serum ble brukt som kontroll for effekten av serumdeprivasjon. Cellenes utseende ble studert under Nikon lysmikroskop og bilder fra et passende synsfelt ble tatt med et kamera (Spot) som er koblet til mikroskopet. Prosessen ble gjennomført i tre påfølgende dager for et forsøk og på dag 1, 3 og 6 for et annet forsøk. Bildene ble i etterkant kvalitativt analysert. 29

44 2.6 Deteksjon av celledød Celledød detekteres via en enkel metode som farger døde celler med trypanblåløsning. Metoden gjør det mulig å observere og telle døde og levende celler ved bruk av lysmikroskop. Døde celler har penetrerbar cellemembran som gjør at trypanblåløsning kan penetrere gjennom cellemembran og resultere i blåfargede celler. Levende celler har intakt cellemembran som hindrer penetrasjon av trypanblåløsning. Disse cellene forblir derfor ufargede og kan telles (figur 2.2). Figur 2.2. Deteksjon av celledød i PC12-celler. Døde celler har penetrerbar membran som gjør at trypanblåløsning kan penetrere gjennom cellemembran og farge cellene blåt. Intakt membran hos levende celler hindrer penetrasjon av trypanblåløsningen og cellene forblir ufarget. Svart piler viser døde celler. Hvit piler viser levende celler. Bildet viser døde og levende ueksponerte kontrollceller. Celler dyrkes i medium skåler (21,5 cm 2 ) og eksponeres etter 24 timer inkubasjon i inkubatorskap ved 37 C og 5 % CO2. Deteksjon av celledød gjennomføres 48 timer etter eksponering. Lysmikroskop med 40 ganger forstørrelse brukes. 30
45 2.6.1 Prosedyre for farging med trypanblå 1. 2% trypanblå stockløsning og 1,8% NaCl stockløsning blandes i forhold 1:1 (tabell 2.16). 250 μl trypanblåløsning trenges for hver skål (medium skåler). Nødvendig mengde løsning bestemmes utifra dette. 2. Volumet i skålene (3,3 ml) justeres til 1 ml μl trypanblåløsning overføres hver skål og skålen beveges forsiktig for å fordele løsningen over hele skålen. 4. Skålene inkuberes i inkubatorskap ved 37 C og 5 % CO2 i 30 minutter. 5. Trypanblåløsning fjernes helt av skålene. Antall døde celler (blå farget celler) og antall levende celler (ufargede celler) telles under lysmikroskop. Celler i 3 tilfeldig valgte soner i hver skål telles. Andel døde celler av total antall celler beregnes og resultatet gis i prosent. Tabell 2.16 Løsninger benyttet til deteksjon av celledød Løsning Komponent Mengde 2% trypanblå stockløsning Trypanblå MQ-vann 1 g 50 ml Komponentene blandes og varmes opp til kokepunktet under avtrekk. Blandingen avkjøles i romtemperatur og filtreres. Oppbevares i romtemperatur 1,8% NaCl Oppbevares i romtemperatur NaCl MQ-vann 1,8 g 100 ml 31
46 2.7 Statistiske analyser Figurer i resultatet ble tegnet i Microsoft Exel for Mac Resultat er presentert som gjennomsnitt av flere uavhengige forsøk med standardfeil (SE). Programmet Sigmaplot 13.0 ble benyttet for statistiske analyser. For sammenligning av flere normalfordelte datasett ble One Way Analysis of Variance (ANOVA) benyttet. For sammenligning av flere ikke normalfordelte datasett ble Kruskal-Wallis ANOVA on Ranks benyttet. T-test ble benyttet for parvis sammenligning av normalfordelte datasett, og Mann- Whitney Rank Sum Test ble benyttet for parvis sammenligning av ikke normalfordelte datasett. P-verdi på < 0.05 ble vurdert som signifikant og er merket på figurene. 32
 for å se cellenes respondering til NGF og nevrittutvekst. Cellene responderte til NGF og utviklet lange nevritter (figur 3.1 A).")
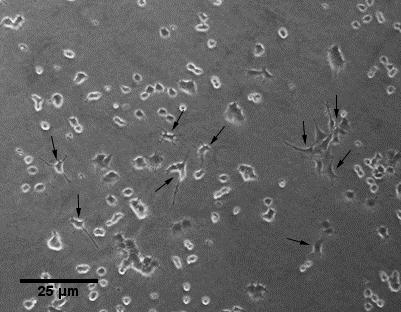
47 3 Resultater 3.1 Morfometrisk analyse Kvalitativ morfometrisk analyse ble gjennomført for å se eventuelle endringer i PC12- cellenes morfologi ved eksponering med SiNP. PC12-cellene ble først eksponert for NGF (Nervevekstfaktor) for å se cellenes respondering til NGF og nevrittutvekst. Cellene responderte til NGF og utviklet lange nevritter (figur 3.1 A). Deretter ble to uavhengige forsøk med NP-er gjennomført. Mikroskopisk analyse av cellene i tre påfølgende dager viste ingen klar forskjell i cellene på dag 1 og 2. Cellene hadde varierende form og en del av dem hadde utviklet nevritter (figur 3.1 B-G). Men derimot på dag 3 var det en forskjell i cellene eksponert for både Si10 og Si50 NP-er sammenlignet med kontrollceller (figur 3.1 H-J). I cellene eksponert for Si10 og Si50 NP-er hadde en del av cellene fått avlang, trådlignende form (figur 3.1 I,J). Ingen av kontrollcellene hadde utviklet denne celle formen på dag 3 (figur 3.1 H). (A) PC12-celler eksponert for NGF 33


48 Dag1 (B) Kontroll (C) Si10 40
")

49 (D) Si50 Dag 2 (E) Kontroll 41
 Si50")
50 (F) Si10 (G) Si50 42


51 Dag 3 (H) Kontroll (I) Si10 43
 (C, F, I) og Si50 (50 mg/ml) (D, G, J) og NGF.")
52 (J) Si50 Figur 3.1. Si10 og Si50 NP-er fører til utvikling av avlange trådlignende PC12-celler. PC12-celler ble dyrket og inkubert. Etter 24 timer ble cellene eksponert med følgende NP konsentrasjoner: Si10 (25 mg/ml) (C, F, I) og Si50 (50 mg/ml) (D, G, J) og NGF. Cellen ble morfometrisk analysert under mikroskop, 24, 48 og 72 timer etter eksponering. (A) PC12-celler på dag 3 etter eksponering for NGF. Kontroll er ueksponerte celler. To uavhengige forsøk ble gjennomført. Tre bilder fra 3 synsfelt i kontroll, Si10 og Si50 NP eksponerte celler ble tatt på dag1, 2 og 3. Tre representative bilder fra hver dag er vist her. Svarte piler viser vanlige celler. Hvite piler viser avlange celler som ses i Si10 og Si50 eksponerte PC12 celler 72 timer etter eksponering (I, J). 3.2 Celledød Celledød ved bruk av batch 1 nanopartikler Endepunktet celledød ble valgt for å studere i hvilken grad eksponering med Si10 og Si50 NP ville påvirke cellenes overlevelse og om hvilke konsentrasjon som ville gi levende celler under promotoraktivitet studier. Effekt av følgende Si10 NP konsentrasjoner: 12,5 mg/ml, 25 44
Sikkerhetsfarmakologiske studier av SSRI-er mulige effekter på nevronkulturer
 Sikkerhetsfarmakologiske studier av SSRI-er mulige effekter på nevronkulturer Marte Rodum Masteroppgave for graden Master i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Sikkerhetsfarmakologiske studier av SSRI-er mulige effekter på nevronkulturer Marte Rodum Masteroppgave for graden Master i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK. EKSAMEN I FAG CELLEBIOLOGI 1 august 1997 Tid: kl
 NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK Faglig kontakt under eksamen: Navn: Professor Tore Lindmo Tlf.:93432 EKSAMEN I FAG 74618 CELLEBIOLOGI 1 august 1997 Tid: kl
NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK Faglig kontakt under eksamen: Navn: Professor Tore Lindmo Tlf.:93432 EKSAMEN I FAG 74618 CELLEBIOLOGI 1 august 1997 Tid: kl
Sikkerhetsfarmakologiske studier av deksametason - mulige effekter på nevronkulturer
 Sikkerhetsfarmakologiske studier av deksametason - mulige effekter på nevronkulturer Karina Borgund Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Sikkerhetsfarmakologiske studier av deksametason - mulige effekter på nevronkulturer Karina Borgund Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
17α-østradiol nedregulerer γ-glutamylcystein syntetase (γ-gcs) promotor i serumdepriverte PC-12 celler
 promotor i serumdepriverte PC-12 celler") 17α-østradiol nedregulerer γ-glutamylcystein syntetase (γ-gcs) promotor i serumdepriverte PC-12 celler Gunn Rebecca Øverby Masteroppgave Biologisk institutt UNIVERSITETET I OSLO Juni 2008 2 3 Forord Arbeidet
17α-østradiol nedregulerer γ-glutamylcystein syntetase (γ-gcs) promotor i serumdepriverte PC-12 celler Gunn Rebecca Øverby Masteroppgave Biologisk institutt UNIVERSITETET I OSLO Juni 2008 2 3 Forord Arbeidet
Toksiske effekter av metaller og selen i storvokstørret fra Mjøsa og Losna
 1 Toksiske effekter av metaller og selen i storvokstørret fra Mjøsa og Losna Eugen Gravningen Sørmo, Forsker, Dr scient Norges teknisk-naturvitenskapelige universitet (NTNU) Institutt for biologi, Realfagsbygget,
1 Toksiske effekter av metaller og selen i storvokstørret fra Mjøsa og Losna Eugen Gravningen Sørmo, Forsker, Dr scient Norges teknisk-naturvitenskapelige universitet (NTNU) Institutt for biologi, Realfagsbygget,
Laboratorieprotokoll for manuell rensing av DNA fra 0,5 ml prøve
 Laboratorieprotokoll for manuell rensing av DNA fra 0,5 ml prøve For rensing av genomisk DNA fra innsamlingssett i seriene Oragene og ORAcollect. Du finner flere språk og protokoller på vårt nettsted,
Laboratorieprotokoll for manuell rensing av DNA fra 0,5 ml prøve For rensing av genomisk DNA fra innsamlingssett i seriene Oragene og ORAcollect. Du finner flere språk og protokoller på vårt nettsted,
Kapittel 20, introduksjon
 Kapittel 20, introduksjon Ekstracellulær signalisering Syntese Frigjøring Transport Forandring av cellulær metabolisme, funksjon, utvikling (trigga av reseptor-signal komplekset) Fjerning av signalet Signalisering
Kapittel 20, introduksjon Ekstracellulær signalisering Syntese Frigjøring Transport Forandring av cellulær metabolisme, funksjon, utvikling (trigga av reseptor-signal komplekset) Fjerning av signalet Signalisering
LEHNINGER PRINCIPLES OF BIOCHEMISTRY
 David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 19 Oxidative Phosphorylation 2008 W. H. Freeman and Company Cellulær respirasjon: siste trinn Elektronoverføring
David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 19 Oxidative Phosphorylation 2008 W. H. Freeman and Company Cellulær respirasjon: siste trinn Elektronoverføring
Sikkerhetsfarmakologiske studier av lamotrigin og valproat mekanismestudier i nevronkulturer
 Sikkerhetsfarmakologiske studier av lamotrigin og valproat mekanismestudier i nevronkulturer Kjersti Jenssen Daling Masteroppgave i farmasi Farmasøytisk institutt Avdeling for farmasøytisk biovitenskap
Sikkerhetsfarmakologiske studier av lamotrigin og valproat mekanismestudier i nevronkulturer Kjersti Jenssen Daling Masteroppgave i farmasi Farmasøytisk institutt Avdeling for farmasøytisk biovitenskap
Caspase-3-avhengig celledød i PC12-celler og kyllingkornceller
 Caspase-3-avhengig celledød i PC12-celler og kyllingkornceller Markus Henriksen Bjørdal Avdeling for farmasøytisk biovitenskap Farmasøytisk Institutt Det matematisk-naturvitenskapelige fakultet UNIVERSITETET
Caspase-3-avhengig celledød i PC12-celler og kyllingkornceller Markus Henriksen Bjørdal Avdeling for farmasøytisk biovitenskap Farmasøytisk Institutt Det matematisk-naturvitenskapelige fakultet UNIVERSITETET
Nytte av prokalsitonin og nøytrofil CD64 som markør for postoperativ infeksjon
 Nytte av prokalsitonin og nøytrofil CD64 som markør for postoperativ infeksjon Kurs i hematologi 18. 19. mai 2015 KS møtesenter, Oslo Kristin Husby, Spesialbioingeniør Prosjektet er utført ved Tverrfaglig
Nytte av prokalsitonin og nøytrofil CD64 som markør for postoperativ infeksjon Kurs i hematologi 18. 19. mai 2015 KS møtesenter, Oslo Kristin Husby, Spesialbioingeniør Prosjektet er utført ved Tverrfaglig
Hensikten med forsøket er å isolere eget DNA fra kinnceller, se hvordan det ser ut og hva det kan brukes til videre.
 DNA HALSKJEDE Hensikt Hensikten med forsøket er å isolere eget DNA fra kinnceller, se hvordan det ser ut og hva det kan brukes til videre. Bakgrunn Det humane genomet består av omtrent 2.9 milliarder basepar.
DNA HALSKJEDE Hensikt Hensikten med forsøket er å isolere eget DNA fra kinnceller, se hvordan det ser ut og hva det kan brukes til videre. Bakgrunn Det humane genomet består av omtrent 2.9 milliarder basepar.
Forelesninger i BI Cellebiologi. Enzymer : senker aktiveringsenergien. Figure 6.13
 Enzymer : senker aktiveringsenergien Figure 6.13 Aktive seter : camp-avhengig protein kinase *For å illustrere hvordan det aktive setet binder et spesifikt substrat er valgt som eksempel camp-avhengig
Enzymer : senker aktiveringsenergien Figure 6.13 Aktive seter : camp-avhengig protein kinase *For å illustrere hvordan det aktive setet binder et spesifikt substrat er valgt som eksempel camp-avhengig
Transport av ioner og næringsstoffer over cellemembraner. Læringsmål IA: Beskrive ulike mekanismer for transport over membraner
 Transport av ioner og næringsstoffer over cellemembraner Læringsmål IA: 2.1.2.6 Beskrive ulike mekanismer for transport over membraner Transport Passiv Aktiv Diffusjon Fasilitert diffusjon Ionekanaler
Transport av ioner og næringsstoffer over cellemembraner Læringsmål IA: 2.1.2.6 Beskrive ulike mekanismer for transport over membraner Transport Passiv Aktiv Diffusjon Fasilitert diffusjon Ionekanaler
HOVEDFAGSOPPGAVE FOR GRADEN Candidatus pharmaciae. Celledød, aktivering av caspase 3, og NGFI- B-forflytning i cellelinjene PC12 og CV1
 HOVEDFAGSOPPGAVE FOR GRADEN Candidatus pharmaciae Celledød, aktivering av caspase 3, og NGFI- B-forflytning i cellelinjene PC12 og CV1 Gunnar Fløan Rimul Avdeling for farmasøytisk biovitenskap Farmasøytisk
HOVEDFAGSOPPGAVE FOR GRADEN Candidatus pharmaciae Celledød, aktivering av caspase 3, og NGFI- B-forflytning i cellelinjene PC12 og CV1 Gunnar Fløan Rimul Avdeling for farmasøytisk biovitenskap Farmasøytisk
Vekstfaktorers interaksjoner med caspaser i PC12-cellelinjen og kornceller fra rotte
 Vekstfaktorers interaksjoner med caspaser i PC12-cellelinjen og kornceller fra rotte Helle Lindland-Tjønn Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Vekstfaktorers interaksjoner med caspaser i PC12-cellelinjen og kornceller fra rotte Helle Lindland-Tjønn Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskapelige
Kreftforskning.no/myklebost. Eva Wessel Pedersen. Cancer Stem Cell Innovation Centre
 Stam Celler og Kreft Eva Wessel Pedersen Avdeling for Tumorbiologi,, Radium Hospitalet Cancer Stem Cell Innovation Centre Oversikt Stamceller generelt Hvorfor vi forsker på stamceller Kreft-stamceller
Stam Celler og Kreft Eva Wessel Pedersen Avdeling for Tumorbiologi,, Radium Hospitalet Cancer Stem Cell Innovation Centre Oversikt Stamceller generelt Hvorfor vi forsker på stamceller Kreft-stamceller
Amgen Europe B.V. Nplate_EU_DosingCalculator_RMP_v3.0_NO_MAR2019. Nplate (romiplostim) Dosekalkulator
 Dosekalkulator") Amgen Europe B.V. Nplate_EU_DosingCalculator_RMP_v3.0_NO_MAR2019 Nplate (romiplostim) Dosekalkulator Veiledning for håndtering Nplate kan kun rekonstitueres med sterilt vann til injeksjonsvæsker uten konserveringsmidler.
Amgen Europe B.V. Nplate_EU_DosingCalculator_RMP_v3.0_NO_MAR2019 Nplate (romiplostim) Dosekalkulator Veiledning for håndtering Nplate kan kun rekonstitueres med sterilt vann til injeksjonsvæsker uten konserveringsmidler.
Colostrum FAQ. Hyppig stilte spørsmål om LRs Colostrum-produkter
 Colostrum FAQ Hyppig stilte spørsmål om LRs Colostrum-produkter Innholdsfortegnelse I. Hva er colostrum? S 3 II. Hvilket dyr kommer colostrum fra? S 3 III. Hva skjer med kalvene? S 3 IV. Hvorfor er colostrum
Colostrum FAQ Hyppig stilte spørsmål om LRs Colostrum-produkter Innholdsfortegnelse I. Hva er colostrum? S 3 II. Hvilket dyr kommer colostrum fra? S 3 III. Hva skjer med kalvene? S 3 IV. Hvorfor er colostrum
Forelesninger i BI Cellebiologi. Protein struktur og funksjon - Kap. 3
 Forelesninger i BI 212 - Cellebiologi Protein struktur og funksjon - Kap. 3 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio.ntnu.no Tlf. 73 59
Forelesninger i BI 212 - Cellebiologi Protein struktur og funksjon - Kap. 3 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio.ntnu.no Tlf. 73 59
FLERVALGSOPPGAVER - CELLEBIOLOGI
 FLERVALGSOPPGAVER - CELLEBIOLOGI Hvert spørsmål har ett riktig svaralternativ. Cellebiologi 1 Hvilken celleorganell er vanlig i både plante- og dyreceller? A) kloroplast B) cellevegg av cellulose C) mitokondrium
FLERVALGSOPPGAVER - CELLEBIOLOGI Hvert spørsmål har ett riktig svaralternativ. Cellebiologi 1 Hvilken celleorganell er vanlig i både plante- og dyreceller? A) kloroplast B) cellevegg av cellulose C) mitokondrium
Interaksjoner mellom MEK/ERK-signalveien og celledød indusert av caspaseaktivatorene PAC-1 og 1541
 Interaksjoner mellom MEK/ERK-signalveien og celledød indusert av caspaseaktivatorene PAC-1 og 1541 Annine Thomassen Gjesvik Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt
Interaksjoner mellom MEK/ERK-signalveien og celledød indusert av caspaseaktivatorene PAC-1 og 1541 Annine Thomassen Gjesvik Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt
Cellesignalisering II: Reseptor tyrosin kinaser, cytosoliske kinaser
 Cellesignalisering II: Reseptor tyrosin kinaser, cytosoliske kinaser! Introduksjon! Definisjon og klassifisering! Kinasefamilier: Receptor/cytosol! Receptor Tyrosin kinase-mediert signalisering! MAP kinase
Cellesignalisering II: Reseptor tyrosin kinaser, cytosoliske kinaser! Introduksjon! Definisjon og klassifisering! Kinasefamilier: Receptor/cytosol! Receptor Tyrosin kinase-mediert signalisering! MAP kinase
Uke 16 (nb, spm. fra uke 16 og uke 17 overlapper ofte)
") Uke 16 (nb, spm. fra uke 16 og uke 17 overlapper ofte) Høst 07 konte 28. Forklar begrepene celledeling og cellevekst, og sett begrepene i sammenheng med hyperplasi og hypertrofi. Celledeling vil si økning
Uke 16 (nb, spm. fra uke 16 og uke 17 overlapper ofte) Høst 07 konte 28. Forklar begrepene celledeling og cellevekst, og sett begrepene i sammenheng med hyperplasi og hypertrofi. Celledeling vil si økning
Mikroalger til medisin; krefthemmere
 Mikroalger til medisin; krefthemmere Kari Skjånes og Hanne Skomedal Bioforsk Jord og Miljø og Plantehelse Agenda Hvorfor mikroalger som krefthemmere Kreftutvikling Potensiale Hva kan utvikles Hvordan utvikle
Mikroalger til medisin; krefthemmere Kari Skjånes og Hanne Skomedal Bioforsk Jord og Miljø og Plantehelse Agenda Hvorfor mikroalger som krefthemmere Kreftutvikling Potensiale Hva kan utvikles Hvordan utvikle
Så, hvordan lager man nye nerveceller?
 Forskningsnyheter om Huntingtons sykdom. I et lettfattelig språk. Skrevet av forskere. Til det globale HS-fellesskapet. Å omdanne hudceller til hjerneceller: et gjennombrudd innen forskning på Huntingtons
Forskningsnyheter om Huntingtons sykdom. I et lettfattelig språk. Skrevet av forskere. Til det globale HS-fellesskapet. Å omdanne hudceller til hjerneceller: et gjennombrudd innen forskning på Huntingtons
Enzymes make the world go around. Enzymer i dagliglivet
 Enzymes make the world go around Enzymer i dagliglivet Innledning Enzymer er i de fleste tilfellene proteiner som øker reaksjonshastigheten til biologiske prosesser. Derfor blir enzymer ofte kalt biologiske
Enzymes make the world go around Enzymer i dagliglivet Innledning Enzymer er i de fleste tilfellene proteiner som øker reaksjonshastigheten til biologiske prosesser. Derfor blir enzymer ofte kalt biologiske
Sikkerhetsfarmakologiske studier av lamotrigin og valproat - mulige effekter på nevronkulturer
 Sikkerhetsfarmakologiske studier av lamotrigin og valproat - mulige effekter på nevronkulturer Sissel Evensen Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskaplige
Sikkerhetsfarmakologiske studier av lamotrigin og valproat - mulige effekter på nevronkulturer Sissel Evensen Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk institutt Det matematisk-naturvitenskaplige
NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR FYSIKK EKSAMEN I EMNE TFY4260 CELLEBIOLOGI OG CELLULÆR BIOFYSIKK
 Side av 1 av5 NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR FYSIKK EKSAMEN I EMNE TFY4260 CELLEBIOLOGI OG CELLULÆR BIOFYSIKK Faglig kontakt under eksamen: Catharina Davies Tel 73593688 eller
Side av 1 av5 NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR FYSIKK EKSAMEN I EMNE TFY4260 CELLEBIOLOGI OG CELLULÆR BIOFYSIKK Faglig kontakt under eksamen: Catharina Davies Tel 73593688 eller
Oncogenic Mutations Affecting Cell Proliferation
 Oncogenic Mutations Affecting Cell Proliferation Fra RTK til Nucleus (Boka s.1070-74) Normalt kreves et vekst stimulerende signal ( growth factor eks. PDGF, EGF, NGF) for at celler skal gå inn i celledeling,
Oncogenic Mutations Affecting Cell Proliferation Fra RTK til Nucleus (Boka s.1070-74) Normalt kreves et vekst stimulerende signal ( growth factor eks. PDGF, EGF, NGF) for at celler skal gå inn i celledeling,
Norecopas pris til fremme av de 3 R-ene
 Norecopas pris til fremme av de 3 R-ene (Replacement, Reduction, Refinement) for 2013 Et diplom og kr. 30 000 Priskomitéen - Lasse A. Skoglund, akademia - Heidi Bugge, forvaltning - Torill Malmstrøm, dyrevernorganisasjoner
Norecopas pris til fremme av de 3 R-ene (Replacement, Reduction, Refinement) for 2013 Et diplom og kr. 30 000 Priskomitéen - Lasse A. Skoglund, akademia - Heidi Bugge, forvaltning - Torill Malmstrøm, dyrevernorganisasjoner
1 Bakgrunn Metode og gjennomføring Belegg Biofilmdannelse Resultater Biofilmdannelse Diskusjon...
 Innholdsfortegnelse Bakgrunn... 3 2 Metode og gjennomføring... 4 2. Belegg... 4 2.2 Biofilmdannelse... 4 3 Resultater... 5 3. Biofilmdannelse... 5 4 Diskusjon... 6 5 Foreløpige konklusjoner... 7 60205-2
Innholdsfortegnelse Bakgrunn... 3 2 Metode og gjennomføring... 4 2. Belegg... 4 2.2 Biofilmdannelse... 4 3 Resultater... 5 3. Biofilmdannelse... 5 4 Diskusjon... 6 5 Foreløpige konklusjoner... 7 60205-2
Renholdskontroll for forbygging av Listeria 6. MARS 2018
 Renholdskontroll for forbygging av Listeria 6. MARS 2018 Solveig Langsrud, Seniorforsker Nofima Unngå Listeria på produkt fire områder med ulike aktører Internasjonalt anerkjent regime for håndtering av
Renholdskontroll for forbygging av Listeria 6. MARS 2018 Solveig Langsrud, Seniorforsker Nofima Unngå Listeria på produkt fire områder med ulike aktører Internasjonalt anerkjent regime for håndtering av
Cerebral parese og genetikk Har det noe med hverandre å gjøre?
 Cerebral parese og genetikk Har det noe med hverandre å gjøre? Espen Lien Barneklinikken, St. Olavs hospital Institutt for laboratoriemedisin, barne- og kvinnesykdommer NTNU 1. CP som genetisk tilstand?
Cerebral parese og genetikk Har det noe med hverandre å gjøre? Espen Lien Barneklinikken, St. Olavs hospital Institutt for laboratoriemedisin, barne- og kvinnesykdommer NTNU 1. CP som genetisk tilstand?
Reproduksjon av dyrevirus. Adsorpsjon Penetrasjon og avkledning Replikasjon og transkripsjon Syntese og samling (assembly) av viruskapsid Frigjøring
 av viruskapsid Frigjøring") Reproduksjon av dyrevirus Adsorpsjon Penetrasjon og avkledning Replikasjon og transkripsjon Syntese og samling (assembly) av viruskapsid Frigjøring ATTACHMENT Click after each step to view process PENETRATION
Reproduksjon av dyrevirus Adsorpsjon Penetrasjon og avkledning Replikasjon og transkripsjon Syntese og samling (assembly) av viruskapsid Frigjøring ATTACHMENT Click after each step to view process PENETRATION
BIOS 2 Biologi
 . Figurer kapittel 2: Energi Figur s. 48 Solenergi Økosystem CO 2 + 2 O Fotosyntese i kloroplaster Organiske molekyler + O 2 Celleånding i mitokondrier Energi til arbeid Varme rodusentene i økosystemet
. Figurer kapittel 2: Energi Figur s. 48 Solenergi Økosystem CO 2 + 2 O Fotosyntese i kloroplaster Organiske molekyler + O 2 Celleånding i mitokondrier Energi til arbeid Varme rodusentene i økosystemet
Viktige opplysninger: Oppgavesettet utgjør totalt 100 vekttall. Antall vekttall er vist i parentes ved hver spørsmålsgruppe.
 Ordinær eksamen, MEDSEM/ODSEM/ERNSEM2 Vår 2012 Onsdag 20. juni 2012 kl. 09:00-15:00 Oppgavesettet består av 6 sider, inkludert vedlegg Viktige opplysninger: Oppgavesettet utgjør totalt 100 vekttall. Antall
Ordinær eksamen, MEDSEM/ODSEM/ERNSEM2 Vår 2012 Onsdag 20. juni 2012 kl. 09:00-15:00 Oppgavesettet består av 6 sider, inkludert vedlegg Viktige opplysninger: Oppgavesettet utgjør totalt 100 vekttall. Antall
Bioenergetikk og Krebs syklus Oksidativ fosforylering
 Bioenergetikk og Krebs syklus Oksidativ fosforylering Bioenergetikk, IA 2015 Det store bildet Bioenergetikk ATP Den mengden ATP som brytes ned og dannes pr dag hos mennesket, tilsvarer omtrent kroppsvekten
Bioenergetikk og Krebs syklus Oksidativ fosforylering Bioenergetikk, IA 2015 Det store bildet Bioenergetikk ATP Den mengden ATP som brytes ned og dannes pr dag hos mennesket, tilsvarer omtrent kroppsvekten
Introduksjon til nevrotoksikologi
 Introduksjon til nevrotoksikologi Professor Tore Syversen Institutt for nevromedisin Det medisinske fakultet NTNU Tore.syversen@medisin.ntnu.no Aktualitet Metaller: kvikksølv, bly, aluminium, mangan Løsningsmidler
Introduksjon til nevrotoksikologi Professor Tore Syversen Institutt for nevromedisin Det medisinske fakultet NTNU Tore.syversen@medisin.ntnu.no Aktualitet Metaller: kvikksølv, bly, aluminium, mangan Løsningsmidler
Forløp av ikke-adaptiv og adaptiv immunrespons. Mononukleære celler, metylfiolett farging
 Forløp av ikke-adaptiv og adaptiv immunrespons Mononukleære celler, metylfiolett farging 1 Nøytrofile granulocytter Gjenkjennelsesprinsipper medfødt vs. adaptiv immunitet Toll Like Receptors Mikroorganismer
Forløp av ikke-adaptiv og adaptiv immunrespons Mononukleære celler, metylfiolett farging 1 Nøytrofile granulocytter Gjenkjennelsesprinsipper medfødt vs. adaptiv immunitet Toll Like Receptors Mikroorganismer
Obligatorisk oppgave 2 MBV1030 Høst 2005
 Obligatorisk oppgave 2 MBV1030 Høst 2005 Levert av (navn): Første del: Flervalgsspørsmål. Angi det svaralternativet (ett) du mener er korrekt. I-1: Ved anaerob glykolyse dannes det laktat. Dersom glukosen
Obligatorisk oppgave 2 MBV1030 Høst 2005 Levert av (navn): Første del: Flervalgsspørsmål. Angi det svaralternativet (ett) du mener er korrekt. I-1: Ved anaerob glykolyse dannes det laktat. Dersom glukosen
Antioksidanter: mat eller tilskudd?
 Antioksidanter: mat eller tilskudd? Rune Blomhoff Institutt for medisinske basalfag, Universitetet i Oslo, Kreft, kirurgi og transplantasjonsklinikken, Oslo Universitetssykehus Oksygen et tveegget sverd
Antioksidanter: mat eller tilskudd? Rune Blomhoff Institutt for medisinske basalfag, Universitetet i Oslo, Kreft, kirurgi og transplantasjonsklinikken, Oslo Universitetssykehus Oksygen et tveegget sverd
Mekanismer ved dexametason- og glutamatindusert celledød i nervecellekulturer Minh Dung Trinh
 HOVEDFAGSOPPGAVE FOR GRADEN Candidatus pharmaciae Mekanismer ved dexametason- og glutamatindusert celledød i nervecellekulturer Minh Dung Trinh Farmasøytisk institutt Det matematiske-naturvitenskapelige
HOVEDFAGSOPPGAVE FOR GRADEN Candidatus pharmaciae Mekanismer ved dexametason- og glutamatindusert celledød i nervecellekulturer Minh Dung Trinh Farmasøytisk institutt Det matematiske-naturvitenskapelige
In vitro cytotoksisk effekt av venom fra vestlig honningbie (Apis mellifera) hos glioblastoma cellelinjer av typen U251 og C6.
 hos glioblastoma cellelinjer av typen U251 og C6.") In vitro cytotoksisk effekt av venom fra vestlig honningbie (Apis mellifera) hos glioblastoma cellelinjer av typen U251 og C6. av Sivert Andreas Bruvoll Knutsen Masteroppgave i farmasi Juni 2008 Senter
In vitro cytotoksisk effekt av venom fra vestlig honningbie (Apis mellifera) hos glioblastoma cellelinjer av typen U251 og C6. av Sivert Andreas Bruvoll Knutsen Masteroppgave i farmasi Juni 2008 Senter
Side 1 ARBEIDSBESKRIVELSE Institutt for husdyr- og akvakulturvitenskap, NMBU
 1 EIDSBESKRIVELSE Institutt for husdyr- og akvakulturvitenskap, NMBU Metodenavn: Tryptofan BIOVIT-nr: Arb1051 1. Innledning/hensikt Metoden bestemmer totalinnholdet av tryptofan i fôr og faeces, ved basisk
1 EIDSBESKRIVELSE Institutt for husdyr- og akvakulturvitenskap, NMBU Metodenavn: Tryptofan BIOVIT-nr: Arb1051 1. Innledning/hensikt Metoden bestemmer totalinnholdet av tryptofan i fôr og faeces, ved basisk
Membran-proteiner (Del 3.4)
") Membran-proteiner (Del 3.4) Poriner adskiller seg dramatisk fra andre integral proteiner. Finnes bl.a. i ytre membranen hos E.coli (se Figure 1-7). Poriner er med å beskytte bakterien mot toksiske forbindelser
Membran-proteiner (Del 3.4) Poriner adskiller seg dramatisk fra andre integral proteiner. Finnes bl.a. i ytre membranen hos E.coli (se Figure 1-7). Poriner er med å beskytte bakterien mot toksiske forbindelser
Oppgave: MED1100-3_OPPGAVE2_H16_KONT
 Side 10 av 35 Oppgave: MED1100-3_OPPGAVE2_H16_KONT Del 1: Ola har en arvelig betinget kombinert immundefekt med mangel på både T-celler og B-celler. Ola får derfor gjentatte Hvorfor er Ola beskyttet mot
Side 10 av 35 Oppgave: MED1100-3_OPPGAVE2_H16_KONT Del 1: Ola har en arvelig betinget kombinert immundefekt med mangel på både T-celler og B-celler. Ola får derfor gjentatte Hvorfor er Ola beskyttet mot
Celledød og organellefordeling i sentrifugeringsfraksjoner fra kornceller og PC12-celler
 Celledød og organellefordeling i sentrifugeringsfraksjoner fra kornceller og PC12-celler Stine Malme Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk Institutt Det matematisk-naturvitenskapelige
Celledød og organellefordeling i sentrifugeringsfraksjoner fra kornceller og PC12-celler Stine Malme Masteroppgave i farmasi Avdeling for farmasøytisk biovitenskap Farmasøytisk Institutt Det matematisk-naturvitenskapelige
Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu
,Botanisk institutt,ntnu") Forelesninger i BI 212 - Cellebiologi Biomembraner og subcellular organisering av eukaryote celler - Kap. 5 - vår 2002 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-
Forelesninger i BI 212 - Cellebiologi Biomembraner og subcellular organisering av eukaryote celler - Kap. 5 - vår 2002 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-
Grunnleggende cellebiologi
 Grunnleggende cellebiologi Ann Kristin Sjaastad Sert. yrkeshygieniker, Dr. Philos HMS-seksjonen, NTNU Tema Cellens oppbygning Transportmekanismer Arvestoff og proteinsyntese Mutasjoner og genotoksisitet
Grunnleggende cellebiologi Ann Kristin Sjaastad Sert. yrkeshygieniker, Dr. Philos HMS-seksjonen, NTNU Tema Cellens oppbygning Transportmekanismer Arvestoff og proteinsyntese Mutasjoner og genotoksisitet
Grunnleggende om nervesystemet
 Grunnleggende om nervesystemet Nevronal cellebiologi Membranpotensialet Aksjonspotentialet Synapsepotensialet Nevrotransmittere Gliaceller Synaptisk plastisitet Basal medisinsk nevrobiologi Underviser:
Grunnleggende om nervesystemet Nevronal cellebiologi Membranpotensialet Aksjonspotentialet Synapsepotensialet Nevrotransmittere Gliaceller Synaptisk plastisitet Basal medisinsk nevrobiologi Underviser:
Amplifikasjonsteknikker - andre metoder
 Amplifikasjonsteknikker - andre metoder Svein Arne Nordbø TH-28973 17.03.15 Alternative amplifikasjonsmetoder Templat-amplifikasjons metoder Signal-amplifikasjonsmetoder Templat-amplifikasjons metoder
Amplifikasjonsteknikker - andre metoder Svein Arne Nordbø TH-28973 17.03.15 Alternative amplifikasjonsmetoder Templat-amplifikasjons metoder Signal-amplifikasjonsmetoder Templat-amplifikasjons metoder
Bokmål. Skriftlig eksamen MD4011 semester IA/B kull 11
 NTNU Det medisinske fakultet Bokmål Sensurfrist: 12. juni 2012 Skriftlig eksamen MD4011 semester IA/B kull 11 Tirsdag 22. mai 2012 Kl. 09.00 15.00 (16.00) Oppgavesettet er på 27 sider inklusive forsiden
NTNU Det medisinske fakultet Bokmål Sensurfrist: 12. juni 2012 Skriftlig eksamen MD4011 semester IA/B kull 11 Tirsdag 22. mai 2012 Kl. 09.00 15.00 (16.00) Oppgavesettet er på 27 sider inklusive forsiden
Undersøkelse av beskyttelse mot elektromagnetisk stråling med Aires Shield.
 Undersøkelse av beskyttelse mot elektromagnetisk stråling med Aires Shield. Det er gjennomført en rekke undersøkelser med deltakere i alderen 18 til 70 år, som beviste effektiviteten av dette produktet.
Undersøkelse av beskyttelse mot elektromagnetisk stråling med Aires Shield. Det er gjennomført en rekke undersøkelser med deltakere i alderen 18 til 70 år, som beviste effektiviteten av dette produktet.
SPISS. Bakterier under UV-stråler. Naturfaglige artikler av elever i videregående opplæring 56 SPISS. Innledning
 SPISS Naturfaglige artikler av elever i videregående opplæring Forfatter: Jonas Blårud, Kuben videregående skole I dette forsøket undersøkes det om eksponering for Ultrafiolette stråler har en skadelig
SPISS Naturfaglige artikler av elever i videregående opplæring Forfatter: Jonas Blårud, Kuben videregående skole I dette forsøket undersøkes det om eksponering for Ultrafiolette stråler har en skadelig
Preanalyse. Kurs i Molekylærpatologi Oslo, juni 2017
 Preanalyse Kurs i Molekylærpatologi Oslo, 07.-08. juni 2017 Heidemarie Svendsen Bioingeniør Enhet for molekylærpatologi Avdeling for Patologi Oslo Universitetssykehus Innhold Prøvemateriale Preparering
Preanalyse Kurs i Molekylærpatologi Oslo, 07.-08. juni 2017 Heidemarie Svendsen Bioingeniør Enhet for molekylærpatologi Avdeling for Patologi Oslo Universitetssykehus Innhold Prøvemateriale Preparering
Lourdes Hydrogenvann maskin. Vann Gården As
 Lourdes Hydrogenvann maskin Vann Gården As S Hydrogen S S S S Er det minste atomet som finner (1 proton 1 elektron) Komponerer 90% av universet En transportør for elektron (bevegelser mellom elektroner
Lourdes Hydrogenvann maskin Vann Gården As S Hydrogen S S S S Er det minste atomet som finner (1 proton 1 elektron) Komponerer 90% av universet En transportør for elektron (bevegelser mellom elektroner
Reproduksjon av dyrevirus. Adsorpsjon Penetrasjon og avkledning Replikasjon og transkripsjon Syntese og samling (assembly) av viruskapsid Frigjøring
 av viruskapsid Frigjøring") Reproduksjon av dyrevirus Adsorpsjon Penetrasjon og avkledning Replikasjon og transkripsjon Syntese og samling (assembly) av viruskapsid Frigjøring ATTACHMENT Click after each step to view process PENETRATION
Reproduksjon av dyrevirus Adsorpsjon Penetrasjon og avkledning Replikasjon og transkripsjon Syntese og samling (assembly) av viruskapsid Frigjøring ATTACHMENT Click after each step to view process PENETRATION
TRANSPORT GJENNOM CELLEMEMBRANEN
 TRANSPORT GJENNOM CELLEMEMBRANEN MÅL: Forklare transport gjennom cellemembranen ved å bruke kunnskap om passive og aktive transportmekanismer Cellemembranen - funksjon - beskytte innholdet i cellen kontroll
TRANSPORT GJENNOM CELLEMEMBRANEN MÅL: Forklare transport gjennom cellemembranen ved å bruke kunnskap om passive og aktive transportmekanismer Cellemembranen - funksjon - beskytte innholdet i cellen kontroll
Forelesninger i BI Cellebiologi Proteinrensing - Væskekromatografi. Figure 3-43 b
 Proteinrensing - Væskekromatografi Figure 3-43 b Proteinrensing - Væskekromatografi Ved affinitets-kromatografi brukes en søyle med kuler som er dekket med ligander (f.eks. et enzym-substrat eller et annet
Proteinrensing - Væskekromatografi Figure 3-43 b Proteinrensing - Væskekromatografi Ved affinitets-kromatografi brukes en søyle med kuler som er dekket med ligander (f.eks. et enzym-substrat eller et annet
Testing av rengjøring og renhet
 Dekontamineringsdagene 2019 Testing av rengjøring og renhet Egil Lingaas Nasjonal kompetansetjeneste for dekontaminering Rent eller urent? Rengjort eller ikke? Nasjonal kompetansetjeneste for dekontaminering
Dekontamineringsdagene 2019 Testing av rengjøring og renhet Egil Lingaas Nasjonal kompetansetjeneste for dekontaminering Rent eller urent? Rengjort eller ikke? Nasjonal kompetansetjeneste for dekontaminering
Forelesninger i BI Cellebiologi. Denaturering og renaturering. Figure 3-13
 Figure 3.9 Denaturering og renaturering Figure 3-13 Denaturering og renaturering Figure 3-14 Viser tre trinn i refolding av et protein som har vært denaturert. Molten globule -formen er en intermediær
Figure 3.9 Denaturering og renaturering Figure 3-13 Denaturering og renaturering Figure 3-14 Viser tre trinn i refolding av et protein som har vært denaturert. Molten globule -formen er en intermediær
Nr. 46/108 EØS-tillegget til De Europeiske Fellesskaps Tidende KOMMISJONSDIREKTIV 1999/76/EF. av 23. juli 1999
 Nr. 46/108 EØS-tillegget til De Europeiske Fellesskaps Tidende KOMMISJONEN FOR DE EUROPEISKE FELLESSKAP HAR under henvisning til traktaten om opprettelse av Det europeiske fellesskap, under henvisning
Nr. 46/108 EØS-tillegget til De Europeiske Fellesskaps Tidende KOMMISJONEN FOR DE EUROPEISKE FELLESSKAP HAR under henvisning til traktaten om opprettelse av Det europeiske fellesskap, under henvisning
Bruksanvisning IVD Matrix HCCA-portioned
 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
ML-208, generell informasjon
 ML-208, generell informasjon Emnekode: ML-208 Emnenavn: Molekylærbiologi Dato:20.12.2017 Varighet:4 timer Tillatte hjelpemidler: Ingen Merknader:Lag gjerne tegninger og figurer for å illustrere og forklare
ML-208, generell informasjon Emnekode: ML-208 Emnenavn: Molekylærbiologi Dato:20.12.2017 Varighet:4 timer Tillatte hjelpemidler: Ingen Merknader:Lag gjerne tegninger og figurer for å illustrere og forklare
BI 212- Protein Sorting - Kap. 17 Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.)
") Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.) Veiene for opptak fra cytosol av kloroplast-proteiner Opptak av proteiner fra cytosol til kloroplaster ligner mye på mitokondrie-importen
Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.) Veiene for opptak fra cytosol av kloroplast-proteiner Opptak av proteiner fra cytosol til kloroplaster ligner mye på mitokondrie-importen
Glutamat reseptorer og signaltransduksjon. Læringsmål: Ha grundig forståelse for oppbygningen, funksjonen og utbredelsen av glutamat reseptorer.
 Glutamat reseptorer og signaltransduksjon Læringsmål: Ha grundig forståelse for oppbygningen, funksjonen og utbredelsen av glutamat reseptorer. Ha grundig forståelse for prinsippene for signaltransduksjon
Glutamat reseptorer og signaltransduksjon Læringsmål: Ha grundig forståelse for oppbygningen, funksjonen og utbredelsen av glutamat reseptorer. Ha grundig forståelse for prinsippene for signaltransduksjon
Forskning med nanopartikler til bruk innen kreftdiagnose og behandling
 Nano World Cancer Day 31.1.14 An ETP Nanomedicine event across 13 European Countries Forskning med nanopartikler til bruk innen kreftdiagnose og behandling Et nytt nasjonalt kompetansebyggende prosjekt
Nano World Cancer Day 31.1.14 An ETP Nanomedicine event across 13 European Countries Forskning med nanopartikler til bruk innen kreftdiagnose og behandling Et nytt nasjonalt kompetansebyggende prosjekt
4 KONSENTRASJON 4.1 INNLEDNING
 4 KONSENTRASJON 4.1 INNLEDNING 1 Terminologi En løsning er tidligere definert som en homogen blanding av rene stoffer (kap. 1). Vi tenker vanligvis på en løsning som flytende, dvs. at et eller annet stoff
4 KONSENTRASJON 4.1 INNLEDNING 1 Terminologi En løsning er tidligere definert som en homogen blanding av rene stoffer (kap. 1). Vi tenker vanligvis på en løsning som flytende, dvs. at et eller annet stoff
Flervalgsoppgaver: proteinsyntese
 Flervalgsoppgaver - proteinsyntese Hver oppgave har ett riktig svaralternativ. Proteinsyntese 1 Hva blir transkribert fra denne DNA sekvensen: 3'-C-C-G-A-A-T-G-T-C-5'? A) 3'-G-G-C-U-U-A-C-A-G-5' B) 3'-G-G-C-T-T-A-C-A-G-5'
Flervalgsoppgaver - proteinsyntese Hver oppgave har ett riktig svaralternativ. Proteinsyntese 1 Hva blir transkribert fra denne DNA sekvensen: 3'-C-C-G-A-A-T-G-T-C-5'? A) 3'-G-G-C-U-U-A-C-A-G-5' B) 3'-G-G-C-T-T-A-C-A-G-5'
Trening øker gjenvinning i celler Natur og miljø
 Forskningsnyheter om Huntingtons sykdom. I et lettfattelig språk. Skrevet av forskere. Til det globale HS-fellesskapet. Trening øker gjenvinning i celler Trening øker cellulær gjenvinning hos mus. Er det
Forskningsnyheter om Huntingtons sykdom. I et lettfattelig språk. Skrevet av forskere. Til det globale HS-fellesskapet. Trening øker gjenvinning i celler Trening øker cellulær gjenvinning hos mus. Er det
LEKSJON 4: BIOTEKNOLOGI HVORDAN VI BRUKER NATURENS EGNE MEKANISMER TIL VÅR FORDEL, OG UTFORDRINGENE SOM FØLGER MED
 LEKSJON 4: BIOTEKNOLOGI HVORDAN VI BRUKER NATURENS EGNE MEKANISMER TIL VÅR FORDEL, OG UTFORDRINGENE SOM FØLGER MED KOMPETANSEMÅL Forklarebegrepene krysning og genmodifisering, og hvordan bioteknologi brukes
LEKSJON 4: BIOTEKNOLOGI HVORDAN VI BRUKER NATURENS EGNE MEKANISMER TIL VÅR FORDEL, OG UTFORDRINGENE SOM FØLGER MED KOMPETANSEMÅL Forklarebegrepene krysning og genmodifisering, og hvordan bioteknologi brukes
Apoptose/Nekrose mekanismer
 Apoptose/Nekrose mekanismer Post-doc Marit W. Anthonsen, DU1-115 marit.w.anthonsen@chembio.ntnu.no OVERSIKT: * Definisjoner apoptose/nekrose * To hovedsignalveier for apoptose * Caspaser (mordere som medierer
Apoptose/Nekrose mekanismer Post-doc Marit W. Anthonsen, DU1-115 marit.w.anthonsen@chembio.ntnu.no OVERSIKT: * Definisjoner apoptose/nekrose * To hovedsignalveier for apoptose * Caspaser (mordere som medierer
... Proteiner og enzymer. kofaktor. polypeptid
 30 Proteiner og enzymer Proteiner er bygd opp av rekker av aminosyrer som er kveilet sammen ved hjelp av bindinger på kryss og tvers, såkalte peptidbindinger. Slike oppkveilete rekker av aminosyrer kaller
30 Proteiner og enzymer Proteiner er bygd opp av rekker av aminosyrer som er kveilet sammen ved hjelp av bindinger på kryss og tvers, såkalte peptidbindinger. Slike oppkveilete rekker av aminosyrer kaller
Idrett og energiomsetning
 1 Medisin stadium IA, Tonje S. Steigedal 2 ATP er den eneste forbindelsen som kan drive kontraksjon av musklene. ATPnivået i muskelcellene er imidlertid begrenset, og må etterfylles kontinuerlig. Ved ulike
1 Medisin stadium IA, Tonje S. Steigedal 2 ATP er den eneste forbindelsen som kan drive kontraksjon av musklene. ATPnivået i muskelcellene er imidlertid begrenset, og må etterfylles kontinuerlig. Ved ulike
Informasjon om Olivita
 Informasjon om Olivita Les nøye gjennom dette før du begynner å bruke Olivita - Ta vare på dette pakningsvedlegget. Du kan få behov for å lese det igjen. - Hvis du har ytterligere spørsmål, kontakt Olivita
Informasjon om Olivita Les nøye gjennom dette før du begynner å bruke Olivita - Ta vare på dette pakningsvedlegget. Du kan få behov for å lese det igjen. - Hvis du har ytterligere spørsmål, kontakt Olivita
Immunstimulanter for potensiering av torskens naturlige immunsystem
 Store programmer HAVBRUK - En næring i vekst Faktaark www.forskningsradet.no/havbruk Immunstimulanter for potensiering av torskens naturlige immunsystem Prosjekt: Immunostimulation of Atlantic cod (Gadus
Store programmer HAVBRUK - En næring i vekst Faktaark www.forskningsradet.no/havbruk Immunstimulanter for potensiering av torskens naturlige immunsystem Prosjekt: Immunostimulation of Atlantic cod (Gadus
FLERVALGSOPPGAVER - CELLEMEMBRANEN
 FLERVALGSOPPGAVER - CELLEMEMBRANEN Hvert spørsmål har ett riktig svaralternativ. Transport cellemembranen 1 På hvilken måte er ulike membraner i en celle forskjellige? A) Fosfolipider finnes bare i enkelte
FLERVALGSOPPGAVER - CELLEMEMBRANEN Hvert spørsmål har ett riktig svaralternativ. Transport cellemembranen 1 På hvilken måte er ulike membraner i en celle forskjellige? A) Fosfolipider finnes bare i enkelte
ML-208, generell informasjon
 ML-208, generell informasjon Emnekode: ML-208 Emnenavn: Molekylærbiologi Dato:20.12.2017 Varighet:4 timer Tillatte hjelpemidler: Ingen Merknader:Lag gjerne tegninger og figurer for å illustrere og forklare
ML-208, generell informasjon Emnekode: ML-208 Emnenavn: Molekylærbiologi Dato:20.12.2017 Varighet:4 timer Tillatte hjelpemidler: Ingen Merknader:Lag gjerne tegninger og figurer for å illustrere og forklare
1. Medfødt og ervervet immunitet. Karl Schenck, V2015
 1. Medfødt og ervervet immunitet Karl Schenck, V2015 Medfødt og ervervet immunforsvar Antimicrobial peptides «Alltid beredt!» Relativt uspesifikt Må aktiveres Spesifikt Komponenter av medfødt immunitet
1. Medfødt og ervervet immunitet Karl Schenck, V2015 Medfødt og ervervet immunforsvar Antimicrobial peptides «Alltid beredt!» Relativt uspesifikt Må aktiveres Spesifikt Komponenter av medfødt immunitet
Fiksering. Er det viktig?
 Fiksering. Er det viktig? 2. Parafininnstøpt materiale 1 Fiksering 2 Dehydrering 3 Klaring 4 Parafininnfiltrasjon 5 Parafininnstøping FIKSERING ----hvorfor er dette trinnet så viktig? 3. Kjemisk fiksering
Fiksering. Er det viktig? 2. Parafininnstøpt materiale 1 Fiksering 2 Dehydrering 3 Klaring 4 Parafininnfiltrasjon 5 Parafininnstøping FIKSERING ----hvorfor er dette trinnet så viktig? 3. Kjemisk fiksering
Kapittel 16 Utvikling: differensielt genuttrykk
 Kapittel 16 Utvikling: differensielt genuttrykk 1. Utvikling 2. Differensielt genuttrykks rolle i celledifferensiering 3. Polaritets rolle i cellebestemmelse 4. Embryonisk induksjon i cellebestemmelse
Kapittel 16 Utvikling: differensielt genuttrykk 1. Utvikling 2. Differensielt genuttrykks rolle i celledifferensiering 3. Polaritets rolle i cellebestemmelse 4. Embryonisk induksjon i cellebestemmelse
Disposisjon. Noen fakta om hjernen 06.10.2014. KROSS 2014 Kompetanse om Rehabilitering Om Syn og Slag. KFAmthor VVHF 1
 Hjernens plastisitet - konsekvenser for slagrehabilitering Karl-Friedrich Amthor Nevrolgisk avdeling Drammen sykehus Disposisjon Noen (u)nyttige fakta om hjernen Hva skjer ved et hjerneslag Behandling
Hjernens plastisitet - konsekvenser for slagrehabilitering Karl-Friedrich Amthor Nevrolgisk avdeling Drammen sykehus Disposisjon Noen (u)nyttige fakta om hjernen Hva skjer ved et hjerneslag Behandling
Smerte. Arne Tjølsen
 Smerte Arne Tjølsen Om smerte Disposisjon 1) Hva er smerte? 2) Nosiseptiv smerte 3) Modulering og sensitivisering Smerte er en ubehagelig sensorisk og emosjonell opplevelse som følge av faktisk eller potensiell
Smerte Arne Tjølsen Om smerte Disposisjon 1) Hva er smerte? 2) Nosiseptiv smerte 3) Modulering og sensitivisering Smerte er en ubehagelig sensorisk og emosjonell opplevelse som følge av faktisk eller potensiell
På de åpne spørsmålene (26-30) kan det oppnås maksimalt 5 poeng per oppgave.
 kan det oppnås maksimalt 5 poeng per oppgave.") 051HOEM2 2-1 Prøve i anatomi og fysiologi. 18.10.2010 På spørsmål 1-25 skal det markeres med ett kryss ut for det svaralternativet du mener er korrekt. Riktig svar på spørsmål 1-25 gir 1 poeng, feil svar
051HOEM2 2-1 Prøve i anatomi og fysiologi. 18.10.2010 På spørsmål 1-25 skal det markeres med ett kryss ut for det svaralternativet du mener er korrekt. Riktig svar på spørsmål 1-25 gir 1 poeng, feil svar
Lab forelesning. C-vitamin. Enzymer i hverdagen
 Lab forelesning C-vitamin Enzymer i hverdagen C-vitamin eller askorbinsyre Finnes i svært mange frukter og grønnsaker Viktige kilder: appelsin paprika poteter C-vitamin Har mange viktige funksjoner i kroppen
Lab forelesning C-vitamin Enzymer i hverdagen C-vitamin eller askorbinsyre Finnes i svært mange frukter og grønnsaker Viktige kilder: appelsin paprika poteter C-vitamin Har mange viktige funksjoner i kroppen
TEKNOMAT ER DET SÅ ENKELT?
 TEKNOMAT ER DET SÅ ENKELT? Tor Lea Gruppe for molekylær cellebiologi Institutt for kjemi, bioteknologi og matvitenskap 2111 2005 Innhold Dagens situasjon Alternativ kjøttproduksjon In vitro- kjøtt/laboratoriekjøtt
TEKNOMAT ER DET SÅ ENKELT? Tor Lea Gruppe for molekylær cellebiologi Institutt for kjemi, bioteknologi og matvitenskap 2111 2005 Innhold Dagens situasjon Alternativ kjøttproduksjon In vitro- kjøtt/laboratoriekjøtt
Bruksanvisning IVD Matrix HCCA-portioned
 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til massespektrometri med matriksassistert laserdesorpsjonsionisasjonflytid (MALDI-TOF-MS). CARE- produkter er beregnet på å forsyne våre internasjonale
Antitrombin i laboratoriet
 Antitrombin i laboratoriet Carola Henriksson, Seksjonsoverlege, hematologiseksjonen, avdeling for medisinsk biokjemi, Oslo Universitetssykehus, Ullevål Hurdalsjøen, 8. juni Oslo Universitetssykehus (OUS)
Antitrombin i laboratoriet Carola Henriksson, Seksjonsoverlege, hematologiseksjonen, avdeling for medisinsk biokjemi, Oslo Universitetssykehus, Ullevål Hurdalsjøen, 8. juni Oslo Universitetssykehus (OUS)
Kap. 20 Mikrobiell vekstkontroll
 Kap. 20 Mikrobiell vekstkontroll Fokuset er her rettet mot reagenser og metoder som anvendes for vekstkontroll av mikrober utenfor kroppen Et utvalg av stoffert benyttes i det daglige liv: såper/vaskemidler
Kap. 20 Mikrobiell vekstkontroll Fokuset er her rettet mot reagenser og metoder som anvendes for vekstkontroll av mikrober utenfor kroppen Et utvalg av stoffert benyttes i det daglige liv: såper/vaskemidler
Bakgrunn Kvinner føder barn på et senere tidspunkt i livet enn tidligere Føder færre barn enn tidligere. Alder og fertilitet. Årsaker: Eggkvantitet
 Alder og fertilitet Mette Haug Stensen Senior klinisk embryolog Laboratoriesjef @ Bakgrunn Kvinner føder barn på et senere tidspunkt i livet enn tidligere Føder færre barn enn tidligere 23.05.14 Fakta:
Alder og fertilitet Mette Haug Stensen Senior klinisk embryolog Laboratoriesjef @ Bakgrunn Kvinner føder barn på et senere tidspunkt i livet enn tidligere Føder færre barn enn tidligere 23.05.14 Fakta:
STERIL KUN TIL ENGANGSBRUK IKKE-PYROGEN
 0086 TM Leveringssystem for legemidler til embolisering BRUKSANVISNING DC Bead Leveringssystem for legemidler til embolisering STERIL KUN TIL ENGANGSBRUK IKKE-PYROGEN Norsk BRUKSANVISNING DC Bead Leveringssystem
0086 TM Leveringssystem for legemidler til embolisering BRUKSANVISNING DC Bead Leveringssystem for legemidler til embolisering STERIL KUN TIL ENGANGSBRUK IKKE-PYROGEN Norsk BRUKSANVISNING DC Bead Leveringssystem
Klinisk molekylærmedisin (5): Eksempler på funksjonelle analyser
: Eksempler på funksjonelle analyser") Pediatrisk Endokrinologi 2003;17: 64-69 Klinisk molekylærmedisin (5): Eksempler på funksjonelle analyser Pål Rasmus Njølstad 1,2,3, Lise Bjørkhaug 1 1 Seksjon for pediatri, Institutt for klinisk medisin
Pediatrisk Endokrinologi 2003;17: 64-69 Klinisk molekylærmedisin (5): Eksempler på funksjonelle analyser Pål Rasmus Njølstad 1,2,3, Lise Bjørkhaug 1 1 Seksjon for pediatri, Institutt for klinisk medisin
Bruksanvisning IVD Matrix HCCA-portioned
 8290200 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til matriksassistert laserdesorpsjons-ionisasjon flytid for massespektrometri (MALDI-TOF-MS). CARE- produkter er utformet for å støtte
8290200 Bruksanvisning IVD Matrix HCCA-portioned Renset matriksstoff til matriksassistert laserdesorpsjons-ionisasjon flytid for massespektrometri (MALDI-TOF-MS). CARE- produkter er utformet for å støtte
Antioksidanter - mye eller nok
 Antioksidanter - mye eller nok Ole Terland Institutt for Biokjemi og Molekylær Biologi (IBMB), Universitetet i Bergen Oksidativt stress - Frie radikaler Molekylært oksygen er ansvarlig 1. Termodynamisk
Antioksidanter - mye eller nok Ole Terland Institutt for Biokjemi og Molekylær Biologi (IBMB), Universitetet i Bergen Oksidativt stress - Frie radikaler Molekylært oksygen er ansvarlig 1. Termodynamisk
NYHET! Basert på klinisk forskning Prisvinnende naturlig tomatekstrakt En kapsel om dagen
 Basert på klinisk forskning Prisvinnende naturlig tomatekstrakt En kapsel om dagen FOR GOD HJERTEHELSE* NYHET! SanoKardio inneholder et standardisert, tomatbasert ekstrakt med næringsstoffer som ivaretar
Basert på klinisk forskning Prisvinnende naturlig tomatekstrakt En kapsel om dagen FOR GOD HJERTEHELSE* NYHET! SanoKardio inneholder et standardisert, tomatbasert ekstrakt med næringsstoffer som ivaretar
Sammenligningen mellom Arabidopsis thaliana genomet og de kjente genomene fra cyanobakterier, gjær, bananflue og nematode, viser bl. a.
 Sammenligningen mellom Arabidopsis thaliana genomet og de kjente genomene fra cyanobakterier, gjær, bananflue og nematode, viser bl. a. Antall gener som er involvert i cellulær kommunikasjon og signaloverføring
Sammenligningen mellom Arabidopsis thaliana genomet og de kjente genomene fra cyanobakterier, gjær, bananflue og nematode, viser bl. a. Antall gener som er involvert i cellulær kommunikasjon og signaloverføring
Identifisering av mulige interaksjoner mellom NGFI-B og proteiner i BCL-2-familien
 HOVEDFAGSOPPGAVE FOR GRADEN Candidata pharmaciae Identifisering av mulige interaksjoner mellom NGFI-B og proteiner i BCL-2-familien Bjørg-Marie Grjotheim 2007 Avdelingen for Farmasøytisk Biovitenskap Farmasøytisk
HOVEDFAGSOPPGAVE FOR GRADEN Candidata pharmaciae Identifisering av mulige interaksjoner mellom NGFI-B og proteiner i BCL-2-familien Bjørg-Marie Grjotheim 2007 Avdelingen for Farmasøytisk Biovitenskap Farmasøytisk
Eksamensoppgave i PSY3111 Individuell utvikling, gener, nervesystem og atferd
 Psykologisk institutt Eksamensoppgave i PSY3111 Individuell utvikling, gener, nervesystem og atferd Faglig kontakt under eksamen: Dawn Behne Tlf.: Psykologisk institutt 73 59 19 60 Eksamensdato: 18.12.2014
Psykologisk institutt Eksamensoppgave i PSY3111 Individuell utvikling, gener, nervesystem og atferd Faglig kontakt under eksamen: Dawn Behne Tlf.: Psykologisk institutt 73 59 19 60 Eksamensdato: 18.12.2014
Transport av ioner og næringsstoffer over cellemembranen
 Transport av ioner og næringsstoffer over cellemembranen Terje Espevik, IKM Innhold: Diffusjonshastighet av molekyler over et lipiddobbeltlag Ionekonsentrasjoner innenfor og utenfor en typisk celle Transportere
Transport av ioner og næringsstoffer over cellemembranen Terje Espevik, IKM Innhold: Diffusjonshastighet av molekyler over et lipiddobbeltlag Ionekonsentrasjoner innenfor og utenfor en typisk celle Transportere