(12) Oversettelse av europeisk patentskrift
|
|
|
- Rudolf Egeland
- 8 år siden
- Visninger:
Transkript
1 (12) Oversettelse av europeisk patentskrift (11) NO/EP B1 (19) NO NORGE (51) Int Cl. A61K 31/70 ( ) A61K 38/16 ( ) A61K 39/395 ( ) A61P 25/00 ( ) C07H 21/00 ( ) C07K 14/00 ( ) C07K 16/00 ( ) C07K 16/28 ( ) C12N 5/20 ( ) C12N 15/13 ( ) Patentstyret (21) Oversettelse publisert (80) Dato for Den Europeiske Patentmyndighets publisering av det meddelte patentet (86) Europeisk søknadsnr (86) Europeisk innleveringsdag (87) Den europeiske søknadens Publiseringsdato (30) Prioritet , US, P (84) Utpekte stater AT BE BG CH CY CZ DE DK EE ES FI FR GB GR HR HU IE IS IT LI LT LU LV MC MT NL NO PL PT RO SE SI SK TR (73) Innehaver Biogen Idec MA Inc., 14 Cambridge Center, Cambridge, MA 02142, US-USA (72) Oppfinner MI, Sha, 4 Vernon Road, Belmont, MA 02478, US-USA PEPINSKY, R., Blake, 30 Falmouth Road, Arlington, MA 02474, US-USA SHAO, Zhaohui, 126 Babcock Street, 38, Brookline, MA 02466, US-USA GARBER, Ellen, A., 14 Donnell Street, Cambridge, MA 02138, US-USA MIKLASZ, Steven, D., 74 High Street, Upton, MA 01568, US-USA GRAFF, Christilyn, 632 Massachusetss Avenue nr. 614, Cambridge, Massachusetts 02139, US-USA (74) Fullmektig Zacco Norway AS, Postboks 2003 Vika, 0125 OSLO, Norge (54) Benevnelse SP 35-ANTISTOFFER OG BRUKEN AV DEM (56) Anførte publikasjoner WO-A2-2007/ WO-A2-2010/ US-A ALMAGRO JUAN C ET AL: "Humanization of antibodies", FRONTIERS IN BIOSCIENCE, FRONTIERS IN BIOSCIENCE, ALBERTSON, NY, US, vol. 13, 1 January 2008 ( ), pages , XP , ISSN: FOOTE J ET AL: "Antibody framework residues affecting the conformation of the hypervariable loops", JOURNAL OF MOLECULAR BIOLOGY, ACADEMIC PRESS, UNITED KINGDOM, vol. 224, no. 2, 20 March 1992 ( ), pages , XP , ISSN: , DOI: / (92)91010-M [retrieved on ]
 Prioritet 2007.01.")
2 1 SP 35-ANTISTOFFER OG BRUKEN AV DEM 5 Oppfinnelsens område OPPFINNELSENS BAKGRUNN 10 [0001] Denne oppfinnelsen relaterer seg til nevrologi, nevrobiologi og molekylarbiologi. Mer spesifikt relaterer denne oppfinnelsen seg til Sp 35-antistoffer samt disse antistoffene til bruk i behandling av nevrologiske sykdommer, lidelser og skader som f.eks. ryggmargsskader. Oppfinnelsens bakgrunn [0002] Aksoner og dendritter strekker seg fra nevroner. Den distale spissen til et utstrakt akson eller en utstrakt nevritt inkluderer et spesialisert område som kalles en voksende tapp (growth cone). Voksende tapper registrerer de lokale omgivelsene og styrer aksonal vekst mot en målcelle til et nevron. Voksende tapper reagerer på miljøsignaler som f.eks. overflatebindekraft, vekstfaktorer, signalsubstanser og elektriske felt. De voksende tappene beveger seg fremover generelt ved en hastighet på én eller to millimeter per dag. De voksende tappene undersøker områdene rundt dem og på begge sidene ved forlengelser som klassifiseres som lamellipodier og filopodier. Når en forlengelse kommer i kontakt med en ugunstig flate, trekker den seg tilbake. Når en forlengelse kommer i kontakt med en gunstig vekstflate, forsetter den å strekke seg og styrer den voksende tappen i den retningen. Når den voksende tappen når en egnet målcelle, dannes en synoptisk forbindelse. [0003] Nervecellefunksjonen påvirkes av kontakten mellom nevroner og andre celler i deres nærmeste omgivelse (Rutishauser, et al., 1988, Physiol. Rev. 68:819). Disse cellene inkluderer spesialiserte gliaceller, oligodendrocytter i det sentrale nervesystemet (CNS) og Schwann-celler i det perifere nervesystemet (PNS) som dekker nervefiberen med myelin (Lemke, 1992, i An Introduction to Molecular Neurobiology, Z. Hall, Ed., p. 281, Sinauer). [0004] CNS-nevroner har naturlig potensiale til å regenerere etter en skade, men de hindres i dette av hemmende proteiner som finnes i myelin (Brittis et al., 2001, Neuron 30:11-14; Jones et al., 2002, J. Neurosci. 22: ; Grimpe et al., 2002, J. Neurosci.:22: ). [0005] Flere myelinhemmende proteiner som finnes på oligodendrocytter, er blitt karakterisert. Kjente eksempler på myelinhemmende proteiner inkluderer NogoA (Chen et al., Nature, 2000, 403, ; Grandpre et al., Nature 2000, 403, ), myelin associated glycoprotein (myelinassosiert glykoprotein) (MAG) (McKerracher et al., 1994, Neuron 13: ; Mukhopadhyay et al., 1994, Neuron 13: ) og oligodendrocyttglykoprotein (OM-gp), Mikol et al., 1988, J. Cell. Biol.106: ). Hvert av disse proteinene har hver for seg vist seg å være en ligand til den nevronale Nogo-reseptoren-1 (NgR1 (Wang et al., Nature 2002, 417, , Grandpre et al., Nature 2000, 403, , Chen et al., Nature, 2000, 403, , Domeniconi et al., Neuron 2002, publisert på Internett 28. juni 2002).
![Oppfinnelsens bakgrunn 15 20 25 30 35 40 [0002] Aksoner og dendritter strekker seg fra nevroner.](/docs-images/56/5927828/images/page_2.jpg "Den distale spissen til et utstrakt akson eller en utstrakt nevritt inkluderer et spesialisert område som kalles en voksende tapp (growth cone).")
3 [0006] Nogo-reseptor-1 (NgR1) er et GPI-forankret membranprotein som inneholder 8 leucinrike gjentakelser (Fournier et al., 2001, Nature 409: ). Ved samhandling med hemmende proteiner (f.eks. NogoA, MAG og OM-gp), sender NgR1-komplekset signaler som fører til sammenbrudd av den voksende tappen og hemming av nevrittutvekst. [0007] Det finnes et udekket behov for molekyler og metoder for å hemme NgR1-mediert sammenbrudd av den voksende tappen og den resulterende hemming av nevrittutvekst. I tillegg finnes det et behov for molekyler som øker nevronoverlevelse og aksonregenerering. Spesielt ved behandling av sykdom, lidelser eller skader som involverer aksonskade, nevron- eller oligodendrocyttcelledød, demyelinisering eller demyelinering eller generelt relatert til nervesystemet. [0008] Slike sykdommer, lidelser eller skader inkluderer, men er ikke begrenset til multippel sklerose (MS), progressiv multifokal leukoencefalopati (PML), enkefalomyelitt (EPL), sentral pontin myelolyse (CPM), adrenoleukodystrofi, Alexanders sykdom, Pelizaeus Merzbacher-sykdom (PMZ), globoid celleleukodystrofi (Krabbes sykdom) og Wallerisk degenerasjon, optikusnevritt, transversell myelitt, amylotrofisk-lateralsklerose (ALS), Huntingtons sykdom, Alzheimers sykdom, Parkinsons sykdom, ryggmargsskade, traumatisk hjerneskade, skader etter stråling, nevrologiske komplikasjoner ved kjemoterapi, slag, akutt iskemisk optisk nevropati, vitamin E-mangel, isolert vitamin E-mangel syndrom, AR, Bassen-Kornzweig-syndrom, Marchiafava-Bignami-syndrom, metakromatisk leukodystrofi, trigeminusnevralgi og ansiktslammelse. Blant disse sykdommene er MS mest utbredt og påvirker omtrent 2,5 millioner personer globalt. [0009] MS begynner generelt med et tilbakefallende-remitterende mønster som involverer nervene, som deretter utvikler seg til en kronisk fase med økende nevrologisk skade. MS er forbundet med destruksjon av myelin, oligodendrocytter og aksoner som befinner seg i kroniske lesjoner. Demyelinering som observeres i MS, er ikke alltid permanent og remyelinering er dokumentert i tidlige sykdomstrinn. Remyelinering av nevroner krever oligodendrocytter. [0010] Det finnes forskjellige sykdomsmodifiserende behandlinger av MS inkludert bruken av kortikosteroider og immunomodulatorer som interferon beta og Tysabri. I tillegg, på grunn av den sentrale rollen til oligodendrocytter og myelinering i MS, er det gjort tiltak for å utvikle terapier for å øke antall oligodendrocytter eller forbedre myelinering. Se f.eks. Cohen et al., amerikansk patentnr. 5,574,009; Chang et al., N. Engl. J. Med:. 346: (2002). (WO 2007/ offentliggjør antistoffer som er spesifisert for Sp35 og metoder som bruker slike antistoffer som antagonister av endogen Sp35-funksjon.) Det finnes imidlertid fremdeles et pressende behov for å finne opp flere terapier for MS og andre demyelinerings- og disemyelinerings-lidelser KORT SAMMENDRAG AV OPPFINNELSEN [0011] Den gjeldende offentliggjøringen er basert på oppdagelsen av at Sp35 (Sp35 kalles også LINGO- 1 og LRRN6 i litteraturen) uttrykkes i oligodendrocytter and nerveceller og regulerer oligodendrocytt- /nevrondifferensiering, overlevelse og aksonmyelinering negativt. I tillegg fremmer visse antagonister av Sp35 overlevelse, spredning og differensiering av oligodendrocytter og nerveceller samt myelinering av nevroner. Basert på disse oppdagelsene, relatere oppfinnelsen som inkluderer den
![[0007] Det finnes et udekket behov for molekyler og metoder for å hemme NgR1-mediert sammenbrudd av den voksende tappen og den resulterende hemming av nevrittutvekst.](/docs-images/56/5927828/images/page_3.jpg "I tillegg finnes det et behov for molekyler som øker nevronoverlevelse og aksonregenerering.")
4 gjeldende oppfinnelsen, seg generelt til antistoffer, antigenbindende fragment eller avledning av dem som kan brukes som antagonist av Sp35. Mer spesifikt gir den gjeldende oppfinnelsen et isolert antistoff eller et antigenbindende fragment av det som kan spesifikt bindes til et Sp35-polypeptid der antistoffet eller fragmentet av det omfatter et VH-område som omfatter VH CDR1-, CDR2- og CDR3-sekvenser av henholdsvis SEQ ID NR:436, SEQ ID NR:437 og SEQ ID NR:438 og et VL-område som omfatter VL CDR1-, CDR2- og CDR3-sekvenser av henholdsvis SEQ ID NR:442, SEQ ID NR:443 og SEQ ID NR:444 og der fragmentet av antistoffet binder seg til et Sp35-polypeptid med affinitet som karakteriseres av en dissosiasjonskonstant som ikke er større enn 5 x M som målt med FACS på CHO-celler som er stabilt transfektert med humant Sp35. Det finnes flere aspekter og utforminger av oppfinnelsen i de vedlagte patentkravene. 20 KORT BESKRIVELSE AV TEGNINGENE/FIGURENE [0027] FIG. 1: SDS-PAGE-gel viser immunopresipitering av Sp35 med de monoklonale antistoffene 1A7 og 2F3. [0028] FIG.2: FACS-resultatet viser at MAb-1A7 og -2F3 bindes til COS-7- eller 293-cellene som uttrykker Sp35, men ikke for å kontrollere celler uten Sp35-uttrykk. [0029] FIG.3: MAb-1A7 og -2F3 beskytter DRG-nevroner mot myelinmediert hemming av nevrittutvekst. [0030] FIG.4A-G: Immunohistokjemisk farging (IHC) av ko-kulturer av DRG-nevroner og oligodendrocytter behandlet med monoklonale antistoffer 1A7 og 2F3 eller kontroll-antistoff. Panelene D og E er forstørrelser av henholdsvis panelene B og C. Farging av anti-βiii-tubuling-antistoff for å indentifisere aksoner eller anti-mbp-antistoffer for å identifisere oligodendrocytter. F: Kvantitering av MBP+ myelinerende celler ved behandling av ko-kulturer med 1A7 eller 2F3. G: Western blot-analyse for å kvantifisere MBP-en som produseres fra ko-kulturer av DRG-nevroner og oligodendrocytter behandlet med monoklonale antistoffene 1A7 og 2F3. [0031] FIG. 5A-C: A: CC1-antistoff-farging av oligodendrocytter fra mus i cuprizon-modellen. B. Anti- MBP- proteinantistoff eller Luxol Fast Blue-farging av musenevroner i Cuprizon-modellen. C: Kvantifisering av CC1-antistoff-positive oligodendrocytter ved fire uker og seks uker. [0032] FIG. 6: Overlevende RGC-er. Ved behandling med monoklonalt antistoff 1A7 anti-sp35- antistoff, viste 1A7-behandlede dyr signifikant nevral overlevelse (80 %) sammenlignet med kontrollantistoff eller PBS-behandlede dyr som begge kun utviser omtrent 50 % nevral overlevelse. [0033] FIG. 7. BBB-skårer av musemottakende anti-sp35-antistoff 1A7 etter ryggmargsskader som beskrevet i eksempel 8. [0034] FIG. 8. Western blot av samdyrkede oligodendrocytter og DRG-er etter inkubering med anti- Sp35-antistoffene Li05, Li06 og 3, 10 and 30 mg Sp35-Fc (LINGO-1-Ig) som beskrevet i eksempel 9.

5 [0035] FIG. 9. Fotoer av synsnerver av A) normale rotter, B) myelinoligodendrocyttglykoprotein (MOG)-indusert eksperimentale autoimmunenkefalomyelititt (EAE)- rotter og C) myelinoligodendrocyttglykoprotein (MOG) indusert eksperimentale autoimmun enkefalomyelitt (EAE)-rotter behandlet med Sp35-antistoff 1A7. Elektronmikrografer av hver synsnerve vises nedenfor i hvert foto av synsnerven. [0036] FIG. 10. Diagram med antall regenerative nervefibre per del regnet i dyr som mottar en intravitreal injeksjon av Sp35-antistoff 1A7 etter synsnerveknusing. [0037] FIG. 11. FACS-resultatet viser at MA-3B5.2 (3BS) og -7P1D5.1G9 (1D5) bundet til CHO-celler som er stabilt transfektert med Sp35 (LINGO-1). DETALJERT BESKRIVELSE AV OPPFINNELSEN I. DEFINISJONER [0038) Det gjøres oppmerksom på at begrepet «en» eller «et» henviser til én eller flere av den enheten, f.eks. «et Sp35-antistoff» forstås som å representere ett eller flere Sp35-stoffer. Dermed kan begrepet «en» (eller «ett»), «én/ett eller flere og «minst én/ett» brukes om hverandre i dette dokumentet. [0039] Slik som brukt i dette dokumentet er begrepet «polypeptid» beregnet på å omfatte et enkelt «polypeptid» samt flere «polypeptider» og henviser til et molekyl som omfatter monomerer (aminosyrer) som er lineært koplet til amidbindinger (kalles også peptidbindinger). Begrepet «polypeptid» henviser til en hvilken som helst kjede eller kjeder til to eller flere aminosyrer, og henviser ikke til en bestemt lengde av produktet. Dermed er peptider, dipeptider, tripeptider, oligopeptider, «protein», «aminosyrekjede» eller andre begrep som brukes for å henvise til en kjede eller kjeder til to eller flere aminosyrer, inkludert innenfor definisjonen av «polypeptid» og begrepet «polypeptid» kan brukes istedenfor eller byttes med et hvilket som helst av disse begrepene. Begrepet «polypeptid» er også beregnet på å henvise til produkter med modifikasjoner etter uttrykk av polypeptidet, inkludert uten begrensning glykosylering, acetylering, fosforylering, amidatering, avledning av kjente beskyttelses-/blokkerende grupper, proteolytisk spalting eller modifikasjon av ikke-naturlig oppstående aminosyrer. Et polypeptid kan avledes fra en naturlig biologisk kilde eller produsert med rekombinant teknologi, men translateres nødvendigvis ikke fra en utpekt nukleinsyresekvens. Det kan genereres på en hvilken som helst måte, inkludert kjemisk syntese. [0040] Et polypeptid av oppfinnelsen kan ha en størrelse på omtrent 3 eller flere, 5 eller flere, 10 eller flere, 20 eller flere, 25 eller flere, 50 eller flere, 75 eller flere, 100 eller flere, 200 eller flere, 500 eller flere, 1000 eller flere eller 2000 eller flere aminosyrer. Polypeptidene kan ha en bestemt tredimensjonalt struktur, selv om de ikke nødvendigvis har en slik struktur. Polypeptider med bestemt tredimensjonal struktur kalles foldet, og polypeptider som ikke har bestemt tredimensjonal struktur, men i stedet kan tilpasses et stort antall forskjellige former og kalles ufoldet. Slik som brukt i dette dokumentet, henviser begrepet «glykoprotein» til et protein som er koplet til minst én
![indusert eksperimentale autoimmun enkefalomyelitt (EAE)-rotter behandlet med Sp35-antistoff 1A7. Elektronmikrografer av hver synsnerve vises nedenfor i hvert foto av synsnerven. [0036] FIG. 10.](/docs-images/56/5927828/images/page_5.jpg "Diagram med antall regenerative nervefibre per del regnet i dyr som mottar en intravitreal injeksjon av Sp35-antistoff 1A7 etter synsnerveknusing. [0037] FIG. 11. FACS-resultatet viser at MA-3B5.")
6 karbohydratmoietet som er festet til proteinet via en sidekjede som inneholder oksygen eller nitrogen til en aminosyrerest, f.eks. serinrest eller asparaginrest. [0041] Ved et «isolert» polypeptid eller et fragment, en variant eller avledninger av det som er beregnet som et polypeptid som ikke befinner seg i dets naturlige omgivelser. Det kreves ikke noe bestemt renhetsnivå. Et isolert polypeptid kan f.eks. fjernes fra dets native eller naturlige omgivelser. Rekombinantproduserte polypeptider og proteiner som uttrykkes i vertscellene regnes som isolert når det gjelder offentliggjøringen, og det samme gjelder native eller rekombinante polypeptider som er separert, fraksjonert eller delvis eller stort sett renset med en hvilken som helst egnet metode. [0042] Det som også inkluderes som polypeptider i den gjeldende offentliggjøringen er fragmenter, avledninger, analoger eller varianter av de førnevnte polypeptidene eller en hvilken som helst kombinasjon av dem. Når begrepene «fragment», «variant», «avledning» og «analog» henviser til Sp35-antistoffer eller antistoffpolypeptider, inkluderes alle polypeptider som beholder minst noen av de antigenbindende egenskapene til det tilstedeværende native antistoffet eller polypeptidet. Fragmenter av polypeptider inkluderer proteolytiske fragmenter samt slettingsfragmenter i tillegg til bestemte antistoffragmenter som omtales et annet sted i dette dokumentet. Varianter av Sp35- antistoffer og antistoffpolypeptider inkluderer fragmenter som omtales ovenfor i tillegg til polypeptider med endrede aminosyresekvenser som er forårsaket av aminosyreerstatninger, - slettinger eller innsettinger. Varianter kan oppstå naturlig eller skje unaturlig. Varianter som skjer unaturlig, kan produseres ved å bruke artskjente mutagenesemetoder. Variantpolypeptider kan omfatte konservative eller ikke-konservative aminosyreerstatninger, -slettinger eller -tillegg. Avledninger av Sp 35-antistoffer og antistoffpoplypeptider eller polypeptider er blitt endret slik at de utviser tilleggsfunksjoner som ikke finnes i det native polypeptidet. Eksempler inkluderer fusjonsproteiner. Variantpolypeptider kan også kalles «polypeptidanologer» i dette dokumentet. Slik «avledning» av et Sp35-antistoff eller antistoffpolypeptid brukes i dette dokumentet, henviser det til det aktuelle polypeptidet med én eller flere rester som er kjemisk avledet fra reaksjonen til en funksjonell sidegruppe. I tillegg inkluderer «avledninger» peptider som inneholder én eller flere naturlige forekomster av aminosyreavledninger til tjue standard aminosyrer. 4 hydroksyprolin kan f.eks. erstatte prolin. 5-hydroksylysin kan erstatte lysin, 3-metylhistidin kan erstatte histidin. Homoserin kan erstatte serin og ornitin kan erstatte lysin. [0043] Begrepet «polynukleotid» er beregnet på å innbefatte en enkel nukleinsyre samt flere nukleinsyrer og henviser til et isolert nukleinsyremolekyl eller konstruksjon, f.eks. budbringer-rna (mrna) eller plasmid-dna (PDNA). Et polynukleotid kan omfatte en konvensjonell fosfodiesterbinding eller en ikke-konvensjonell binding (f.eks. en amidbinding som finnes i peptidnukleinsyrer (PNA)). Begrepet «nukleinsyre» henviser til ett eller flere nukleinsyresegmenter, f.eks. DNA- eller RNAfragmenter som finnes i et polynukleotid. En «isolert» nukleinsyre og et «isolert» polynukleotid betegner et nukleinsyremolekyl, DNA eller RNA som er fjernet fra sine naturlige omgivelser. Et rekombinant polynukleotid som koder et Sp 35-antistoff som f.eks. inneholder en vektor, regnes som isolert når det gjelder den gjeldende offentliggjøringen. Andre eksempler på et isolert polynukleotid inkluderer rekombinante polynukleotider som opprettholdes i heterologe vertsceller eller rensede

7 (delvis eller stort sett) polynukleotider i løsning. Isolerte RNA-molekyler inkluderer in vivo eller in vitro RNA-transkripsjoner av polynukleotider. I tillegg inkluderer isolerte polynukleotider eller nukleinsyrer slike molekyler som produseres syntetisk. I tillegg kan polynukleotider eller en nukleinsyre være med eller uten et regulerende element som f.eks. en promoter, et ribosomt bindingssted eller en transkripsjonsterminator. [0044] Slik som brukt i dette dokumentet er en «kodesekvens» en del av en nukleinsyre som består av kodoner som er translatert til aminosyrer. Selv om «stoppkodon» (TAG, TGA, or TAA) ikke translateres til en aminosyre, kan den regnes som en del av en kodesekvens, men eventuelle flankerende sekvenser, f.eks. promoterere, ribosome bindingssteder, transkripsjonsterminatorer, introner og lignende, er ikke en del av en kodesekvens. To eller flere kodesekvenser av den gjeldende offentliggjøringen, kan finnes i en enkel polynukleotidkonstruksjon, f.eks. en enkel vektor, eller i separate polynukleotide konstruksjoner, f.eks. separate (forskjellige) vektorer. I tillegg kan en hvilken som helst vektor innehold en enkel kodesekvens, eller omfatte to eller flere kodesekvenser, f.eks. kan en enkel vektor kode en variabel immunoglobulinsekvens med tungkjede og en variabel immunoglobulinsekvens med lettkjede. I tillegg kan en vektor, et polynukleotid eller en nukleinsyre til offentliggjøringen, kode heterologe kodesekvenser, enten sammenfestet eller ikke sammenfestet til en nukleinsyre som koder et Sp35-antistoff eller fragment, variant eller avledning av det. Heterologe kodesekvenser inkluderer, uten begrensninger, spesialiserte elementer eller motiver, som f.eks. et sekretorisk signalpeptid eller et heterologt funksjonelt domene. [0045] I visse tilfeller er polynukleotidet eller nukleinsyren DNA. I tilfellet med DNA kan polynukleotidet omfatte en nukleinsyre som koder en polypeptid normalt, inkludere en promoter og/eller andre transkripsjons- eller translasjonskontrollelementer som er driftsmessig forbundet med én eller flere kodesekvenser. En driftsforbindelse er når en kodesekvens til et genprodukt, f.eks. et polypeptid, er forbundet med én eller flere regulerende sekvenser på en slik måte at det plasseres uttrykk av genproduktet under innflytelse eller kontroll av den/de regulerende sekvensen(e). To DNAfragmenter (som f.eks. polypeptidkodesekvens og en promoter forbundet med den) er «driftsforbundet» hvis induksjonen av promoterfunksjonen fører til transkripsjonen av mrna-koding gir det ønskede genproduktet, og hvis karakteren til koplingen mellom de to DNA-fragmentene ikke forstyrrer muligheten til at uttrykksregulerende sekvenser kan rette uttrykket til genproduktet eller forstyrre muligheten til at DNA-malet kan transkriberes. Dermed ville en promotorsekvens være driftsforbundet med en nukleinsyre som koder et polypeptid hvis promoteren har muligheten til effektiv transkripsjon av den nukleinsyren. Promotoren kan være en cellespesifikk promoter som retter vesentlig transkripsjon av DNA-en kun i forhåndsbestemte celler. Andre transkripsjonskontrollelementer, utenom en promoter, f.eks. enhancere, operatører, undertrykkere og transkripsjons-avslutningssignaler, kan være driftsforbundet med polynukleotidet for å styre cellespesifikk transkripsjon. Egnede promoterer og andre transkripsjons-kontrollsekvenser offentliggjøres i dette dokumentet. [0046] En rekke transkripsjons- kontrollsekvenser er kjent for personer med ferdigheter i faget. Disse inkluderer, uten begrensning, transkripsjons-kontrollsekvenser som funksjon i virveldyrceller, som f.eks., men ikke begrenset til promotorer- og enhancersegmenter fra cytomegaloviruser (tidlig

8 promoter i forbindelse med intron-a), apevirus 40 (tidlig promoter) og retroviruser (som f.eks. Roussarkomavirus). Andre transkripsjons-kontrollsekvenser inkluderer de som er avledet fra virveldyrgener som aktin, heat shock-proteiner, bovine veksthormoner og hare-β-globin samt andre sekvenser som kan kontrollere genuttrykk i eukaryotiske celler. Andre egnede transkripsjons-cellesekvenser inkluderer vevsspesifikke promoterere og enhancere samt lymfokininduserte promotere (f.eks. promoterere som kan slutte seg til interferoner og interleukiner). [0047] På lignende måte er en rekke translasjons-kontrollelementer kjent hos personer med vanlige ferdigheter i faget. Disse inkluderer, men er ikke begrenset til ribosome bindingssteder, translasjonsinnføring og avslutningskodoner og elementer avledet fra pikornaviruser (spesielt et internt ribosomt inngangssted eller IRES som også kalles en CITE-sekvens). [0048] I andre tilfeller er f.eks. polynukleotidet til den gjeldende offentliggjøringen RNA som er utformet som budbringer-rna (mrna). [0049] Polynukleotid- og nukleinsyrekodesekvenser til den gjeldende offentliggjøringen, kan være forbundet med flere andre kodesekvenser som koder sekretoriske eller signalpeptider som styrer utskillingen til et polypeptid kodet av et polynukleotid til den gjeldende offentliggjøringen. Iht. signalhypotesen er proteiner som utskilles av pattedyrceller et signalpeptid eller sekretoriske ledersekvenser som er spaltet fra det modne proteinet etter at eksport av det voksende proteinkjedet over den ujevne endoplasmiske retikulum, er satt i gang. Personer med vanlige ferdigheter i faget kjenner til at polypeptider som skilles ut av virveldyrceller, har generelt et signalpeptid som er festet til N-terminus til polypeptid, som er spaltet fra hele eller polypeptidet med «full lengde» for å produsere en utskilt eller «moden» type polypeptid. I visse tilfeller bruker det native signalpeptidet, f.eks. et tungkjede-immunoglobulin eller lettkjede-signalpeptid eller en funksjonell avledning av denne sekvensen som beholder evnen til å styre utskillelsen av polypeptidet som er driftsforbundet med det. Som alternativ kan et heterologt pattedyrsignalpeptid eller funksjonell avledning av det brukes. Den ville typen ledersekvens kan erstattes med ledersekvensen til humant plasminogen aktivator (TPA) eller mus-β-glukuronidase. [0050] Den gjeldende offentliggjøringen rettes mot visse Sp35-antistoffer eller antigenbinding, fragmenter, varianter eller avledning av dem. Med mindre det spesifikt henvises til antistoffer med full størrelse som f.eks. naturlig forekommende antistoffer, innbefatter begrepet «Sp35-antistoffer» antistoffer med full størrelsen samt antigenbindende fragmenter, varianter, analoger eller avledningen av slike antistoffer, f.eks. naturlig forekommende antistoff eller immunoglobinmolekyler eller konstruerte antistoffmolekyler eller fragmenter som binder antigen på en måte som ligner på antistoffmolekyler. [0051] Begrepene «antistoff» og «immunoglobulin» brukes om hverandre i dette dokumentet. Et antistoff eller immunoglobulin omfatter minst det variable domenet til en tungkjede og omfatter normalt minst de variable domene til en tungkjede eller en lettkjede. Grunnleggende immunoglobulinstrukturer i virveldyrsystemer er forholdsvis godt forstått. Se f.eks. Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2. utgave 1988). [0052] Slik som omtalt tidligere, omfatter begrepet «immunoglobulin» grove klasser av polypeptider som kan skjelnes biokjemisk. Personer med ferdigheter i faget vil sette pris på at tunge kjeder er
. [0047] På lignende måte er en rekke translasjons-kontrollelementer kjent hos personer med vanlige ferdigheter i faget.")
9 klassifisert som gamma, mu, alpha, delta eller epsilon, (γ, μ, α, δ, ε) med noen underklasser blant dem (f.eks. γ1-γ4). Det er karakteren til dette kjedet som bestemmer antistoffets «klasse» som henholdsvis IgG, IgM, IgA IgG eller IgE. Immunoglobulin-underklassene (isotypene) f.eks. IgG 1, IgG 2, IgG 3, IgG 4, IgA 1, osv. er godt karakterisert, og det er kjent at de gir funksjonell spesialisering. Modifiserte versjoner av hver av disse klassene og isotypene er lett å skjelne for er person med ferdigheter når det gjelder omgående offentliggjøring og er derfor innenfor omfanget til offentliggjøringen. Alle immunoglobinklassene er tydelig innenfor omfanget til offentliggjøringen. Følgende omtale vil være generelt rettet mot IgG-klassen med immunoglobulinmolekyler. Når det gjelder IgG, omfatter en standard immunoglobulinmolekyl to identiske lette kjedepolypeptider med en molekylvekt på omtrent daltoner og to identiske tunge kjedepolypeptider med en molekylvekt på De fire kjedene er vanligvis koplet sammen med disulfidbindering i en «Y»-konfigurasjon der de lette kjedene understøtter de tunge kjedene fra begynnelsen av «Y»- munnen og videre gjennom det variable området. [0053] De lette kjedene er klassifiserte enten som kappa eller lambda (κ, λ). Hver tungkjede-klasse kan være bundet enten med en kappa eller lambda lettkjede. Generelt er lette og tunge kjeder kovalent bundet til hverandre, og «haledelene» til de to tunge kjedene er bundet til hverandre med kovalente disulfidkoplinger eller ikke-kovalente kopling når immunoglobulinene er generert enten av hybridomer, B-celler eller genetisk konstruerte vertsceller. I den tunge kjeden, kjøres aminosyresekvensene fra en N-terminus i grenenden av «Y»-konfigurasjonen til C-terminum i bunnen av hvert kjede. [0054] Både de lette og tunge kjedene er delt inn i områder med strukturell og funksjonell homologi. Begrepene «konstant» og «variabel» brukes funksjonelt. Når det gjelder dette, vil det bli satt pris på at de variable domenene til både den lette (V L ) og tunge (V H ) kjedeleden bestemmer antigengjenkjenning og spesifisitet. Omvendt overfører konstante domene til det lette kjedet (C L ) og det tunge kjedet(c H 1, C H 2 or C H 3) viktige biologiske egenskaper som utskilling, transplasensial mobilitet, Fc-reseptor-binding, komplementbinding og lignende. Som akseptert prinsipp øker antall konstantområde-domener i det de blir mer distale fra antigenbindingsstedet eller amino-terminus til antistoffet. N-terminaldelen er et variabelt område og C-terminaldelen er et konstant område. C H 3- og C L -domer omfatter faktisk karboksyterminus til henholdsvis tunge og lette kjeder. [0055] Som indikert ovenfor, muliggjør det variable området at antistoffet kan selektivt gjenkjenne og spesifikt binde epitoper på antigener. Dvs. at V L -domenet og V H -domenet eller subsett av de komplementære bestemmende områdene (CDR-er) til et antistoff, forenes og danner et variabelt område som utgjør et tredimensjonalt antigenbindingssted. Denne kvantenære antistoffstrukturen danner antigenbindingsstedet som finnes på enden av hver arm til «Y»-en. Mer spesifikt utgjør antigenbindingsstedet tre CDR-er på hvert av V H - og V L -kjedene. I noen tilfeller, f.eks. visse immunoglobinmolekyler avledet fra kamelarter eller konstruert basert på kamelimmunoglobuliner, kan et komplett immunoglobulingmolekyl bestå kun av tunge kjeder uten lette kjeder. Se f.eks. Hamers-Casteman et al., Nature 363: (1993). [0056] I naturlig forekommende antistoffer, er de seks «komplementære bestemmende områdene» eller «CDR-ene» som finnes i hvert antigenbindingsdomene, korte, ikke-sammenhengende sekvenser

10 med aminosyrer som er spesifikt plassert for å danne antigenbindingsdomenet i det antistoffet som inntar sitt tredimensjonale konfigurasjon i et vannholdig miljø. Resten av aminosyrene i antigenbindingsdomene som kalles «rammeverksområder», utviser mindre intermolekylær variabilitet. Rammeverksområdene tilpasser seg stort sett en β-ark-konfigurasjon og CDR-ene danner sløyfer som koples, og i noen tilfeller danner de en del av β-ark-strukturen. Dermed fungerer rammeverksområdene som en stillas som gir posisjonering av CDR-er i riktig retning med interkjede-, ikke-kovelente interaksjoner. Antigenbindingsdomenet som dannes av de posisjonerte CDR-ene, utgjør en flate som er komplementær med epitopen på det immunoreaktive antigenet. Denne komplementære flaten fremme ikke-kovalent binding av antistoffet til den kognate epitopen. Aminosyrene som omfatter henholdsvis CDR-ene og rammeverksområdene, kan lett identifiseres for et hvilket som helst bestemt variabelt område med tung- eller lettkjede, av en person med vanlige ferdigheter i faget fordi de er presist definert (se «Sequences of Proteins of Immunological Interest,» Kabat, E., et al., U.S. Department of Health and Human Services, (1983) og Chothia and Lesk, J. Mol. Biol., 196: (1987). [0057] I tilfellet der to eller flere definisjoner av et begrep brukes og/eller aksepteres innen faget, er definisjonen av begrepet som det brukes i dette dokumentet tiltenkt å inkludere alle slike betydninger med mindre det uttrykkelig står det motsatte. Et spesifikt eksempel er bruken av begrepet «komplementært bestemmende område» (CDR) for å beskrive de ikke-sammenhengende antigenkombinerende steder som finnes inne i det variable området til både de lette og tunge kjedepolypeptidene. Dette bestemte området beskrives av Kabat et al., U.S. Dept. of Health and Human Services, «Sequences of Proteins of Immunological Interest» (1983) og av Chothia et al., J. Mol. Biol. 196: (1987), der definisjonene inkluderer overlapping eller subsett av aminosyrerester når de sammenlignes med hverandre. Ikke desto mindre er det tiltenkt at bruken av begge definisjonene til en CDR av et antistoff eller varianter av det, skal være innenfor omfanget av begrepet slik det defineres og brukes i dette dokumentet. De egnede aminosyrerestene som omfatter CDR-ene som definert i hver av de henviste referansene, presenteres i tabell 1 til sammenligning. De nøyaktige restetallene som omfatter en bestemt CDR, vil variere avhengig av sekvensen og størrelsen til CDR-en. Personer med ferdigheter i faget kan på regelmessige basis bestemme hvilke rester omfatter en bestemt CDR når det variable området og aminosyresekvensen til antistoffet er oppgitt. TABELL 1. CDR-definisjoner l Nummerering av alle CDR-definisjonene i tabell 1 er iht. de generelle prinsippene som legges frem av Kabat et al. (se nedenfor)...

11 [0058] Kabat et al. har også definert nummereringssystemet til variable domenesekvenser som gjelder et hvilket som helst antistoff. En person med vanlige ferdigheter i faget kan klart tildele dette systemet med «Kabat-nummerering» til en hvilken som helst variabel domenesekvens uten å måtte stole på noen forsøksdata utenom selve sekvensen. Slik som «Kabat-nummerering» brukes i dette dokumentet, henviser det til nummereringssystemet som presenteres av Kabat et al., U.S. Dept. of Health and Human Services, «Sequence of Proteins of Immunological Interest" (1983). Med mindre noe annet spesifiseres, er henvisninger til nummerering av bestemte aminosyrerestdeler i et Sp35-antistoff eller antigenbindende fragment, variant, eller avledning av det iht. Kabat-nummereringssystemet. [0059] I kamelarter danner det variable området med tungkjede og som kalles V H H, hele antigenbindingsdomenet. Hovedforskjellen mellom kamel-v H H variable områder og de som er avledet fra konvensjonelle antistoffer (V H ) inkluderer (a) flere hydrofonaminosyrer i kontaktflaten i det lette kjedet i V H i forhold til det tilsvarende området i V H H, (b) større CDR3 i V H H og (c) hyppig forekomst av en disulfidbinding mellom CDR1 og CDR3 i V H H. [0060] Antistoffer eller antigenbindende fragmenter, varianter eller avledninger av offentliggjøringen inkludert, men ikke begrenset til polyklonale, monoklonale, multispesifikke, humane, humaniserte, primatiserte eller kimære antistoffer, antistoffer med enkelkjede, epitopbindende fragmenter, f.eks. Fab, Fab' og F(ab') 2, Fd, Fvs, enkeltkjedet Fvs (scfv), enkeltkjedede antistoffer, disulfidkoplet Fvs (sdfv), fragment som omfatter enten V L - eller V H -domene, fragmenter produsert av et Fabuttrykksbibliotek og antiidiotypiske (anti-id) antistoffer (inkludert f.eks. anti-id-antistoffer til Sp35- antistoffer som offentliggjøres i dette dokumentet). ScFv-molekyler er kjent innen faget og beskrives f.eks. i det amerikanske patentet 5,892,019. Immunoglobulin- eller antistoffmolekyler til offentliggjøringen kan være en hvilken som helst type (f.eks. IgG, IgE, IgM, IgD, 1gA og IgY), klasse (f.eks. IgG1, IgG2, IgG3, IgG4, 1gA1 og 1gA2) eller underklasse av immunoglobulinmolekylet. [0061] Antistoff-fragmenter inkludert enkeltkjedede antistoffer, kan omfatte det/de variable området/områdene alene eller sammen med hele eller en del av følgende: hengselområde, C H 1-, C H 2- og C H 3-domene. Offentliggjøringen inkluderer også antigenbindende fragmenter som omfatter en hvilken som helst kombinasjon av et variabelt område / variable områder med et hengselsområde, C H 1- C H 2- og C H 3-domene. Antistoffer eller immunospesifikke fragmenter av dem til bruk i diagnostiske og terapeutiske metoder som offentliggjøres i dette dokumentet, kan ha hvilken som helst animalsk opprinnelse inkludert fugler og pattedyr. Det foretrekkes at antistoffene er antistoffer fra menneske, mus/rotte, esel, hare, geit, marsvin, kamel, lama, hest eller kyllinger. I et annet tilfelle kan det variable området stamme fra bruskfisker (f.eks. fra hai). Slik som brukt i dette dokumentet, inkluderer «humane» antistoffer med aminosyresekvens fra et humant immunoglobulin og inkluderer antistoffer isolert fra humane immunoglobulinbiblioteker eller fra dyr som er transgeniske for ett eller flere humane immunoglobuliner og som ikke uttrykker endogene immunoglobuliner som beskrevet nedenfor og f.eks. i det amerikanske patentnr. 5,939,598 av Kucherlapati et al. [0062] Slik som brukt i dette dokumentet, inkluderer begrepet «tungkjededel» aminosyresekvenser som er avledet fra en immunoglobulin tungkjede. Et polypeptid som omfatter en tungkjededel omfatter minst ett av: Et C H 1-domene, et hengsel (f.eks. øvre, midtre og/eller nedre hengselområde)-
.")
12 domene, et C H 2-domene, et C H 3-domene eller variant eller fragment av det. En binding kan f.eks. omfatte en polypeptidkjede som omfatter et C H 1-domene, polypeptidkjede som omfatter et C H 1- domene, minst en del av et hengseldomene og et C H 2-domene, polypeptidkjede som omfatter et C H 1- domene og et C H 3-domene, en polypeptidkjede som omfatter et C H 1-domene, minst en del av et hengseldomene og et C H 3-domene eller en polypeptidkjede som omfatter et C H 1-domene, minst en del av et hengseldomene, et C H 2-domene og et C H 3-domene. I et annet tilfelle omfatter polypeptider til offentliggjøringen, et polypeptidkjede som omfatter et C H 3-domene. I tillegg kan et polypeptid til bruk i offentliggjøringen, mangle minst en del av et C H 2-domene (f.eks. hele eller en del av et C H 2- domene). Slik som beskrevet ovenfor, vil det være forstått av en person med vanlige ferdigheter i faget at disse domene (f.eks. de tunge kjededelene) kan modifiseres slik at de varierer i aminosyresekvensen fra det naturlig forekommende immunoglobulinmolekylet. [0063] I visse Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledning av disse og som offentliggjøres i dette dokumentet, er de tunge kjededelene av én polypeptidkjede til en multimer identiske med dem på en andre polypeptidkjede til multimeren. Som et alternativ er den tunge kjededelen som inneholder monomerer til offentliggjøringen, ikke identiske. Hver monomer kan f.eks. omfatte et annet målbindingssted, og f.eks. danne et bispesifikt antistoff. [0064] De tunge kjededelene til et bindingspolypeptid som brukes ved diagnostiske og behandlingsmetoder som offentliggjøres i dette dokumentet, kan avledes fra forskjellige immunoglobulinmolekyler. En tungkjededel til et polypeptid kan f.eks. omfatte et C H 1-domene avledet fra et IgG1-molekyl og et hengselsområde avledet fra et IgG3-molekyl. I et annet eksempel kan en tungkjededel omfatte et hengselområde som er delvis avledet fra et IgG1-molekyl og delvis fra et IgG3-molekyl. I et annet eksempel kan en tungkjededel omfatte et kimærisk hengsel som er delvis avledet fra et IgG1-molekyl og delvis fra et IgG4-molekyl. [0065] Slik som brukt i dette dokumentet, inkluderer begrepet «lettkjededel» aminosyresekvenser som er avledet fra en immunoglobulin lettkjede. Det foretrekkes at den lette kjededelen omfatter minst én av et V L - eller C L domene. [0066] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse og som offentliggjøres i dette dokumentet eller spesifisert i som epitop(er) eller del(er) til et antigen, f.eks. et målpolypeptid (Sp35) som de gjenkjenner eller bindes spesifikt. Delen av en målpolypeptid som spesifikt samhandler med antigenbindingsdomenet til et antistoff, er en «epitop» eller «antigent determinant». Et målpolypeptid kan omfatte en enkel epitop, men vanligvis omfatter det minst to epitoper og kan inkludere et hvilket som helst antall epitoper avhengig av størrelsen, formen og typen antigen. I tillegg gjøres det oppmerksom på at en «epitop» på et målpolypeptid kan være eller inkludere ikke-polypeptide elementer, f.eks. kan en epitop inneholde en karbohydrat-sidekjede. [0067] Minste størrelsen til et peptid eller en polypeptidepitop til et antistoff anslås til å være fire til fem aminosyrer. Det foretrekkes at peptid- eller polypeptidepitoper inneholder minst sju, og det foretrekkes enda mer at de inneholder ni eller aller helst foretrekkes minst omtrent 15 til omtrent 30 aminosyrer. Fordi en CDR kan gjenkjenne et antigent peptid eller polypeptid i dets tredje form, trenger ikke aminosyrer som omfatter en epitop, å være sammenhengende, og i noen tilfeller er det

13 mulig at offentliggjøringen ikke engang er i samme peptidkjede. I den gjeldende offentliggjøringen inneholder peptid- eller polypeptidepitopen som gjenkjenner Sp35-antistoffer i den gjeldende offentliggjøringen, en sekvens på minst 4, på minst 5, på minst 6, på minst 7, på minst 8 foretrekkes mer, på minst 9, på minst 10, på minst 15, på minst 20, på minst 25 eller mellom omtrent 15 og omtrent 30 sammenhengende eller ikke-sammenhengende Sp35-aminosyrer. [0068] Generelt betyr «spesifikt bindes» at et antistoff bindes til en epitop via dens antigenbindingsdomene og at bindingen innbefatter noen komplementaritet mellom antigenbindingsdomenet og epitopen. I henhold til denne definisjonen sies det at antistoffet «spesifikt bindes» til en epitop når den bindes til den epitopen via dens antigenbindingsdomene lettere enn den ville bindes til en tilfeldig, urelatert epitop. Begrepet «spesifisitet» brukes i dette dokumentet for å kvalifisere den relative affiniteten som gjør at et visst antistoff bindes til en viss epitop. Det kan f.eks. regnes med at antistoff «A» har større spesifisitet for en bestemt epitop enn antistoff «B» eller at det blir sagt at antistoff «A» bindes til epitop «C» med større spesifisitet enn den relaterte epitopen «D». [0069] «Fortrinnsvis bindes» betyr at antistoffet bindes spesifikt til en epitop lettere enn det ville bindes til en relatert, lignende, homolog eller analog epitop. Derfor vil et antistoff som «fortrinnsvis bindes» til en bestemt epitop mer sannsynlig bindes til den epitopen enn en relatert epitop, selv om et slikt antistoff kan kryssreagere med den relaterte epitopen. [0070] Som ikke begrensende eksempel, kan et antistoff regnes som fortrinnsvis bundet til en første epitop hvis det binder den første epitopen med en dissosiasjonskonstant (K D ) som er mindre enn antistoffets K D ved den andre epitopen. I et annet ikke begrensende eksempel, kan et antistoff regnes som fortrinnsvis bundet til et første antigen hvis det binder den første epitopen med en affinitet som er minst én størrelsesorden i forhold til antistoffets K D ved den andre epitopen. I et annet ikke begrensende eksempel, kan et antistoff regnes som fortrinnsvis bundet til en første epitop hvis det binder den første epitopen med en affinitet som er minst to størrelsesordener i forhold til antistoffets K D ved den andre epitopen. [0071] I et annet ikke begrensende eksempel, kan et antistoff regnes som fortrinnsvis bundet til en første epitop hvis det binder den første epitopen med en av-rate (k(off)) som er mindre enn antistoffets k(off) ved den andre epitopen. I et annet ikke begrensende eksempel, kan et antistoff regnes som fortrinnsvis bundet til en første epitop hvis det binder den første epitopen med en affinitet som er minst én størrelsesorden i forhold til antistoffets k(off) ved den andre epitopen. I et annet ikke begrensende eksempel, kan et antistoff regnes som fortrinnsvis bundet til en første epitop hvis det binder den første epitopen med en affinitet som er minst to størrelsesordener i forhold til antistoffets k(off) ved den andre epitopen. [0072] Det kan sies at et antistoff eller antigenbindende fragment, variant eller avledning som offentliggjøres i dette dokumentet, binder et målpolypeptid som offentliggjøres i dette dokumentet eller et fragment eller variant av det med en av-rate (k(off)) som er mindre eller lik 5 X 10-2 sek -1, 10-2 sek -1, 5 X 10-3 sek -1 eller 10-3 sek -1. Det foretrekkes mer et antistoff av offentliggjøringen binder et målpolypeptid som offentliggjøres i dette dokumentet eller et fragment eller variant av det med en

14 av-rate (k(off)) som er mindre eller lik 5 X 10-4 sek -1, 10-4 sek -1, 5 X 10-5 sek -1 eller 10-5 sek -1 5 X 10-6 sek sek -1, 5 X 10-7 sek -1 eller 10-7 sek -1. [0073] Det kan sies at et antistoff eller antigenbindende fragment, variant eller avledning som offentliggjøres i dette dokumentet, binder et målpolypeptid som offentliggjøres i dette dokumentet eller et fragment eller variant av det som en på-rate (k(on)) som er større eller lik 10 3 M -1 sek -1, 5 X 10 3 M -1 sek -1, 10 4 M -1 sek -1 eller 5 X 10 4 M -1 sek -1. Det foretrekkes mer at et antistoff av offentliggjøringen som binder et målpolypeptid som offentliggjøres i dette dokumentet eller fragment eller variant av det med en på-rate (k(on)) som er større enn eller lik 10 5 M -1 sek -1, 5 X 10 5 M -1 sek -1, 10 6 M -1 sek -1 eller 5 X 10 6 M -1 sek -1 eller 10 7 M -1 sek -1. [0074] Det sies at et antistoff er kompetitivt hemmingsbindende av et referanse-antistoff til en bestemt epitop hvis den fortrinnsvis bindes til denne epitopen i den utstrekning som den blokkeres i noen grad og binder referanse-antistoffet til epitopen. Kompetitiv hemming kan bestemmes ved bruk av en hvilken som helst metode som er kjent innen faget, f.eks. kompetitive ELISA-analyser. Det kan sies at et antistoff er kompetitivt hemmingsbindende av referanse-antistoffet til en bestemt epitop med minst 90 %, med minst 80 %, med minst 70 %, men minst 60 % eller med minst 50 %. [0075] Slik som brukt i dette dokumentet, henviser begrepet «affinitet» til en styrkemåling av bindingen til en enkel epitop med CDR til et immunoglobulinmolekyl. Se f.eks. Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, ) pp Slik som brukt i dette dokumentet henviser begrepet «aviditet» til den samlede stabiliteten til komplekset mellom en gruppe med immunoglobuliner og et antigen, dvs. den samlede funksjonelle styrken til en immunoglobulinblanding med antigenet. Se f.eks. Harlow på side Aviditet relaterer seg både til affiniteten til det enkelte immunoglobulinmolekylet i gruppen med spesifikke epitoper og også valensene til immunoglobulinene og antigenet. F.eks. ville interaksjonen mellom et bivalent monoklonalt antistoff og et antigen med høyt gjentakende epitopstruktur, som f.eks. et polymer, ha høy aviditet. [0076] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse kan også beskrives eller spesifiseres i forhold til deres kryssreaktivitet. Slik som brukt i dette dokumentet, henviser begrepet «kryssreaktivitet» til evnen til et antistoff, spesifikk for ett antigen, å reagere med et annet antigen, en måling av relasjonen mellom to forskjellige antigenstoffer. Dermed er et antistoff kryssreaktivt hvis det bindes til en epitop annet enn det som fremkalte formasjonen. Den kryssreaktive epitopen inneholder generelt mange av de samme komplementære strukturelle egenskapene som den induserte epitopen og i noen tilfeller kan denne faktisk passe bedre enn originalen. [0077] Visse antistoffer har f.eks. en viss grad av kryssreaktivitet i det de binder relaterte, men ikke identiske epitoper, f.eks. epitoper med minst 95 %, minst 90 %, minst 85 %, minst 80 %, minst 75 %, minst 70 %, minst 65 %, minst 60 %, minst 55 % og minst 50 % identiske (som beregnet med metoder som er kjent innen faget og som beskrives i dette dokumentet) til referanse-epitopen. Det kan sies at et antistoff har liten eller ingen kryssreaktivitet hvis det ikke binder epitoper med mindre enn 95 %, mindre enn 90 %, mindre enn 85 %, mindre enn 80 %, mindre enn 75 %, mindre enn 70 %, mindre enn 65 %, mindre enn 60 %, mindre enn 55 % og mindre enn 50 % identiske (som beregnet med metoder
) som er større")
15 som er kjent innen faget og beskrevet i dette dokumentet) til en referanse-epitop. Et antistoff kan regnes som «svært spesifikk» for en viss epitop hvis det ikke binder noen annen analog, ortolog eller homolog til den epitopen. [0078] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse kan også beskrives eller spesifiseres i forhold til deres affinitet til et polypeptid. Foretrukne bindingsaffiniteter inkluderer dem med en dissosiasjonskonstant eller Kd som er mindre enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 X 10-5 M, 10-5 M, 5 x 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X 10-9 M, 10-9 M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M eller M. [0079] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledning av dem, kan være «multispesifikke», f.eks. bispesifikke, trispesifikke eller ha større multispesifisitet som betyr at den gjenkjenner og binder to eller flere forskjellige epitoper som finnes på ett eller flere forskjellige antigener (f.eks. proteiner) samtidig. Dermed, om et Sp35-antistoff er «monospesifikt» eller «multispesifikt», f.eks. «bispesifikt», henviser det til antall forskjellige epitoper som et bindende polypeptid reagerer med. Multispesifikke antistoffer kan være spesifikke for forskjellige epitoper til et målpolypeptid som beskrives i dette dokumentet eller kan være spesifikk for et målpolypeptid i tillegg til et heterolog epitop som f.eks. et heterologt polypeptid eller fast støttemateriale. [0080] Slik som brukt i dette dokumentet, henviser begrepet «valens» til antall mulige bindende domener, f.eks. antigenbindende domener som finnes i et Sp35-antistoff, bindende polypeptid eller antistoff. Hvert bindende domene binder spesifikt én epitop. Når et Sp35-antistoff, bindende polypeptid eller antistoff omfatter mer enn ett bindende domene, kan hvert bindende domene spesifikt binde samme epitop, til et antistoff med to bindende domener som kalles «bivalent monospesifikt» eller to forskjellige epitoper til et antistoff med to bindende domener som kalles «bivalent bispesifikt». Et antistoff kan være bispesifikt og bivalent for hver spesifisitet (kalles «bispesifikke tetravalente antistoffer»). I et annet tilfelle, kan det lages tetravalente ministoffer eller domeneslettede antistoffer. [0081] Bispesifikke bivalente antistoffer og metoder for å lage dem beskrives f.eks. i de amerikanske patentnr. 5,731,168, 5,807,706, 5,821,333, og de amerikanske søknadspublikasjonsnr. 2003/ og 2002/ , der offentliggjøringen av alle disse innlemmes i dette dokumentet ved henvisning. Bispesifikke tetravalente antistoffer og metoder for å lage dem, beskrives f.eks. i WO 02/ og WO 00/ Se generelt, PCT-publikasjon WO 93/17715, WO 92/08802, WO 91/00360, WO 92/05793, Tutt et al., J. Immunol. 147:60-69 (1991), amerikanske patentnr. 4,474,893, 4,714,681, 4,925,648, 5,573,920, 5,601,819, Kostelny et al., J. Immunol. 148: (1992). [0082] Som tidligere indikert, er subenhetsstrukturene og tredimensjonal konfigurasjon av konstantområdene til de forskjellige immunoglobulinklassene godt kjent. Slik som brukt i dette dokumentet, inkluderer begrepet «V H -domene» det aminoterminale variable domenet til en tung immunoglobulinkjede og begrepet «C H 1-domene» inkluderer det første (mest aminoterminal) konstantområde-domene til et immunoglobulin med tungkjede. C H 1-domenet er ved siden av V H - domenet og er aminoterminalt til hengselsområdet til et immunoglobulin-tungkjedemolekyl.
![[0078] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse kan også beskrives eller spesifiseres i forhold til deres affinitet til et polypeptid.](/docs-images/56/5927828/images/page_15.jpg "Foretrukne bindingsaffiniteter inkluderer dem med en dissosiasjonskonstant eller Kd som er mindre enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 X 10-5 M, 10-5 M, 5 x 10-6 M, 10-6")
16 [0083] Slik som brukt i dette dokumentet, inkluderer begrepet «C H 2-domene» den delen av en tungkjedemolekyl som strekker seg, f.eks. fra omtrent rest 244 til rest 360 til et antistoff, ved bruk av konvensjonelle nummereringsplaner (restene 244 til 360, Kabat-nummereringsystem og restene , EU-nummereringssystem. Se Kabat EA et al. op. cit. C H 2-domenet er unikt i og med at det er ikke tett paret med et annet domene. I stedet er to N-koplede karbohydratkjeder med grener innkoplet mellom to C H 2-domener til et intakt nativt IgG-molekyl. Det er også godt dokumentert at C H 3- domenet strekker seg fra C H 2-domenet til C-terminalen til IgG-molekylet og omfatter omtrent 108 rester. [0084] Slik som brukt i dette dokumentet, inkluderer begrepet «hengselsområde» den delen av et tungkjedemolekyl som kopler C H 1-domenet til C H 2 domenet. Dette hengselsområdet omfatter omtrent 25 rester og er fleksibelt, noe som gjøre at de to N-terminale antigenbindingsområdene kan bevege seg uavhengig av hverandre. Hengselsområdene kan deles inn i tre atskilte domener: øvre, midtre og nedre hengselsdomene (Roux et al., J Immunol. 161:4083 (1998)). [0085] Slik som brukt i dette dokumentet, inkluderer begrepet «disulfidbinding» den kovalente bindingen som dannes mellom to svovelatomer. Aminosyrecysteinet omfatter en tiolgruppe som kan danne en disulfidbinding eller bro med en andre tiolgruppe. I de fleste naturlig forekommende IgGmolekyler, er C H 1- og C L -område koplet sammen med en disulfidbinding og de to tunge kjedene er koplet sammen med to disulfide bindinger som er posisjonert tilsvarende 239 og 242 ved bruk av Kabat-nummereringssystemet (posisjon 226 eller 229, EU-nummereringssystemet). [0086] Slik som brukt i dette dokumentet, betyr begrepet «kimærisk antistoff» et hvilket som helst antistoff der det immunoreaktive området eller stedet skaffes eller avledes fra en første art og det konstante området (som kan være intakt, partielt eller modifisert iht. offentliggjøringen skaffes fra en andre art. I et foretrukket tilfelle vil målbindingsområdet eller -stedet komme fra en ikke-human kilde (f.eks. mus eller primat), og det konstante området er humant. [0087] Slik som brukt i dette dokumentet, henviser begrepet «konstruert antistoff» til et antistoff der det variable domenet i enten den tunge eller lette kjeden eller begge endres med minst en delvis erstatning av ett eller flere CDR-er fra et antistoff med kjent spesifisitet og om nødvendig, med delvis erstatning av rammeverksområdet og sekvensutbytting. Selv om CDR-ene kan være avledet fra et antistoff i samme klasse eller til og med samme underklasse som antistoffet som rammeverksområdene er avledet fra, regnes det med at CDR-ene vil bli avledet fra et antistoff i en annen klasse og helst fra et antistoff fra en annen art. Et konstruert antistoff der én eller flere «donorer»-cdr-er fra et ikke-humant antistoff med kjent spesifisitet podes inn et humant rammeverksområde med tung- eller lettkjede, kalles i dette dokumentet et «humanisert antistoff». Der er muligens ikke nødvendig å erstatte alle CDR-ene med de komplette CDR-ene fra det donorvariable området for å overføre den antigenbindende kapasiteten til et variabelt domene til et annet. I stedet kan det muligens bare være nødvendig å overføre restene som trengs for å opprettholde aktiviteten til målbindingsstedet. Med forklaringene som finnes i f.eks. de amerikanske patentnr. 5,585,089, 5,693,761, 5,693,762 og 6,180,370 vil det være godt innenfor kompetansen til personer med ferdigheter i faget, enten for å utføre rutinemessige forsøk eller testing med prøving og feiling for å skaffe et funksjonelt konstruert eller humanisert antistoff.

17 [0088] Slik som brukt i dette dokumentet, inkluderer begrepet «riktig foldet polypeptid» (f.eks. Sp35- antistoffer) der alle de funksjonelle domene som omfatter polypeptidet, er hver for seg aktive. Slik som brukt i dette dokumentet, inkluderer begrepet «uriktig foldet polypeptid» polypeptider der minst ett av de funksjonelle domenene til polypeptidet ikke er aktivt. I ett tilfelle omfatter et riktig fold polypeptid polypeptidkjeder som er koplet sammen med minst én disulfidbinding og omvendt, en uriktig foldet polypeptid omfatter polypeptidkjeder som ikke er koplet sammen med minst én disulfidbinding. [0089] Slik som brukt i dette dokumentet, inkluderer begrepet «konstruert» manipuleringer av nukleinsyre eller polypeptidmolekyler på syntetisk måte (f.eks. rekombinante metoder, in vitro peptidsyntese, enzymatisk eller kjemisk kopling av peptider eller en kombinasjon av disse metodene). [0090] Slik som brukt i dette dokumentet, brukes begrepene «koplet», «fusjonert» eller «fusjon» om hverandre. Disse begrepene henviser til sammenkoplingen av to eller flere elementer eller komponenter med en hvilken som helst metode inkludert kjemisk forbindelse eller rekombinante metoder. En «fusjon i rammen» henviser til en sammenkopling av to eller flere polynukleotide ORF-er (open reading frames) for å danne en kontinuerlig lengre ORF som opprettholder riktig translasjonsavlesingsramme til de opprinnelige ORF-ene. Dermed er et rekombinant fusjonsprotein et enkelt protein som inneholder to eller flere segmenter som korresponderer med polypeptidene som er kodet av de opprinnelige ORF-ene (som segmentene ikke naturlig er koplet til). Selv om avlesningsrammen er laget kontinuerlig gjennom hele de fusjonerte segmentene, kan segmentene være fysisk eller spatialt atskilt med f.eks. en tilkoplingssekvens i rammen. Polynukleotidene som koder CDR-ene til et immunoglobulinvariabelt område, kan f.eks. være fusjonert, i rammen, men være atskilt med en polynukleotidkode i minste ett immunoglobulin-rammeverksområde eller flere CDR-områder i tillegg, så lenge som de «fusjonerte» CDR-ene er samtranslatert som en del av et kontinuerlig polypeptid. [0091] I kontekst i forbindelse med polypeptider er en «lineær sekvens» eller en «sekvens» en rekkefølge med aminosyrer i et polypeptid i en amino-til-karboksylterminal retning der naborester i sekvensen er sammenhengende i den primære strukturen til polypeptidet. [0092] Slik som brukt i dette dokumentet, henviser begrepet «uttrykk» til en prosess der et gen produserer et biokjemikalie som f.eks. RNA eller polypeptid. Prosessen inkluderer en hvilken som helst manifestasjon av den funksjonelle tilstedeværelsen av genet inni cellen inkludert uten begrensing, gen-«knockdown» samt både som transient uttrykk og stabilt uttrykk. Det inkluderer uten begrensing, transkripsjon av genet inni mrna (messenger RNA), trna (transfer RNA), shrna (small hairpin RNA), sirna (small interfering RNA) eller et hvilket som helst annet RNA-produkt og omgjøring av en slik RNA til polypeptid(er). Hvis sluttproduktet som ønskes, er et biokjemikalie, inkluderer uttrykket dannelsen av det biokjemikalie og eventuelle forstadier. Uttrykk av et gen produserer et «genprodukt». Slik som brukt i dette dokumentet, kan et genprodukt være enten en nukleinsyre, f.eks. en budbringer/rna produsert ved transkripsjon av et gen eller et polypeptid som er overført fra et transkript. I tillegg inkluderer genprodukter som beskrives i dette dokumentet, nukleinsyrer med transkriptale modifikasjoner, f.eks. polyadenylering eller polypeptider med

18 postoverføringsmodifikasjoner, f.eks. metylering, glykosylering, tilsetting av lipider, forbindelse med andre protein-subenheter, proteolytisk spalting og lignende. [0093] Slik som brukt i dette dokumentet, henviser begrepene «behandle» eller «behandling» til både terapeutisk behandling og profylaktiske eller forebyggende tiltak der målet er å hindre eller sinke (minke) en uønsket fysiologisk endring eller lidelse som f.eks. progresjonen av multippel sklerose. Fordelaktige eller ønskede kliniske resultater inkluderer, men er ikke begrenset til lindring av symptomer, minskning av utbredelsen av sykdommen, stabilisering av (dvs. ikke forverring) sykdomstilstand, forsinket eller avtagende sykdomsprogresjon, forbedring eller lindring av sykdomstilstanden og tilbakegang (enten delvis eller total) om det er påviselig eller ikke. «Behandling» kan også bety forlenget overlevelse sammenlignet med forventet overlevelse uten å motta behandling. Personer som trenger behandling, inkluderer dem som allerede har en tilstand eller lidelse samt dem som er tilbøyelige for å ha tilstanden eller lidelsen eller personer der tilstanden eller lidelsen skal hindres. [0094] «Forsøksobjekt» eller «individ» eller «dyr» eller «pasient» eller «pattedyr» betyr et hvilket som forsøksobjekt, spesielt et pattedyrobjekt som man ønsker å stille diagnose eller prognose eller ønsker å behandle: Pattedyrobjekter inkluderer mennesker, husdyr og dyr i en dyrepark, dyr som brukes innen sport eller kjæledyr som f.eks. hunder, katter, marsvin, kaniner, rotter, mus, hester, kveg, kyr, osv. [0095] Slik som brukt i dette dokumentet, inkluderer slike fraser som «et forsøksobjekt som ville dra nytte av tilførsel av et Sp35-antistoff» eller «et dyr som trenger behandling» forsøksobjekter som f.eks. pattedyrobjekter som ville dra nytte av tilførsel av et Sp35-antistoff som f.eks. brukes for å detektere et Sp35-polypeptid (f.eks. ved diagnostisering) og/eller av behandling, dvs. lindring eller forebyggelse av f.eks. MS med et Sp35-antistoff. Slik som beskrevet i mer detalj i dette dokumentet, kan Sp35-antistoffet brukes i ukonjugert form, f.eks. et medikament, prekursor eller en isotop. II. Sp [0096] Naturlig forekommende humant Sp35 (Sp35) er et glykosylert sentral nervesystem spesifikt protein som anslås at ha 614 aminosyrer (SEQ ID NR: 2), inkluderer en 33 aminosyre-signalsekvens. Sp35 er også kjent innen faget under navnene LINGO-1, LRRN6, LRRN6A, FLJ14594, LERN1, MGC17422 og UNQ201. Det humane, villtype Sp35-polypeptidet med full lengde, inneholder et LRR-domene som består av 14 leucinrike gjentakelser (inkludert N- og C-terminale hetter), et Ig-domene, et transmembrant område og et cytoplasmisk domene. Det cytoplasmiske domenet inneholder et kanonikalt tyrosinfosforyleringssted. I tillegg inneholder det naturlig forekommende Sp35-proteinet en signalsekvens, et kort grunnleggende området mellom LRRCT og Ig-domenet, et transmembrant område mellom Ig-domenet og det cytoplasmiske domenet. Det humane Sp35-genet (SEQ ID NR:1) inneholder alternative translasjonsstartkondoner, slik at seks ekstra aminosyrer, dvs. MQVSKR (SEQ ID NR: 3) finnes eller ikke finnes ved N-terminus til Sp35-signalsekvensen. Tabell 2 inneholder Sp35- domene og andre områder iht. aminosyre-restnummeret, basert på Sp35-aminosyresekvensen som

19 18 gjengis som SEQ ID NR: 2. Sp35-polypeptidet gjengis i mer detalj i PCT-publikasjonsnr. WO 2004/ TABELL 2--Sp35-domener Domene eller region Rest på begynnelse Rest på slutten Signalsekvens 1 33 eller 35 LRRNT 34 eller LRR LRR LRR LRR LRR LRR LRR LRR LRR LRR LRR LRR LRRCT eller 416 Grunnleggende 415 eller Ig Koplende sekvens Transmembrant Cytoplasmisk [0097] Vevsfordeling og utviklingsuttrykk av Sp35 er studert hos mennesker og rotter. Sp35-biologi er studert i en forskningsdyremodell (rotter). Sp35-rotteuttrykk er lokalisert i nevroner og oligodendrocytter som bestemt med Northern blot og immunohistokjemisk farging. Nivået ved Sp35 mrna-rotteuttrykk reguleres utviklende, med topp rett etter fødsel, dvs. omtrent postnatalt dag én. I en skademodell med tverrsnitt av rotteryggmargen, oppreguleres Sp35 ved skadestedet som bestemt med RT-PCR. Se Mi et al. Nature Neurosci. 7: (2004). [0098] I konteksten til aminosyrene som omfatter forskjellige strukturelle og funksjonelle domener av et Sp35-polypeptid, inkluderer begrepet «omtrent» oppgitte verdier og verdier som er større eller mindre med flere (f.eks. 10,9,8, 7, 6, 5,4,3,2 eller 1) aminosyrer. Fordi posisjonen til disse domenene som står i tabell 1, er anslått med datagrafikk, vil en person med vanlige ferdigheter i faget forstå på at aminosyrerestene som utgjør domenene, kan variere litt (f.eks. med omtrent 1 til 15 rester) avhengig av kriteriene som brukes for å definere domenet.
.")

20 19 5 [0099) Oppfinnerne har oppdaget at villtype Sp35 med full lengde bindes til NgR1. Se PCTpublikasjonsnr. WO 2004/ Oppfinnerne har også oppdaget at Sp35 uttrykkes i oligodendrocytter og at Sp35-proteinet er involvert i regulering av oligodendrocyttmedierte myelineringer av aksoner. Se det amerikanske patentpublikasjonsnr. 2006/ A1. [0100] Nukleotidsekvensen til Sp35-molekylet med full lengde, er følgende: NR:1). [0101] Polypeptidsekvensen til Sp35-polypeptidet med full lengde, er følgende: 10 SEQ ID NR:2). III. Sp35-ANTISTOFFER
.")
21 [0102] I ett tilfelle rettes den gjeldende offentliggjøringen mot Sp35-antistoffer eller antigenbinding, fragmenter, varianter eller avledning av dem. Den gjeldende offentliggjøringen inkluderer f.eks. i minste fall de antigenbindende domenene til visse monoklonale antistoffer og fragmenter, varianter og avledninger av dem som vises i tabell 3A til 3E. [0103] Tabell 3A beskriver området til Sp35-polypeptidet som er bundet av visse fagbibliotekavledede antistoffer med full lengde. Disse antistoffene har samme variable områder som Fab-fragmentene som er avledet fra fagvisningsbiblioteket 1 som vist i tabell 3B (D05 i tabellen har f.eks. samme variabelt område som Li05 i tabell 3B, D06 i tabell 3A har samme variabelt område som Li06 i tabell 3B. osv.). Antistoffene ble testet for binding av Sp35-fragmenter som definert i tabell 3A ved bruk av metoder som er godt kjent innen faget. [0104] Tabell 3B-3E beskriver evnen til de angitt monoklonale antistoffene eller Fab-fragmentene til å påvise Sp35 i forskjellige analyser som f.eks.: FACS (Fluorescent Activated Cell Sorting), IP (Immunoprecipitation), Western blot-analyse, IHC (Immunohistochemistry) til ELISA (Enzyme Linked hnmunosorbent Assay). Detaljerte protokoller for å utføre disse analysene beskrives i dette dokumentet eller er godt kjent og forstått av personer med ferdigheter i faget. Hybridomavledede monoklonale antistoffer som står oppført i tabell 3B og 3C, ble produsert med injeksjon av oppløselig Sp35 og deretter isolert ved bruk av hybridomteknologi som er godt kjent innen farget og som beskrives i dette dokumentet. Monoklonale antistoffer og antistoff Fab-fragmenter som står oppført i tabell 3B, ble isolert fra to forskjellige fagvisningsbiblioteker ved bruk av metoder som er kjent innen faget. Sp35- fragment rotte-fc rotte-fc AP-Sp (1-419) AP-Sp ( ) human- Fc human- Fc human- Fc D03 (Li03 variabelt område) D05 (Li05 variabelt område) D06 (Li06 variabelt område) TABELL 3A D08 (Li08 variabelt område) D11 (Li03 variabelt område) D13 (Li13 variabelt område) D33 (Li33 variabelt område) /- +/ Ikke bestemt Ikke bestemt + +/- +/- Ikke bestemt Ikke bestemt Ikke bestemt Ikke bestemt Ikke bestemt Ikke bestemt
22 TABELL 3B - SP35 MONOKLONALE ANTISTOFFER HYDRIDOMAVLEDEDE MONOKLONALE ANTISTOFFER Immonopresipitasjon ja ja nei nei nei nei nei nei ja mus / rotte sp35 Nei (mus og rotte) Nei (mus og rotte) Nei (mus og rotte) Nei (mus og rotte) Nei (mus og rotte) Nei (mus og rotte) Nei (mus og rotte) ja med overuttrykt msp35 NO/EP human- Fc
23 TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.) ++ (bindes til 293 celler) ++ (bindes til 293 celler) +++ (øvre bånd) +++ (øvre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (øvre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (nedre bånd) +++ (nedre bånd) Ja Nei Nei Nei Nei Ja Ja Ja Nei Ja Ja (mus) Nei (mus) Nei (mus) Nei (mus) Nei (mus) Ja (mus) Ja (mus) Ja (mus) Nei (mus) Ja (mus) NO/EP
24 +++ (øvre bånd) +++ (øvre bånd) +++ (øvre bånd) + (nedre bånd) Nei Nei Nei Nei (mus) Nei (mus) Nei (mus) NO/EP TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.) 5
25 HYDRIDOMAVLEDEDE MONOKLONALE ANTISTOFFER (fort.) IHC på Transfekterte IHC på vev celler I/R I/R nei nei Ja med nei bakgrunn Ja med bakgrunn Ja med bakgrunn Ja med bakgrunn nei ja nei nei nei nei ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja NO/EP TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.)
26 25 TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.) 5

27 FAGEDISPLAY-BIBLIOTEK-1-AVLEDEDE MONOKLONALE Fab-FRAGMENTER mmonopresipitasjon Western Mus/rotte sp35 NO/EP TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.)

28 FAGEDISPLAY-BIBLIOTEK-1-AVLEDEDE MONOKLONALE Fab-FRAGMENTER IHC på transfekterteceller IHC på vev NO/EP TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.) 5

29 FAGEDISPLAY-BIBLIOTEK-2-AVLEDEDE MONOKLONALE Fab-FRAGMENTER Immunopresipitasjon klebete klebete klebete ja nei svak Mus/rotte sp35 nei no NO/EP TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.) 5
30 FAGEDISPLAY-BIBLIOTEK-2-AVLEDEDE MONOKLONALE Fab-FRAGMENTER nei nei nei veldig svakt nei nei nei nei nei nei svakt nei ja ja ja med høy bakgrunn nei 29 TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.) IHC på transfekterte celler IHC på vev ja med høy bakgrunn ja ja I / R svakt nei nei nei ja ja nei nei nei nei nei veldig svakt bare farge noen få celler nei ja med høy bakgrunn ja med høy bakgrunn veldig svakt nei I / R I / R ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja ja NO/EP
31 Fagedisplay-bibliotek-1 Avledede komplette monoklonale antistoffer Immunopresipitasjon Fagedisplay-bibliotek-1 Avledede komplette monoklonale antistoffer IHC på transfekterte celler IHC på vev Mus/rotte sp35 NO/EP TABELL 3B - SP35 MONOKLONALE ANTISTOFFER (fort.) Nøkkel: husp35 - humant Sp35-protein msp35 mus Sp35-protein WT vill type KO - «knockout» IHC immunohistokjemi FACS Fluorescent aktivert cellesortering 5
32 Antistoff Art murin murin Rottehjernehomogenat ja NO/EP TABELL 3C HYBRIDOMAVLEDEDE SP35 MONOKLONALE ANTISTOFFER 5

33 32 TABELL 3D Antistoff Art murin murin murin murin murin murin murin murin murin murin murin murin 5
 Antistoff Art FACS på 293 celler FACS på stabil CHO murin 0,7 nm murin murin murin murin murin murin murin murin")
34 33 TABELL 3D (fort.) Antistoff Art FACS på 293 celler FACS på stabil CHO murin 0,7 nm murin murin murin murin murin murin murin murin murin c c c c (10,4) nm 3,7 nm (2,7 nm 5,4 nm 5 murin murin <0,4 nm 0,4 nm 5,1 nm 4,4 nm 4,6 nm 6 nm

35 34 TABELL 3D (fort.) Art 5
36 35 TABELL 3D (fort.) Art
 22,8 nm passer ikke 5,5 nm 2,32 nm 26,6 nm 3,3 nm 18,6 nm 0,49 nm 0,74 nm,")
37 36 TABELL 3D (fort.) Art 0,25 nm 0,27 nm 5 Art TABELL 3D (fort.) 22,8 nm passer ikke 5,5 nm 2,32 nm 26,6 nm 3,3 nm 18,6 nm 0,49 nm 0,74 nm, 3,2 (CG) 24,7 nm
 0,261")
38 37 Art 1,39 nm, 4 (CG) passer ikke 0,208 nm for igg2 7,6 nm (CG) 0,261 nm
39 38 Art 0,12 nm 0,156 nm 5

40 39 TABELL 3E Art
41 40 Art 5
42 41 Art (øvre bånd) 5
43 42 Art 5

44 43 TABELL 3E (fort.) Art 5
45 44 Art
46 45 Art 5
![46 TABELL 3E (fort.) Art 5 10 [0105] Slik som brukt i dette dokumentet, inkluderer begrepet «antigenbindende domene» et sted som spesifikt binder en epitop på et antigen (f.eks. en Sp35-epitop).](/docs-images/25/5927828/images/47-0.png "Det antigenbindende domenet til et antistoff inkluderer vanligvis en del av et variabelt område til et tungkjede-immunoglobulin og minst et variabelt område til et lettkjede-immunoglobulin.")
47 46 TABELL 3E (fort.) Art 5 10 [0105] Slik som brukt i dette dokumentet, inkluderer begrepet «antigenbindende domene» et sted som spesifikt binder en epitop på et antigen (f.eks. en Sp35-epitop). Det antigenbindende domenet til et antistoff inkluderer vanligvis en del av et variabelt område til et tungkjede-immunoglobulin og minst et variabelt område til et lettkjede-immunoglobulin. Bindingsstedet som dannes av disse variable områdene, bestemmer spesifisiteten til antistoffet. [0106] Den gjeldende offentliggjøringen er mer spesifikt rettet mot et Sp35-antistoff eller antigenbindende fragment, variant eller avledninger av det der Sp35-antistoffet bindes til samme epitop som et monoklonale antistoff som velges fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 15 [0107] I tillegg trekkes offentliggjøringen til et Sp35-antistoff eller antigenbindende fragment, variant eller avledninger av det der Sp35-antistoffet kompetitivt hemmer et monoklonalt antistoff som velges fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 2B10, 2C11, 2F3, 20 Li81 fra binding til Sp35. [0108] Offentliggjøringen trekkes også til et Sp35-antistoff eller antigenbindende fragment, variant eller avledninger av det der Sp35-antistoffet omfatter minst det antigenbindende området til et
![47 monoklonalt antistoff som velges fra gruppen som består av 201', 3A3, 3A6, 1A7, 5 10 15 20 25 30 [0109] 27.](/docs-images/25/5927828/images/48-0.png "desember 2006 ble følgende hybridomer deponert ved American Type Culture Collection (ATCC) i Manassas, VA, USA: 2.P3B5.2 (ATCC-deponeringsbetegnelse PTA-8106), 7.P1D5.1.G9 (ATCCdeponeringsbetegnelse PTA-8107).")
48 47 monoklonalt antistoff som velges fra gruppen som består av 201', 3A3, 3A6, 1A7, [0109] 27. desember 2006 ble følgende hybridomer deponert ved American Type Culture Collection (ATCC) i Manassas, VA, USA: 2.P3B5.2 (ATCC-deponeringsbetegnelse PTA-8106), 7.P1D5.1.G9 (ATCCdeponeringsbetegnelse PTA-8107). Det deponerte hybridomet 2.P3B5.2 produserer det monoklonale antistoffet 3B5.2, som beskrives i dette dokumentet og det deponerte hybridomet 7.P1D5.1.G9 produserer det monoklonale antistoffet 7P1D5.1.G9 som beskrives i dette dokumentet. Hybridomene kan dyrkes iht. metoder som er godt kjent innen faget og beskrives i dette dokumentet. [0110] I visse tilfeller rettes offentliggjøringen mot et antistoff eller antigenbindende fragment, variant eller avledning av det som spesifikt eller fortrinnsvis til et bestemt Sp35-polypeptidfragment eller domene. Slike Sp35-polypeptidfragmenter inkluderer, men er begrenset til et Sp35- polypeptid som omfatter, hovedsakelig består av, eller består av aminosyrene 34 til 532, 34 til 417, 34 til 425, 34 til 493, 66 til 532, 66 til 417, 66 til 426, 66 til 493,66 til 532,417 til 532, 417 til 425 (grunnleggende Sp35-område), 417 til 493, 417 til 532, 419 til 493 (Sp35- Ig-området) eller 425 til 532 i SEQ ID NR:2 eller et Sp35-variantpolypeptid som er minst 70 %, 75 %, 80 %, 85 %, 90 % eller 95 % likt aminosyrene 34 til 532, 34 til 417, 34 til 425, 34 til 493, 66 til 532, 66 til 417, 66 til 426, 66 til 493, 66 til 532, 417 til 532, 417 til 425 (grunnleggende Sp35-område), 417 til 493, 417 til 532, 419 til 493 (Sp35-Igområdet) eller 425 til 532 til SEQ ID NR:2. [0111] Flere Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter og avledninger av dem til den gjeldende bindingen av offentliggjøringen inkluderer, men er ikke begrenset til disse fragmentene som omfatter, hovedsakelig består av, eller består av én eller flere leucinrike gjentakelser (LRR) av Sp35. Slike fragmenter inkluderer f.eks. fragmenter som omfatter, hovedsakelig består av eller består av aminosyrene 66 til 89, 66 til 113, 66 til 137, 90 til 113, 114 til 137, 138 til 161, 162 til 185, 186 til 209, 210 til 233, 234 til 257, 258 til 281, 282 til 305, 306 til 329 eller 330 til 353 av SEQ ID NR:2. Tilsvarende fragmenter av variant Sp35-polypeptid som er minst 70 %, 75 %, 80 %, 85 %, 90 % eller 95 % lik aminosyrene 66 til 89, 66 til 113, 90 til 113, 114 til 137, 138 til 161, 162 til 185, 186 til 209, 210 til 233, 234 til 257, 258 til 281, 282 til 305, 306 til 329 eller 330 til 353 av SEQ ID NR: 2 er også tiltenkt. [0112] Flere Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter og avledninger av dem til den gjeldende bindingen av offentliggjøringen inkluderer, men er ikke begrenset til disse fragmentene som omfatter, hovedsakelig består av, eller består av ett eller flere cysteinrike områder flankerer LRR til Sp35. Slike fragmenter inkluderer f.eks. et fragment som omfatter, hovedsakelig består av eller består av aminosyrene 34 til 64 av SEQ ID NR:2 (det N-
49 terminale LRR-flankerende området (LRRNT)) eller et fragment som omfatter, hovedsakelig består av eller består av aminosyrene 363 til 416 av SEQ ID NR:2 (det C-terminale LRR-flankerende området (LRRCT)), aminosyrer, tilsvarende fragmenter av et variant Sp35-polypeptid som er minst 70 %, 75 %, 80 %, 85 %, 90 % eller 95 % lik aminosyrene 34 til 64 og 363 til 416 av SEQ ID NR:2 er også tiltenkt. [0113] Som kjent i faget bestemmes «sekvensidentiteten» mellom to polypeptider ved å sammenligne aminosyresekvensen med ett polypeptid med sekvensen til et andre polypeptid. Som omtalt i dette dokumentet, kan det bestemmes at et bestemt polypeptid er minst omtrent 70 %, 75 %, 80 %, 85 %, 90 % eller 95 % likt et annet polypeptid ved bruk av metoder og dataprogrammer/programvarer kjent innen faget som f.eks., men ikke begrenset til BESTFIT-programmet (Wisconsin Sequence Analysis Package, Version 8 for Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Madison, WI 53711, USA). BESTFIT bruker den lokale homologe algoritmen til Smith and Waterman, Advances in Applied Mathematics 2: (1981), for å finne det beste homologsegmentet mellom to sekvenser. Når BESTFIT eller et annet sekvensjusteringsprogram brukes til å bestemme om en bestemt sekvens er f.eks. 95 % identisk med en referansesekvens iht. den gjeldende offentliggjøringen, er parametrene selvsagt stilt inn slik at prosentandelen som disse er like, beregnes av hele lengden til referanse-polypeptidsekvensen og gapene i homologien med inntil 5 % av det totale antall aminosyrer som tillates i referansesekvensen. [0114) Andre Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter, eller avledninger av dem til den gjeldende bindingen av offentliggjøringen inkluderer, men er ikke begrenset til fragmentene som omfatter, hovedsakelig består av eller består av aminosyrene 41 til 525 til SEQ ID NR:2, 40 til 526 til SEQ ID NR:2, 39 til 527 til SEQ ID NR:2, 38 til 528 til SEQ ID NR:2, 37 til 529 til SEQ ID NR:2, 36 til 530 til SEQ ID NR:2, 35 til 531 til SEQ ID NR:2, 34 til 531 i SEQ ID NR:2, 46 til 520 til SEQ ID NR:2, 45 til 521 til SEQ ID NR:2, 44 til 522 til SEQ ID NR:2, 43 til 523 til SEQ ID NR:2 og 42 til 524 til SEQ ID NR:2. [0115] Enda andre Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til bindingen av offentliggjøringen inkluderer, men ikke begrenset til fragmenter som omfatter, hovedsakelig består av eller består av aminosyrene 1 til 33 til SEQ ID NR:2, 1 til 35 til SEQ ID NR:2, 34 til 64 til SEQ ID NR:2, 36 til 64 til SEQ ID NR:2, 66 til 89 til SEQ ID NR:2, 90 til 113 til SEQ ID NR:2, 114 til 137 til SEQ ID NR:2, 138 til 161 til SEQ ID NR:2, 162 til 185 til SEQ ID NR:2, 186 til 209 til SEQ ID NR:2, 210 til 233 til SEQ ID NR:2, 234 til 257 til SEQ ID NR:2, 258 til 281 i SEQ ID NR:2, 282 til 305 til SEQ ID NR:2, 306 til 329 til SEQ ID NR:2, 330 til 353 til SEQ ID NR:2, 363 til 416 til SEQ ID NR:2, 417 til 424 til SEQ ID NR:2, 419 til 493 til SEQ ID NR:2 og 494 til 551 til SEQ ID NR:2. [0116] Enda andre Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til bindingen av offentliggjøringen inkluderer, men ikke begrenset til fragmenter som omfatter, hovedsakelig består av eller består av aminosyrene 1 til 33 til SEQ ID NR:2, 1 til 35 til SEQ ID NR:2, 1 til 64 til SEQ ID NR:2, 1 til 89 til SEQ ID NR:2, 1 til 113 til SEQ ID NR:2, 1 til 137 til SEQ ID NR:2, 1 til 161 til SEQ ID NR:2, 1 til 185 til SEQ ID NR:2, 1 til 209 til SEQ ID NR:2, 1 til 233 til SEQ ID NR:2, 1 til 257 til SEQ ID NR:2, 1 til 281 til SEQ ID NR:2, 1 til 305 til SEQ ID NR:2, 1 til
50 til SEQ ID NR:2, 1 til 353 til SEQ ID NR:2, 1 til 416 til SEQ ID NR:2, 1 til 424 til SEQ ID NR:2, 1 til 493 til SEQ ID NR:2, 1 til 551 til SEQ ID NR:2, 1 til 531 til SEQ ID NR:2 og 1 til 532 til SEQ ID NR:2. [0117] Andre Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til bindingen av offentliggjøringen inkluderer, men ikke begrenset til fragmenter som omfatter, hovedsakelig består av eller består av aminosyrene 34 til 64 til SEQ ID NR:2, 34 til 89 til SEQ ID NR:2, 34 til 113 til SEQ ID NR:2, 34 til 137 til SEQ ID NR:2, 34 til 61 til SEQ ID NR:2, 34 til 185 til SEQ ID NR:2, 34 til 209 til SEQ ID NR:2, 34 til 233 til SEQ ID NR:2, 34 til 257 til SEQ ID NR:2, 34 til 281 til SEQ ID NR:2, 34 til 305 til SEQ ID NR:2, 34 til 329 til SEQ ID NR:2, 34 til 353 til SEQ ID NR:2, 34 til 416 til SEQ ID NR:2, 34 til 424 til SEQ ID NR:2, 34 til 493 til SEQ ID NR:2 og 34 til 551 til SEQ ID NR:2. [0118] Enda andre Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til bindingen av offentliggjøringen inkluderer, men ikke begrenset til fragmenter som omfatter, hovedsakelig består av eller består av aminosyrene 34 til 530 of SEQ ID NR:2, 34 til 531 til SEQ ID NR:2, 34 til 532 til SEQ ID NR:2, 34 til 533 til SEQ ID NR:2, 34 til 534 til SEQ ID NR:2, 34 til 535 til SEQ ID NR:2, 34 til 536 til SEQ ID NR:2, 34 til 537 til SEQ ID NR:2, 34 til 538 til SEQ ID NR:2, 34 til 539 til SEQ ID NR:2, 30 til 532 til SEQ ID NR:2, 31 til 532 til SEQ ID NR:2, 32 til 532 til SEQ ID NR:2, 33 til 532 til SEQ ID NR:2, 34 til 532 til SEQ ID NR:2, 35 til 532 til SEQ ID NR:2, 36 til 532 til SEQ ID NR:2, 30 til 531 til SEQ ID NR:2, 31 til 531 til SEQ ID NR:2, 32 til 531 til SEQ ID NR:2, 33 til 531 til SEQ ID NR:2, 34 til 531 til SEQ ID NR:2, 35 til 531 til SEQ ID NR:2 og 36 til 531 til SEQ ID NR:2. [0119] Enda andre Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til bindingen av offentliggjøringen inkluderer, men ikke begrenset til fragmenter som omfatter, hovedsakelig består av eller består av aminosyrene 36 til 64 til SEQ ID NR:2, 36 til 89 til SEQ ID NR:2, 36 til 113 til SEQ ID NR:2, 36 til 137 til SEQ ID NR:2, 36 til 161 til SEQ ID NR:2, 36 til 185 til SEQ ID NR:2, 36 til 209 til SEQ ID NR:2, 36 til 233 til SEQ ID NR:2, 36 til 257 til SEQ ID NR:2, 36 til 281 til SEQ ID NR:2, 36 til 305 til SEQ ID NR:2, 36 til 329 til SEQ ID NR:2, 36 til 353 til SEQ ID NR:2, 36 til 416 til SEQ ID NR:2, 36 til 424 til SEQ ID NR:2, 36 til 493 til SEQ ID NR:2 og 36 til 551 til SEQ ID NR:2. [0120] Andre Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til bindingen av offentliggjøringen inkluderer, men ikke begrenset til fragmenter som omfatter, hovedsakelig består av eller består av aminosyrene 36 til 530 til SEQ ID NR:2, 36 til 531 til SEQ ID NR:2, 36 til 532 til SEQ ID NR:2, 36 til 533 til SEQ ID NR:2, 36 til 534 til SEQ ID NR:2, 36 til 535 til SEQ ID NR:2, 36 til 536 til SEQ ID NR:2, 36 til 537 til SEQ ID NR:2, 36 til 538 til SEQ ID NR:2 og 36 til 539 til SEQ ID NR:2. [0121] Flere Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til bindingen av offentliggjøringen inkluderer, men ikke begrenset til fragmenter som omfatter, hovedsakelig består eller består av aminosyrene 417 til 493 til SEQ ID NR:2, 417 til 494 til SEQ ID NR:2, 417 til 495 til SEQ ID NR:2, 417 til 496 til SEQ ID NR:2, 417 til 497 til SEQ ID NR:2, 417 til 498 til SEQ ID NR:2, 417 til 499 til SEQ ID NR:2, 417 til 500 til SEQ ID NR:2, 417 til 492 til SEQ ID NR:2, 417 til 491 til SEQ ID NR:2, 412 til 493 til SEQ ID NR:2, 413 til 493 til SEQ ID NR:2, 414 til

51 til SEQ ID NR:2, 415 til 493 til SEQ ID NR:2, 416 til 493 til SEQ ID NR:2, 411 til 493 til SEQ ID NR:2, 410 til 493 til SEQ ID NR:2, 410 til 494 til SEQ ID NR:2, 411 til 494 til SEQ ID NR:2, 412 til 494 til SEQ ID NR:2, 413 til 494 til SEQ ID NR:2, 414 til 494 til SEQ ID NR:2, 415 til 494 til SEQ ID NR:2, 416 til 494 til SEQ ID NR:2, 417 til 494 til SEQ ID NR:2 og 418 til 494 til SEQ ID NR:2. [0122] I et annet tilfelle inkluderer Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til den gjeldende bindingen av offentliggjøringen et Sp35-polypeptid som omfatter, hovedsakelig består av eller består av peptider til Ig-domenet til Sp35 eller fragmenter, varianter eller avledninger av slike polypeptider. Mer spesifikt er polypeptidene som omfatter, hovedsakelig består av eller består av følgende polypeptidsekvenser: ITX 1 X 2 X 3 (SEQ ID NR:287), ACX 1 X 2 X 3 (SEQ ID NR:288), VCX 1 X 2 X 3 (SEQ ID NR:289) and SPX 1 X 2 X 3 (SEQ ID NR:290) der X 1 is lysin, arginin, histidin, glutamin eller asparagin, X 2 er lysin, arginin, histidin, glutamin eller asparagin, og X 3 er lysin, arginin, histidin, glutamin eller asparagin. Sp35- peptidfragmentene som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem som finnes i bindingen av offentliggjøringen inkluderer f.eks. fragmenter som omfatter, hovedsakelig består av eller består av følgende polypeptidsekvenser: SPRKH (SEQ ID NR:291), SPRICK (SEQ ID NR:292), (SEQ ID NR:320). Disse Sp35-polypeptidene inkluderer den grunnleggende «RKH-sløyfen» (argininlysin histidin-aminosyrer ) i Ig-domenet til Sp35. Andre Sp35-peptider som inkluderer et grunnleggende tripeptid er ITPKRR (SEQ ID NR:321), ACHHK (SEQ ID NR:322) og VCHHK (SEQ ID NR:323). [0123] Andre Sp35-peptidfragmenter som inkluderer visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til den gjeldende bindingen av offentliggjøringen, et Sp35-polypeptid som omfatter, hovedsakelig består av eller består av peptider til Ig-domenet til Sp35 eller fragmenter, varianter eller avledninger av slike polypeptider. Mer spesifikt er peptidene som omfatter, hovedsakelig består av eller består av følgende polypeptidsekvenser: X 4 X 5 RKH (SEQ ID NR:324), X 4 X 5 RRR (SEQ ID NR:325), X 4 X 5 KKK. (SEQ ID NR:326), X 4 X 5 HHH (SEQ ID NR:327), X 4 X 5 RKK (SEQ ID NR:328), X 4 X 5 RKR (SEQ ID NR:329), X 4 X 5 KKH (SEQ ID NR:330), X 4 X 5 HKH (SEQ ID NR:331), X 4 X 5 RRH(SEQ ID NR:332) og X 4 X 5 RHH(SEQ ID NR:333) der X 4 er hvilken som helst aminosyre og X 5 er en hvilken som helst aminosyre.
52 [0124] I et annet tilfelle inkluderer Sp35-peptidfragmenter som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem til den gjeldende bindingen av offentliggjøringen, et Sp35-polypeptid som omfatter, hovedsakelig består av eller består av peptider til Ig-domenet til Sp35 eller fragmenter, varianter eller avledninger av slike polypeptider. Mer spesifikt er polypeptidene som omfatter, hovedsakelig består av eller består av følgende polypeptidsekvenser: ITX 6 X 7 X 8 (SEQ ID NR:334), ACX 6 X 7 X 8 (SEQ ID NR:335), VCX 6 X 7 X 8 (SEQ ID NR:336) og SPX 6 X 7 X 8 (SEQ ID NR:337) der X 6 er lysin, arginin, histidin, glutamin eller asparagin, X 7 er en hvilken som helst aminosyre og X 8 er lysin, arginin, histidin, glutamin eller asparagin. F.eks. et polypeptid som omfatter, hovedsakelig består av eller består av følgende polypeptidsekvenser: SPRLH (SEQ ID NR:338). [0125] Sp35-peptidfragmentene som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem som finnes i den gjeldende bindingen av offentliggjøringen, inkluderer et Sp35-polypeptid som omfatter, hovedsakelig består av eller består av aminosyrene i Igdomenet til Sp35 eller avledninger av den der aminosyren 452 er en tryptofan- eller fenylalaninrest. [0126] Andre Sp35-peptidfragmentene som visse antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem som finnes i den gjeldende bindingen av offentliggjøringen, inkluderer et Sp35-polypeptid som omfatter, hovedsakelig består av eller består av peptidet til det grunnleggende domenet til Sp35. Mer spesifikt peptider som omfatter, hovedsakelig består av eller består av følgende polypeptidsekvenser: RRARIRDRK (SEQ ID NR:339), KKVKVKEKR (SEQ ID NR:340), RRLRLRDRK (SEQ ID NR:341), RRGRGRDRK (SEQ ID NR:342) og RRIRARDRK (SEQ ID NR:343). [0127] Andre eksempler på oppløselige Sp35-polypeptider og metoder og materialer for å skaffe disse molekylene for å produsere antistoffer eller antistoffragmenter av den gjeldende offentliggjøringen finnes f.eks. i PCT-publikasjonsnr. WO 2004/ [0128] Metoder for å lage antistoffer er godt kjent innen faget og beskrives i dette dokumentet. Etter at det er produsert antistoffer til forskjellige fragmenter av eller til Sp35 med full lengde uten signalsekvensen, kan det bestemmes hvilke aminosyrer eller epitoper av Sp35 som antistoffet eller antigenbindende fragmenter bindes til, med epitop-kartleggingsprotokoller slik som beskrevet i dette dokumentet samt metoder som er kjent innen farget (f.eks. dobbel antistoff-sandwich-elisa som beskrevet i kapittel 11 «Immunology, Current Protocols in Molecular Biology, Ed. Ausubel et al., v.2, John Wiley & Sons, Inc. (1996)). Det finnes andre epitop-kartleggingsprotokoller i Morris, G. Epitope Mapping Protocols, New Jersey, USA: Humana Press (1996). Epitopkartlegging kan også utføres på kommersielt tilgjengelige måter (dvs. ProtoPROBE, Inc. (Milwaukee, Wisconsin, USA)). [0129] I tillegg kan antistoffer som produseres og som binder seg til en hvilken som helst del av Sp35, deretter screenes med tanke på deres evne til å fungere som antagonist til Sp35 og dermed fremme nevritt utvekst, nevronal and oligodendrocytt-overlevelse, spredning og differensiering samt fremme myelinering. Antistoffer kan sorteres for oligodendrocytt-/nevronal overlevelse ved å bruke metoden som beskrives i eksempel 10 og 11. I tillegg kan antistoffene screenes med tanke på deres evne til å fremme myelinering ved å bruke metoden i eksempel 9. Til slutt kan antistoffene sorteres med tanke på deres evne til å fremme oligodendrocyttspredning og differensiering samt som nevritt-utvekst ved å bruke metoden som beskrives i eksempel 7. Andre antagonistfunksjoner til antistoffene i den
53 gjeldende offentliggjøringen kan testes ved å bruke andre analyser som beskrives i eksemplene i dette dokumentet. [0130] I andre tilfeller inkluderer den gjeldende offentliggjøringen et antistoff eller antigenbindende fragment, variant eller avledninger av dem som spesifikt eller fortrinnsvis bindes til minst én epitop av Sp35, der epitopen omfatter, hovedsakelig består av eller består av minst omtrent fire til fem aminosyrer av SEQ ID NR:2, minst sju, minst ni eller mellom minst omtrent 15 til omtrent 30 aminosyrer av SEQ ID NR:2. Aminosyrene til en bestemt epitop av SEQ ID NR:2 som beskrives kan være, men trenger ikke å være sammenhengende eller lineære. I visse tilfeller omfatter den minst ene epitopen av Sp35, hovedsakelig består av eller består av en ikkelineær epitop som dannes av det ekstracellulære domenet til Sp35 slik det uttrykkes på en celleflate eller et oppløselig fragment, f.eks. fusjonert til et IgG Fc-område. Dermed, i visse tilfeller omfatter den minst ene epitopen av Sp35, hovedsakelig består av eller består av minst 4, minst 5, minst 6, minst 7, minst 8, minst 9, minst 10, minst 15, minst 20, minst 25, mellom omtrent 15 til omtrent 30, eller minst 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, eller 100 sammenhengende eller ikke-sammenhengende aminosyrer av SEQ ID NR:2 der de ikkesammenhengende aminosyrene danner en epitop ved proteinfolding. [0131] I andre tilfeller inkluderer den gjeldende offentliggjøringen et antistoff eller antigenbindende fragment, variant eller avledning av det som spesifikt eller fortrinnsvis bindes til minst én epitop av Sp35 der epitopen omfatter, hovedsakelig består av eller består av i tillegg til én, to, tre, fire, fem, seks eller flere sammenhengende eller ikke-sammenhengende aminosyrer av SEQ ID NR:2 som beskrevet ovenfor og tilleggsmoietet som modifiserer proteinet. En karbohydratmoietet kan f.eks. inkluderes slik at Sp35-antistoffet bindes med høyere affinitet til modifisert målprotein enn til en umodifisert versjon av proteinet. Som et alternativ binder ikke Sp35-antistoffet i den umodifiserte versjonen av målproteinet i det hele tatt. [0132] I visse tilfeller rettes den gjeldende offentliggjøringen mot et antistoff eller antigenbindende fragment, variant eller avledning av det som spesifikt eller fortrinnsvis til et bestemt Sp35- polypeptidfragment eller fragment av det eller en Sp35-variantpolypeptid med en affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som er mindre enn K D til dette monoklonale antistoff som er referanse. [0133] I visse tilfeller bindes et antistoff eller antigenbindende fragment, variant eller avledning av offentliggjøringen spesifikt til minst én epitop av Sp35 eller fragment eller variant som beskrives ovenfor, dvs. bindes til en slik epitop lettere enn det ville bindes til en urelatert eller tilfeldig epitop, bindes fortrinnsvis til minst én epitop av Sp35 eller fragment eller variant som beskrives ovenfor, dvs. bindes til en slik epitop lettere enn det ville bindes til en relatert, lignende, homolog eller analog epitop. Kompetitivitet hemmer binding av et referanse-antistoff som selv bindes spesifikt eller fortrinnsvis til en viss epitop av Sp35 eller fragment eller variant som beskrives ovenfor, eller bindes til minst én epitop av Sp35 eller fragment eller variant som beskrives ovenfor med en affinitet som karakteriseres av en dissosiasjonskonstant K D som er mindre enn omtrent 5 x 10-2 M, omtrent 10-2 M, omtrent 5 x 10-3 M, omtrent 10-3 M, omtrent 5 x 10-4 M, omtrent 10-4 M, omtrent 5 x 10-5 M, omtrent 10-5 M, omtrent 5 x 10-6 M, omtrent 10-6 M, omtrent 5 x 10-7 M, omtrent 10-7 M, omtrent 5 x 10-8 M, omtrent 10-8 M,
54 53 omtrent 5 x 10-9 M, omtrent 10-9 M, omtrent 5 x M, omtrent M, omtrent 5 x M, omtrent M, omtrent 5 x M, omtrent M, omtrent 5 x M, omtrent M, omtrent 5 x M, omtrent M, omtrent 5 x M eller omtrent M. I et bestemt tilfelle bindes antistoffet eller fragment av det fortrinnsvis til et humant Sp35-polypeptid eller fragment av det i forhold til et murint Sp35-polypeptid eller fragment av det. [0134] Som brukt i konteksten med antistoff-bindende dissosiasjonskonstanter, muliggjør begrepet «omtrent» en variasjonsgrad som finnes ved metodene som brukes for å måle antistoffaffinitet. For eksempel, avhengig av presisjonsnivået til instrumentet som brukes, kan standardfeilen basert på antall prøver som er målt og med avrundingsfeil, ved begrepet «omtrent 10-2 M» f.eks. inkludere fra 0,05 M til 0,005 M. [0135] I bestemte tilfeller binder et antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen, Sp35-polypeptider eller fragmenter eller varianter av dem med en avrate (k(off)) som er mindre eller like 5 X 10-2 sek -1, 10-2 sek -1, 5 X 10-3 sek -1 eller 10-3 sek -1. Som et alternativ binder et antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen Sp35-polypeptider eller fragmenter eller varianter av det med en av-rate (k(off) som er mindre eller lik 5 X 10-4 sek -1, 10-4 sek -1, 5 X 10-5 sek -1 eller 10-5 sek -1 5 X 10-6 sek -1, 10-6 sek -1, 5 X 10-7 sek -1 eller 10-7 sek -1. [0136] I andre tilfeller binder antistoffet eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen Sp35-polypeptider eller fragmenter eller varianter av det med en på-rate (k(on)) som er større eller lik 10 3 M -1 sek -1, 5 X 10 3 M -1 sek -1, 10 4 M -1 sek -1 eller 5 X 10 4 M -1 sek -1. Som et alternativ binder et antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen Sp35-polypeptider eller fragmenter eller varianter av det ved en på-rate (k(on)) som er større eller lik 10 5 M -1 sek -1, 5 X 10 5 M -1 sek -1, 10 6 M -1 sek -1 eller 5 X 10 6 M -1 sek -1 eller 10 7 M -1 sek - 1. [0137] I forskjellige tilfeller er et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det som beskrevet i dette dokumentet, en antagonist av Sp35-aktivitet. I visse tilfeller blokkeres f.eks. bindingen av et antagonist Sp35-antistoff til Sp35 som uttrykkes på nevroner, myelinforbundet nevritt utveksthemming eller nevronal celledød. I andre tilfeller blokkerer binding av Sp35-antistoffet til Sp35 slik som uttrykt på oligodendrocytter, hemming av oligodendrocyttvekst eller differensiering eller blokkerer demyelinering eller dysmyelinering av CNS-nevroner. [0138] Med mindre det spesifikt står noe annet, henviser «fragment av det» i dette dokumentet i forbindelse med et antistoff til et antigenbindende fragment, dvs. en del av antistoffet som spesifikt bindes til antigenet. I ett tilfelle er Sp35-antistoffet, f.eks. et antistoff av offentliggjøringen, et bispesifikt Sp35-antistoff, bindende polypeptid eller antistoff, f.eks. et bispesifikt antistoff, ministoff, domeneslettet antistoff eller fusjonsprotein med bindingsspesifisitet til mer enn én epitop, f.eks. ett eller flere antigener eller mer enn én epitop på samme antigen. I ett tilfelle har et bispesifikt Sp35- antistoff, bindende polypeptid eller antistoff minst ett bindende domene som er spesifikk for minst én epitop på et målpolypeptid som offentliggjøres i dette dokumentet, f.eks. Sp35. I ett annet tilfelle har et bispesifikt Sp35-antistoff, bindende polypeptid eller antistoff minst ett bindende domene som er spesifikk for en epitop på et målpolypeptid og minst ett målbindende domene som er spesifikt for
55 et medikament eller toksin. I enda ett annet tilfelle har et bispesifikt Sp35-antistoff, bindende polypeptid eller antistoff minst ett bindende domene som er spesifikk for en epitop på et målpolypeptid som offentliggjøres i dette dokumentet, og minst ett bindende domene som er spesifikt for en prekursor. Et bispesifikt Sp35-antistoff, bindende polypeptid eller antistoff kan være et tetravalent antistoff med to målbindende domener som er spesifikke for en epitop til et målpolypeptid som offentliggjøres i dette dokumentet og to målbindende domener som er spesifikke for et andre mål. Dermed kan et tetravalent bispesifikt Sp35-antistoff, bindende polypeptid eller antistoff være bivalent for hver spesifisitet. [0139] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen og som kjent av personer med vanlige ferdigheter i faget, kan omfatte et konstant område som medierer én eller flere effektorfunksjoner. Binding av C1-komponenten til komplementet til et antistoff-konstantområde kan f.eks. aktivere komplementsystemet. Aktivering av komplementet er viktig ved opsonisering og lysering av cellepatogener. Aktivering av komplementet stimulerer også den inflammatoriske responsen og kan også være involvert i autoimmun hypersensitivitet. I tillegg bindes antistoffer til reseptorer på forskjellige celler via Fc-området med et Fc-reseptorbindende sted på antistoff-fc-området som bindes til en Fc-reseptor (FcR) på en celle. Det finnes en rekke Fcreseptorer som er spesifikke for forskjellige klasser av antistoffer, inkludert IgG (gamma-reseptorer), IgE (epsilon-reseptorer), IgA (alfa-reseptorer) og IgM (mu-reseptorer). Binding av antistoff til Fcreseptorer på celleflater utløser en rekke viktige og forskjellige biologiske responser, inkludert oppsluking og destruksjon av antistoffbelagte partikler, fjerning av immunkomplekser, lysering av antistoffbelagte målceller av dreperceller (kalles antistoffavhengig cellemediert cytotoksisitet eller ADCC), frigivelse av inflammatoriske mediatorer, placentaoverføring og kontroll av immunoglobulinproduksjonen. [0140] Følgelig inkluderer visse tilfeller av offentliggjøringen et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det der minst en fraksjon av ett eller flere konstantområdedomener er slettet eller på annen måte endret slik at de har de ønskede biokjemiske egenskapene slik at det reduserer effektorfunksjonene, evnen til ikke-kovalent dimerisering, økt evne til å lokalisere på tumorstedet, redusert serum-halveringstid eller økt serum-halveringstid i forhold til et helt, uendret antistoff med omtrent samme immunogenisitet. Visse antistoffer som brukes ved diagnostiske og behandlingsmetoder som beskrives i dette dokumentet, er f.eks. domeneslettede antistoffer som omfatter en polypeptidkjede som ligner på en tung immunoglobulinkjede, men som mangler minst en del av ett eller flere tunge kjededomener. I visse antistoffer slettes f.eks. et helt domene til det konstante området til det modifiserte antistoffet, f.eks. alle eller en del av C H 2-domenet slettes. [0141] I visse Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem som beskrives i dette dokumentet, kan Fc-delen muteres for å redusere effektorfunksjonen ved bruk av metoder som er kjent innen faget. Sletting eller deaktivering (ved brukt av punktmuteringer eller andre måter) av et konstantområde-domene kan f.eks. redusere Fc-reseptorbindingen av det sirkulerende modifiserte antistoffet og dermed øke tumorlokaliseringen. I andre tilfeller kan det være at modifikasjonene i det konstante området stemmer med den moderate komplemente bindingen til offentliggjøringen og dermed reduseres serum-halveringstiden og den ikke-spesifikke forbindelsen til
56 et konjugert cytotoksin. Enda andre modifikasjoner av det konstante området kan brukes til å modifisere disulfidkoplinger eller oligosakkaridmoieteter som muliggjør forbedret lokalisering på grunn av økt antigenspesifisitet eller antistoff-fleksibilitet. Resulterende fysiologisk profil, biotilgjengelighet og andre biokjemiske effekter av modifikasjonene som f.eks. tumorlokalisering, biodistribusjon og serum-halveringstid, kan lett måles og kvantifiseres ved bruk av velkjente immunologiske metoder uten «undue experimentation». [0142] Modifiserte former av Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, kan lages fra en hel prekursor eller moder-antistoffer ved bruk av metoder som er kjent innen faget. Eksempler på metoder omtales i mer detalj i dette dokumentet. [0143] I visse tilfeller er både det variable og konstante området til Sp35-antistoffene eller de antigenbindende fragmentene, varianten eller avledningen av disse fullt humane. Fullt humane antistoffer kan lages med en metode som er kjent innen faget og som beskrives i dette dokumentet. Fullt humane antistoffer mot et bestemt antigen, kan f.eks. prepareres ved å administrere antigenet til et transgenisk dyr som er modifisert til å produsere slike antistoffer som respons på antigenisk provokasjon, men der den endogene lokasjonen er deaktivert. Eksempler på metoder som kan brukes for å lage slike antistoffer, beskrives i de amerikanske patentene: 6,150,584; 6,458,592; 6,420,140. Andre metoder er kjent innen faget. Fullt humane antistoffer kan likeledes produseres med forskjellige visningsmetoder, f.eks. fage-display eller andre virtuelle visningsmetoder slik som beskrives i mer detalj et annet sted i dette dokumentet. [0144] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse av offentliggjøringen kan lages eller produseres ved bruk av metoder som er kjent innen faget. I visse tilfeller er antistoff-molekyler eller fragmenter av disse «produsert rekombinant», dvs. at det produseres ved bruk av rekombinant DNA-teknologi. Eksempler på metoder for å lage antistoffmolekyler eller fragmenter av disse, omtales mer i detalj et annet sted i dette dokumentet. [0145] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse av offentliggjøringen inkluderer også avledninger som er modifisert, f.eks. ved kovalent kopling av en annen type molekyl til antistoffet slik at den kovalente koplingen ikke hindrer antistoffet i å spesifikt bindes til den kognate epitopen. F.eks., men ikke som begrensende, inkluderer antistoffavledninger antistoffer som er modifisert, f.eks. ved glykosylering, acetylering, pegylering, fosforylering, amidering, derivatisering av kjente beskyttende, blokkeringsgrupper, proteolytisk spalting, kopling til en celleligand eller annet protein, osv. En hvilken som helst av en rekke kjemiske modifikasjoner kan utføres med kjente metoder, inkludert, men ikke begrenset til spesifikk spalting, acetylering, formylering, metabolsk syntese av tunicamycin, osv. I tillegg kan avledningen inneholde én eller flere ikke-klassiske aminosyrer. [0146] I visse tilfeller fremkaller ikke Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledningene av offentliggjøringen en skadelig immunrespons i dyr som skal behandles, f.eks. i et menneske. I ett tilfelle modifiseres Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse av offentliggjøringen for å redusere deres immunogenisitet ved bruk av metoder kjent innen faget. Antistoffer kan f.eks. humaniseres,
57 primatiseres, avimmuniseres eller det kan lages kimære antistoffer. Disse typer antistoffer avledes fra et ikke-humant antistoff, vanligvis et murint eller primat antistoff som beholder eller stort sett beholder de antigenbindende egenskapene til moderantistoffet, men som er mindre immunogent i mennesker. Dette kan oppnås med forskjellige metoder inkludert (a) transplantering av hele ikkehumane variable domener på konstante humanområder for å generere kimære antistoffer, (b) poding av minst en del av ett eller flere ikke-humane komplementaritetsbestemmende områder (CDR-er) i et humant rammeverk og konstante områder med eller uten å beholde kritiske rammeverksrester eller (c) poding av hele de ikke-humane variable domene, med å «dekke» dem med en humanlignende del ved å erstatte overflaterestene. Slike metoder beskrives i Morrison et al., Proc. Natl. Acad. Sci. 81: (1984); Morrison et al., Adv. Immunol. 44:65-92 (1988), Verhoeyen et al., Science 239: (1988), Padlan, Molec. Immun. 28: (1991), Padlan, Molec. Immun. 31: (1994) og de amerikanske patentnr. 5,585,089, 5,693,761, 5,693,762 og 6,190,370. [0147] Avimmunisering kan også brukes for å redusere immunogenisiteten til et antistoff. Slik som brukt i dette dokumentet, inkluderer begrepet «avimmunisering» alterasjon av et antistoff for å modifisere T-celleepitopene (se f.eks. WO A1, WO A2). F.eks. V H - og V L -sekvensen fra startantistoffet analyseres og et humant T-celleepitop-«kart» fra hvert V-område viser stedet til epitopene i forhold til de komplementaritetsbestemmende områdene (CDR-ene) og andre nøkkelrester i sekvensen. De enkleste T-celleepitopene fra T-celle-epitopkartet analyseres for å kunne identifisere alternative aminosyreerstatning med liten fare for å endre aktiviteten til sluttantistoffet. En rekke alternative V H - og V L -sekvenser utformes som omfatter kombinasjoner av aminosyreerstatninger, og disse sekvensene innlemmes deretter i en rekke bindende polypeptider, f.eks. Sp35-spesifikke antistoffer eller immunospesifikke fragmenter av disse for å brukes i diagnostiske- og behandlingsmetoder som offentliggjøres i dette dokumentet og som deretter testes for funksjon. Vanligvis genereres og testes mellom 12 og 24 variant-antistoffer. Komplette tunge og lette kjedegener som omfatter modifiserte V- og human C-områder, klones deretter inni uttrykksvektorer og de påfølgende plasmider føres inn i cellelinjene til produksjon av helt antistoff. Deretter ble antistoffene sammenlignet i egnede biokjemiske og biologiske analyser, og den optimale varianten identifiseres. [0148] Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledningen av disse av offentliggjøringen, kan genereres med en hvilken som helst egnet metode som er kjent innen faget. Polyklonale antistoffer til et antigen av interesse kan produseres med forskjellige prosedyrer som er velkjent innen faget. F.eks. et Sp35-antistoff, f.eks. et bindende polypeptid, f.eks. et Sp35- spesifikt antistoff eller immunospesifikt fragment av det kan administreres til forskjellige vertsdyr inkludert, men ikke begrenset til kaniner, mus, rotter, kyllinger, hamstere, geiter, esler, osv. for å fremkalle produksjonen av sera som inneholder polyklonale antistoffer som er spesifikke for antigenet. Forskjellige hjelpemidler kan brukes for å øke den immunologiske responsen avhengig av vertsartene og inkluderer, men er ikke begrenset til Freund s (komplett og ikke-komplett), mineralgeler som f.eks. aluminiumhydroksid, overflateaktive stoffer som lysolecitin, pluroniske polyoler, polyanioner, peptider, ojleemulsjoner, KHL-er (keyhole limpet hemocyanin), dinitrofenol og
58 potensiell nyttige humane hjelpemidler som BCG (Bacille Calmette-Guerin) og Corynebacterium parvum. Slike hjelpemidler er også velkjent innen faget. [0149] Monoklonale antistoffer kan prepareres ved bruk av mange forskjellige metoder som er kjent innen faget inkludert bruken av hybridom, rekombinant og fage-display eller en kombinasjon av disse. Monoklonale antistoffer kan f.eks. produseres ved å bruke hybridom-metoder inkludert dem som er kjent innen faget og som læres, f.eks. i Harlow et al., Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 2nd ed. (1988), Hammerling et al., i: Monoclonal Antibodies and T-Cell Hybridomas Elsevier, N.Y., (1981). Begrepet «monoklonalt antistoff» slik det brukes i dette dokumentet, er ikke begrenset til antistoffer som produseres med hybridom-metoden. Begrepet «monoklonalt antistoff» henviser til et antistoff som er avledet fra en enkel klon inkludert alle eukaryotiske, prokaryotiske eller fagekloner og ikke av metoden som brukes for å produsere dem. Dermed er begrepet «monoklonalt antistoff» ikke begrenset til antistoffer som produseres med hybridom-metoden. Monoklonale antistoffer kan prepareres ved bruk av bedøvede Sp35-mus for å øke områder med epitopgjenkjenning. Monoklonale antistoffer kan prepareres ved bruk av mange forskjellige metoder som er kjent innen faget inkludert bruken av hybridom, rekombinant og fagedisplay eller en kombinasjon av disse. [0150] Ved bruk av protokoller som er kjent innen faget, heves antistoffene i dyr i et eksempel ved bruk av flere subkutane eller intraperitoneale injeksjoner i et aktuelle antigen (f.eks. renset tumor forbundet med antigener som Sp35 eller celler eller celleekstrakter som omfatter slike antigener) og et hjelpemiddel. Denne immuniseringen fremkaller vanligvis en immunrespons som omfatter produksjonen av antigenreaktive antistoffer fra aktiverte splenocytter eller lymfocytter. Selv om de resulterende antistoffene kan høstes fra serumet til dyret for å gi polyklonale preparater, er det ofte ønskelig å isolere de enkelte lymfocyttene fra milten, lymfekjertlene og perifert blod for å gi homogene preparater med monoklonale antistoffer (MAb-er). Det foretrekkes at lymfocytter skaffes fra milten. [0151] I denne velkjente prosessen (Kohler et al., Nature 256:495 (1975)) fusjoneres de relativt kortlevde eller dødelige lymfocyttene fra et pattedyr som er injisert med antigen, med en udødelig tumorcellelinje (f.eks. myelom-cellelinje) og dermed produseres hybridceller eller «hybridomer» som er både udødelige og har evnen til å produsere genetisk kodet antistoff til B-cellen. De resulterende hybridene skilles ut i enkle generiske stammer ved valg, fortynning og gjenvekst der hver enkel stamme som omfatter spesifikke gener for å danne et enkelt antistoff. De produserer antistoffer som er homogene mot et ønsket antigen og i forbindelse med deres rene genetiske prosentandel, kalles de «monoklonale». [0152] Hybridomceller som prepareres på denne måten, såes og dyrkes i et egnet dyrkingsmedium som helst inneholder ett eller flere stoffer som hemmer veksten eller overlevelsen av ikke-fusjonerte, modermyelomceller. Personer med ferdigheter i faget vil sette pris på at reagenser, cellelinjer og medier til dannelsen, valg og vekst av hybridomer er kommersielt tilgjengelige fra en rekke forskjellige kilder, og standardiserte protokoller er godt etablert. Generelt analyseres dyrkingsmediet som hybdridomcellene vokser i, for å produsere monoklonale antistoffer med det ønskede antigenet. Det foretrekkes at bindingsspesifisiteten til de monoklonale antistoffene som produseres av
59 hybridomcellen, bestemmes med in vitro analyser som immunopresipitasjon, radioimmunoanalyse (RIA) eller enzymkoplet immunoabsorbentanalyse (ELISA). Etter at hybridomcellene er identifisert som produserer antistoffer med ønsket spesifisitet, affinitet og/eller aktivitet, kan klonene subklones ved å begrense fortynningsprosedyrene og dyrkes med standard metoder (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, pp (1986)). I tillegg vil det settes pris på at de monoklonale antistoffene som skilles ut av subklonene, kan skilles fra dyrkingsmedium, ascitisk væske eller serum ved bruk av konvensjonelle renseprosedyrer som protein A, hydroksylapatitt-kromatografi, gel-elektroforese, dialyse eller affinitetskromatografi. [0153] Antistoff-fragmenter som gjenkjenner bestemte epitoper, kan genereres med kjente metoder. Fragmentene Fab og F(ab') 2 kan f.eks. produseres med proteolytisk spalting av immunoglobulinmolekylene ved bruk av slike enzymer som papain (for å produsere Fab-fragmenter) eller pepsin (for å produsere F(ab') 2 -fragmenter). F(ab') 2 -fragmentene inneholder det variable området, det konstante lettkjedeområdet og C H 1-domenet til det tunge kjedet. [0154] Personer med ferdigheter i faget vil også sette pris et at DNA-koding av antistoffer eller antistoff-fragmenter (f.eks. antigenbindende steder) kan også avledes fra antistoff-biblioteker som fagedisplay-biblioteker. Helt spesifikt kan slik fage brukes for å vise antigenbindende domener som uttrykkes fra et repertoar eller kombinasjons-antistoffbibliotek (f.eks. humant eller murint). Fage som uttrykker et antigenbindende domene som binder antigenet av interesse, kan velges eller identifiseres med antigen, f.eks. ved bruk av merket antigen eller antigen bundet eller fanget til en fast flate eller kule. Fage som brukes ved disse metodene, er vanligvis trådaktig fage inkludert fd og M13-bindende domener som uttrykkes fra fage med Fab, Fv OE DAB (enkelt Fv område fra lette til tunge kjeder) eller disulfidstabilisert Fv-antistoffdomener som er rekombinant fusjonert enten med fage-gen III eller gen-viii-protein. Det finnes f.eks. eksempler på metoder i EP B1; U.S. patent. 5,969,108, Hoogenboom, H.R. and Chames, Immunol. Tolday 21:371 (2000); Nagy et al. Nat. Med. 8:801 (2002); Huie et al., Proc. Natl. Acad. Sci. USA 98:2682 (2001); Lui et al., J. Mol. Bioi. 315:1063 (2002). Flere publikasjoner (f.eks. Marks et al., BiolTechnology 10: (1992)) beskriver produksjonen av humane antistoffer med høy affinitet med kjedeomflytting samt som kombinasjonsinfeksjon og in vivo rekombinasjon som strategi for å bygge store fage-biblioteker. I et annet tilfelle kan Ribosomaldisplay brukes for å erstatte bakteriofage som display-plattform (se f.eks. Hanes et al., Nat. Biotechnol. 18: 1287 (2000); Wilson et al., Proc. Natl. Acad. Sci. USA 98:3750 (2001) eller Irving et al., J. Immunol. Methods 248:31 (2001». I enda et annet tilfelle kan celleflate-bibliotekene screenes for antistoffer (Boder et al., Proc. Natl. Acad. Sci. USA 97:10701 (2000), Daugherty et al., J. Immunol. Methods 243:211 (2000)). Slike prosedyrer gir alternativer til tradisjonelle hybridommetoder for å isolere, og til påfølgende kloning av monoklonale antistoffer. [0155] I fagedisplay-metoder vises funksjonelle antistoffdomener på overflaten til fagepartiklene som bærer polynukleotidsekvensene som koder dem. DNA-sekvenser som koder V H - og V L -områdene forstørres fra dyre-cdna-biblioteker (f.eks. humane eller murine cdna-biblioteker til lymfoidvev) eller syntetiske cdna-biblioteker. I visse tilfeller er DNA-koding av V H og V L av en scfv-kopling via PCR og klonet i en fagemidvektor (f.eks. p CANTAB 6 eller pcomb 3 HSS). Vektoren er elektroporert i E. coli, og E. coli er infisert med hjelperfage. Fage som brukes i disse metodene, er vanligvis trådaktig fage
60 som inkluderer fd og M13 og V H - eller V L -områdene er vanligvis rekombinant fusjonert enten til fage gen III eller gen VIII. Fage som uttrykker et antigenbindende domene som binder et antigen av interesse (dvs. et Sp35-polypeptid eller et fragment av det), kan velges eller identifiseres med antigen, f.eks. ved bruk av merket antigen eller antigen bundet eller fanget til en fast flate eller kule. [0156] Tilleggseksempler av fagedisplay-metoder som kan brukes for å lage antistoffer inkluderer de som offentliggjøres i Brinkman et al., J. Immunol. Methods 182:41-50 (1995), Ames et al., J. Immunol. Methods 184: (1995), Kettleborough et al., Eur. J. Immunol. 24: (1994), Persic et al., Gene 187:9-18 (1997), Burton et al., Advances in Immunology 57: (1994), PCTpublikasjoner WO 1992/001047, WO 90/02809, WO , WO 92/01047, WO , WO , WO 95/15982, WO 95/20401 og de amerikanske patentnr. 5,698,426, 5,223,409, 5,403,484, 5,580,717, 5,427,908, 5,750,753, 5,821,047, 5,571,698, 5,427,908, 5,516,637, 5,780,225, 5,658,727, 5,733,743 og 5,969,108. [0157] Som beskrevet i andre henvisninger, kan antistoffkodede områder fra fagen isoleres og brukes til å generere hele antistoffer etter fagevalg inkludert humane antistoffer eller andre antigenbindende fragmenter som ønskes og som uttrykkes i en hvilken som helst ønsket vert inkludert pattedyrceller, insektceller, planteceller, gjær og bakterier. Metoder for å rekombinant produsere Fab-, Fab'- og F(ab')2-fragmenter kan f.eks. også anvendes ved bruk av metoder som er kjent innen faget som f.eks. dem som offentliggjøres i PCT-publikasjonen WO 92/22324, Mullinax et al., BioTechniques 12(6): (1992) og Sawai et al., AJRI 34:26-34 (1995) og Better et al., Science 240: (1988). [0158] Eksempler på metoder som kan brukes til å produsere enkeltkjedede Fvs og antistoffer inkluderer dem som beskrives i de amerikanske patentnr. 4,946,778 og 5,258,498, Huston et al., Methods in Enzymology 203:46-88 (1991), Shu et al., PNAS 90: (1993) og Skerra et al., Science 240: (1988). Ved noen bruk, inkludert in vivo bruk av antistoff i mennesker og i in vitro påvisningsanalyser, kan det være at bruken av kimæriske, humaniserte eller humane antistoffer foretrekkes. Et kimærisk antistoff er et molekyl der forskjellige deler av antistoffet avledes fra forskjellige dyrearter, som f.eks. antistoffer med et variabelt område som er avledet fra et murint monoklonalt antistoff og et humant immunoglobulinkonstant område. Metoder for å produsere kimæriske antistoffer er kjent innen faget. Se f.eks. Morrison, Science 229:1202 (1985), Oi et al., BioTechniques 4:214 (1986), Gillies et al., J. Immunol. Methods 125: (1989), amerikanske patentnr. 5,807,715; 4,816,567 og 4, Humaniserte antistoffer er antistoffmolekyler som avledes fra en ikke-human art som binder det ønskede antigenet som har ett eller flere komplementaritetsbestemmende områder (CDR-er) fra den ikke-humane arten og rammeverksområder fra et humant immunoglobulinmolekyl. Ofte erstattes rammeverksrester i humane rammeverksområder med tilsvarende rester fra CDR-donorantistoff for å endre, helst forbedre, antigenbindingen. Disse rammeverkserstatningene identifiseres med metoder som er velkjent innen faget ved å modellere samhandlingene til CDR og rammeverksrestene for å identifisere rester som er viktige ved antigenbinding og påfølgende sammenligning for å identifisere uvanlige rammeverksrester i bestemte posisjoner. (Se f.eks. Queen et al., amerikansk patentnr. 5,585,089, Riechmann et al.,
61 Nature 332:323 (1988). Antistoffer kan humaniseres ved bruk av en rekke forskjellige metoder som er kjent innen faget som f.eks. inkluderer CDR-poding (EP 239,400, PCT-publikasjon WO 91/09967, amerikansk patentnr. 5,225,539, 5,530,101 og 5,585,089), finering eller ny overflate (EP 592,106, EP 519,596, Padlan, Molecular Immunology 28(4/5): (1991), Studnicka et al., Protein Engineering 7(6): (1994), Roguska. et al., PNAS 91: (1994)) og kjedeomflytting (amerikansk patentnr. 5,565,332). [0159] Helt humane antistoffer er spesielt ønskelige ved terapeutisk behandling av humane pasienter. Humane antistoffer kan lages med en rekke forskjellige metoder som er kjent innen faget inkludert fagedisplay-metoder som beskrives ovenfor ved bruk av antistoff-biblioteker som avledes fra humane immunoglobulinsekvenser. Se også de amerikanske patentnr. 4,444,887 og 4,716,111 og PCTpublikasjonene WO 98/46645, WO 98/50433, WO 98/24893, WO 98/16654, WO 96/34096, WO 96/33735 og WO 91/ [0160] Humane antistoffer kan også produseres ved bruk av transgeniske mus som ikke er i stand til å uttrykke funksjonelle endogene immunoglobuliner, men som kan uttrykke humane immunoglobulingener. De humane tung- og lettkjede immunoglobulin-genkompleksene kan føres tilfeldigvis eller ved homolog rekombinasjon inn i stamceller i musefoster. Som et alternativ kan det humanvariable området, det konstante området og mangfoldsområdet føres inn i stamcellene på musefosteret i tillegg til de humane tung- og lettkjedegenene. De tung- og lettkjede immunoglobulingenene fra mus kan gjøres ikke-funksjonelle hver for seg eller samtidig med innføring av de humane immunoglobulinstedene med homolog rekombinasjon. Mer spesifikt hindrer homozygsletting av JH-området endogene antistoffproduksjon. De modifiserte fosterstemcellene utvides og mikroinjiseres i blastocytter for å produsere kimæriske mus. De kimæriske musene avles deretter for å produsere homozyge avkom som uttrykker humane antistoffer. De transgeniske musene immuniseres på vanlig måte med et valgt antigen, f.eks. hele eller en del av et ønsket målpolypeptid. Monoklonale antistoffer som er rettet mot antigenet, kan skaffes fra immuniserte, transgeniske mus ved bruk av konvensjonell hybridomteknologi. De humane immunoglobulin-transgenene som befinner seg i transgeniske mus, flytter på seg under B-celledifferensiering og gjennomgår deretter klassebytte og somatisk mutasjon. Dermed er det mulig med denne metoden å produsere terapeutisk nyttige IgG-, 1gA-, IgM- og IgE-antistoffer. Se Lonberg and Huszar Int. Rev. Immunol. 13:65-93 (1995) for å finne en oversikt over disse teknologiene for å produsere humane antistoffer. Se f.eks. PCT-publikasjonene WO 98/24893, WO 96/34096, WO 96/33735, de amerikanske patentnr. 5,413,923, 5,625,126, 5,633,425, 5,569,825, 5,661,016, 5,545,806, 5,814,318 og 5,939,598 for å finne en detaljert omtale om denne teknologien for å produsere humane antistoffer og humane monoklonale antistoffer og protokoller for å produsere slike antistoffer. I tillegg kan selskaper som Abgenix, Inc. (Freemont, CA, USA) og GenPharm (San Jose, CA, USA) engasjeres for å levere humane antistoffer som er rettet mot et valgt antigen ved bruk av teknologi som ligner på den som beskrives ovenfor. [0161] Helt humane antistoffer som gjenkjenner en valgt epitop, kan genereres ved å bruke en metode som kalles «styrt valg». I denne fremgangsmåten brukes et valgt ikke-humant monoklonalt antistoff, f.eks. et mus-antistoff, for å styre valget til et helt humant antistoff som gjenkjenner
62 samme epitopen. (Jespers et al., Bio/Technology 12: (1988). Se også det amerikanske patentnr. 5,565,332. [0162] I tillegg kan antistoffer som brukes for å rette inn mot polypeptider til offentliggjøringen, brukes til antiidiotype antistoffer som «imiterer» målpolypeptider ved bruk av metoder som er velkjent hos personer med ferdigheter i faget. (Se f.eks. Greenspan & Bona, FASEB J. 7(5): (1989) og Nissinoff, J. Immunol. 147(8): (1991)). Antistoffer som f.eks. bindes til og kompetitivt hemmer polypeptidmultimerisering og/eller binding av et polypeptid av offentliggjøringen til en ligand, kan brukes til å generere antiidiotyper som «imiterer» polypeptidmultimeriseringen og/eller bindingsdomenet og som en konsekvens, bindes til og nøytraliserer polypeptidet og/eller dets ligand. Slike nøytraliserende antiidiotyper eller Fab-fragmenter av slike antiidiotyper kan brukes i terapeutiske planer for å nøytralisere polypeptidliganden. Slike antiidiotypiske antistoffer kan brukes til å binde et ønsket målpolypeptid og/eller til å binde ligander/reseptorer og dermed blokkeres dets biologiske aktivitet. [0163] DNA-koding av ønskede monoklonale antistoffer kan lett isoleres og sekvenseres ved bruk av konvensjonelle prosedyrer (f.eks. ved bruk av oligonukleotide prober som kan bindes spesifikt til gener som koder de tunge og lette kjedene til murine antistoffer). De isolerte og subklonede hybridomcellene tjener som foretrukket kilde til en slik DNA. Etter isolering kan DNA-en plasseres i uttrykksvektorer som deretter transfekteres inni prokaryotiske eller eukaryotiske vertsceller som f.eks. E. coli-celler, COS-apeceller, CHO (Chinese Hamster Ovary)-celler eller myelomceller som ellers ikke produserer immunoglobuliner. Mer spesifikt kan den isolerte DNA-en (som kan være syntetisk som beskrevet ovenfor) brukes til å klone konstante og variable områdesekvenser til de fremstilte antistoffene som beskrevet i Newman et al. amerikansk patentnr. 5, , innlevert 25. januar I hovedsak innebærer dette ekstraksjon av RNA fra de valgte cellene, omdannelse til cdna og forsterkning av PCR ved bruk av Ig-spesifikke primere. Egnede primere til dette formålet beskrives også i det amerikanske patentnr. 5,658,570. Slik som det omtales i mer detalj nedenfor, kan omdannede celler som uttrykker det ønskede antistoffet, dyrkes i relativt store mengder for å gi kliniske og kommersielle forsyninger av immunoglobulinet. [0164] I ett tilfelle omfatter et Sp35-antistoff til den offentliggjorte oppfinnelsen minst én tungkjede eller lettkjede CDR til et antistoff-molekyl. I et annet tilfelle omfatter et Sp35-antistoff til offentliggjøringen minst to CDR-er fra ett eller flere antistoff-molekyler. I et annet tilfelle omfatter et Sp35-antistoff til offentliggjøringen minst tre CDR-er fra ett eller flere antistoff-molekyler. I et annet tilfelle omfatter et Sp35-antistoff til offentliggjøringen minst fire CDR-er fra ett eller flere antistoff-molekyler. I et annet tilfelle omfatter et Sp35-antistoff til offentliggjøringen minst fem CDRer fra ett eller flere antistoff-molekyler. I et annet tilfelle omfatter et Sp35-antistoff til offentliggjøringen minst seks CDR-er fra ett eller flere antistoff-molekyler. Eksempler på antistoffmolekyler som omfatter minst én CDR som kan inkluderes i Sp35-antistoffer hos forsøksobjekter, beskrives i dette dokumentet. [0165] I et spesifikt tilfelle kan aminosyresekvensen til de variable domenene med tung- og/eller lettkjede, inspiseres for å identifisere sekvensens til de komplementaritetsbestemmende områdene (CDR-ene) ved bruk av metoder som er velkjent innen faget, f.eks. ved å sammenligne med andre
63 kjente aminosyresekvenser med tung- eller lettkjede for å bestemme områdene med sekvenshypervariabiliteten. Ved bruk av rutinerekombinante DNA-metoder, kan én eller flere CDR-er settes inn i rammeverksområdene, f.eks. i humane rammeverksområder for å humanisere ikke-humant antistoff. Rammeverksområdene kan være naturlig oppstående eller overensstemmende rammeverksområder og helst humane rammeverksområder (se f.eks. Chothia et al., J. Mol. Biol. 278: (1998) for å finne en liste med humane rammeverksområder). Det foretrekkes at polynukleotidet genereres med en kombinasjon av rammeverksområdene og CDR-ene koder et antistoff som spesifikt bindes til minst én epitop til et ønsket polypeptid, f.eks. Sp35. Det foretrekkes at én eller flere aminosyreerstatninger kan lages inni rammeverksområdene, og det foretrekkes at aminosyreerstatningen forbedrer bindingen av antistoffet til dets antigen. I tillegg kan slike metoder brukes til å lage aminosyreerstatninger eller sletting av én eller flere cysteinrester i det variable området som deltar i intrakjede-disulfidbinding for å generere antistoff-molekyler som mangler én eller flere intrakjedede-disulfidbindinger. Andre alternativer til polynukleotider dekkes av offentliggjøringen og er innenfor ferdighet i faget. [0166] I tillegg kan metoder som er utviklet for å produsere «kimæriske antistoffer» (Morrison et al., Proc. Natl. Acad. Sci. 81: (1984), Neuberger et al., Nature 312: (1984), Takeda et al., Nature 314: (1985)) ved å spalte gener fra et mus-antistoffmolekyl med den aktuelle antigenspesifisiteten sammen med gener fra et humant antistoff-molekyl med aktuell biologisk aktivitet. Slik som brukt i dette dokumentet, er et kimærisk molekyl et molekyl der forskjellige deler avledes fra forskjellige dyrearter som f.eks. dem med et variabelt område som er avledet fra et murint antistoff og et humant immunoglobulin-konstantområde, f.eks. humaniserte antistoffer. [0167] Som et alternativ kan metodene som beskrives ved produksjonen av antistoffer med enkelkjede (amerikansk patentnr. 4,694,778, Bird, Science 242: (1988), Huston et al., Proc. Natl. Acad. Sci. USA 85: (1988) og Ward et al., Nature 334: (1989) tilpasses for å produsere antistoffer med enkelkjede. Antistoffer med enkelkjede dannes ved å kople de tung- og lettkjedede fragmentene til Fv-området via en aminosyre-bro som resulterer i et antistoff med enkelkjede. Metoder for å sette sammen funksjonelle Fv-fragmenter i E. coli kan også brukes (Skerra et al., Science 242: (1988)). [0168] I enda et annet tilfelle ved den gjeldende offentliggjøringen, omfatter genereringen av humane eller hovedsakelige humane antistoffer i transgeniske dyr (f.eks. mus) som ikke er i stand til endogen immunoglobulinproduksjon (se f.eks. amerikanske patentnr. 6,075,181, 5,939,598, 5,591,669 og 5,589,369. Det har f.eks. blitt beskrevet at homozygsletting av koplingsområdet til tungkjedeantistoffet i kimæriske og og kimlinjede mus fører til full hemming av endogen antistoffproduksjon. Overføring av en human immunoglobingenanalyse av slike mutante mus med kimlinje vil føre til produksjonen av humane antistoffer ved antigen provokasjon. Andre foretrukne måter å generere humane antistoffer på ved bruk av SCID-mus, offentliggjøres i det amerikanske patentnr. 5,811,524. Det vil bli satt pris på at det genetiske materialet forbundet med disse humane antistoffene, kan også isoleres og manipuleres som beskrevet i dette dokumentet.
64 [0169] Enda andre svært effektive måter å generere rekombinante antistoffer på offentliggjøres av Newman, Biotechnology 10: (1992). Spesifikt resulterer denne metoden i generering av primatiserte antistoffer som inneholder apekattvariable domener og humankonstante sekvenser. I tillegg beskrives også denne metoden i den felles overdratt amerikansk patentnr. 5,658,570, 5,693,780 og 5,756,096. [0170] I et annet tilfelle kan lymfocyttene velges med mikromanipulering og de isolerte variable genene. Perifere blodmononukleære celler kan f.eks. isoleres fra et immunisert pattedyr og dyrkes i omtrent 7 dager in vitro. Kulturene kan screenes for spesifikke IgG-er som tilfredsstiller utvelgelseskriteriene. Celler fra positive brønner isoleres. De enkelte Ig-produserende B-cellene kan isoleres med FACS eller ved å identifisere dem i en komplementmediert hemolytisk plateanalyse. Ig-produserende B-celler kan mikromanipuleres i et rør, og V H - og V L -genene kan forsterkes ved bruk av f.eks. RT-PCR. V H - og V L -gener kan klones inn i en antistoff-uttrykksvektor og tranfektert inni cellene (f.eks. eukaryotiske eller prokaryotiske celler) til uttrykk. [0171] Som et alternativ kan de antistoffproduserende cellelinjene velges og dyrkes ved bruk av metoder som er velkjent av personer med ferdigheter i faget. Slike metoder beskrives i en rekke laboratoriehåndbøker og primære publikasjoner. I denne forbindelsen er metoder som er egnet til bruk i offentliggjøringen og som er beskrevet nedenfor, beskrevet i Current Protocols in Immunology, Coligan et.al., Eds., Green Publishing Associates and Wiley-Interscience, John Wiley and Sons, New York (1991). [0172] Antistoffer til bruk i diagnostiske og terapeutiske metoder som beskrives i dette dokumentet, kan produseres med en hvilken som helst metode som er kjent innen faget til syntese av antistoffer, helt spesifikt, ved kjemisk syntese eller aller helst, med rekombinante uttrykksmetoder som beskrives i dette dokumentet. [0173] I ett tilfelle omfatter et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen et syntetisk konstant område der ett eller flere domener er delvis eller helt slettet («domeneslettede antistoffer»). I visse tilfeller omfatter kompatible modifiserte antistoffer domeneslettede konstruksjoner eller varianter i dette dokumentet der hele C H 2-domenet er fjernet (ΔC H 2-konstruksjoner). I andre tilfeller kan et kort tilkoplingspeptid erstatte det slettede domenet for å gi det variable området fleksibilitet eller bevegelsesfrihet. Personer med ferdigheter i faget vil sette pris på at slike konstruksjoner foretrekkes spesielt på grunn av de regulerende egenskapene til C H 2-domenet på antistoffets katabolske rate. Domeneslettede konstruksjoner kan avledes ved bruk av en vektor (f.eks. fra Biogen IDEC Incorporated) kodet på et humant konstantdomene (se f.eks. WO 02/060955A2 og WO 02/096948A2. Dette eksempelet på en vektor ble konstruert for å slette C H 2-domenet og for å gi en syntetisk vektor som uttrykker et domeneslettet IgG 1 konstant område. [0174] I visse tilfeller er Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledninger av dem av offentliggjøringene, ministoffer. Ministoffer kan lages ved bruk av metodene som beskrives i faget (se f.eks. det amerikanske patentnr. 5,837,821 eller WO 94/09817A1.
65 [0175] I ett tilfelle omfatter et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen et immunoglobulin med tungkjede med sletting eller erstatning av noen få eller til og med en enkel aminosyre så fremt den tillater forbindelse mellom de monomeriske subenhetene. Mutering av en enkel aminosyre i de valgte områdene til C H 2-domenet, kan f.eks. være nok for å stort sett redusere Fc-binding og dermed øke tumorlokalisering. På lignende måte kan det være ønskelig å helt enkelt slette den delen eller flere konstantområde-domener som styrer effektorfunksjonen (f.eks. komplementbinding) som skal moduleres. Slike delvise slettinger av de konstante områdene kan eventuelt forbedre de valgte egenskapene til antistoffet (serumhalveringstid) mens de andre ønskelige funksjonene som er forbundet med det gjeldende konstantområde-domenet, blir værende intakt. Dessuten, som antydet overfor, kan de konstante områdene til de offentliggjorte antistoffene, syntetiseres med mutasjon eller erstatning av én eller flere aminosyrer som forbedrer profilen til den resulterende konstruksjon. I denne sammenhengen kan det være mulig å bryte aktiviteten til et konservert bindingssted (f.eks. Fc-binding) mens konfigurasjonen og den immunogeniske profilen til det modifiserte antistoffet opprettholdes. I enda andre tilfeller omfatter tilførsel av én eller flere aminosyrer til det konstante området for å forbedre de ønskelige egenskapene som f.eks. effektorfunksjon eller sørge for mer cytotoksin eller karbohydratforbindelse. I slike tilfeller kan det være ønskelig å sette inn eller replisere bestemte sekvenser som er avledet fra valgte konstantområde-domener. [0176] Den gjeldende offentliggjøringen har også antistoffer som omfatter, hovedsakelig består av eller består av varianter (inkludert avledninger) av antistoffmolekyler (f.eks. V H -områder og/eller V L - områder) som beskrives i dette dokumentet, hvilke antistoffer eller fragmenter av dem som immunospesifikt bindes til et Sp35-polypeptid eller fragment eller variant av det. Standard metoder som er kjent hos dem med ferdigheter i farget, kan brukes for å innføre mutasjoner i den nukleotide sekvensen som koder et Sp35-antistoff inkludert, men ikke begrenset til stedsrettet mutagenese og PCR-mediert mutagenese som fører til aminosyreerstatninger. Det foretrekkes at variantene (inkludert avledninger) koder mindre enn 50 aminosyreerstatninger, mindre enn 40 aminosyreerstatninger, mindre enn 30 aminosyreerstatninger, mindre enn 25 aminosyreerstatninger, mindre enn 20 aminosyreerstatninger, mindre enn 15 aminosyreerstatninger, mindre enn 10 aminosyreerstatninger, mindre enn 5 aminosyreerstatninger, mindre enn 4 aminosyreerstatninger, mindre enn 3 aminosyreerstatninger eller mindre enn 2 aminosyreerstatninger i forhold til V H -referanseområdet, V H CDR1, V H CDR2, VH H DR3, V L region, V L CDR1, V L CDR2 eller V L CDR3. En «konservativ aminosyreerstatning» er når aminosyreresten erstattes med en aminosyrerest med en sidekjede med lignende ladning. Familier til aminosyrerester som har sidekjeder med lignende ladninger er definert i faget. Disse familiene inkluderer aminosyrer med grunnleggende sidekjeder (f.eks. lysin, arginin, histidin), syreholdige sidekjeder (f.eks. aspartinsyre, glutaminsyre), uladede polarsidekjeder (f.eks. glysin, asparagin, glutamin, serin, treonin, tyrosin, cystein), ikke-polare sidekjeder (f.eks. alanin, valin, leucin, isoleucin, prolin, fenylalanin, metionin, tryptofan), betagrenede sidekjeder (f.eks. treonin, valin, isoleucin) og aromatiske sidekjeder (f.eks. tyrosin, fenylalanin, tryptofan, histidin). Som et alternativ kan mutasjoner innføres tilfeldig langs hele eller deler av kodesekvensen som ved
66 metningsmutagenese og de resulterende mutanter kan screenes for biologisk aktivitet for å identifisere mutanter som beholder aktivitet (f.eks. evnen til å binde et Sp35-polypeptid). [0177] Det er f.eks. mulig å innføre mutasjoner kun i rammeverksområdet eller kun i CDR-områder til et antistoffmolekyl. Innføring av mutasjoner kan være «silent/neutral missense» mutasjoner, dvs. som har ingen eller liten virkning på antistoffets evne til å binde antigen. Disse typene mutasjoner kan være nyttige for å optimalisere kodonbruk eller forbedre hybridom-antistoffets produksjon. Som et alternativ kan ikke-«neutral missense»-mutasjoner endre et antistoffs evne til å binde antigen. Det er sannsynlig at lokasjonen til de fleste «silent/neutral missense» mutasjonene vil være i rammeverksområdene, mens det er sannsynlig at lokasjonen til de fleste ikke-«neutral missense» mutasjoner er i CDR, selv om dette ikke er et absolutt krav. En person med ferdigheter i faget som kunne utforme og teste mutante molekyler med ønskede egenskaper som f.eks. en endring i antigenbindingsaktiviteten eller endring i bindingsaktiviteten (f.eks. forbedringer i antigenbindingsaktiviteten eller endring av antistoff-spesifisiteten). Etter mutagenese kan det kodede proteinet rutinemessig uttrykkes og den funksjonelle og/eller biologiske aktiviteten til det kodede proteinet (f.eks. evnen til immunospesifikk binding av minst én epitop til et Sp35-polypeptid) kan bestemmes ved å bruke metoder som beskrives i dette dokumentet eller med rutinemessige modifisering av metoder som er kjent innen faget IV. POLYNUKLEOTIDKODEDE Sp35-ANTISTOFFER [0178] Den gjeldende offentliggjøringen har også nukleinsyre-molekyler som koder Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledningen av dem av offentliggjøringen. [0179] I ett tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VH) med tungkjede, der minst én av CDR-ene til det variable området med tungkjede eller minst to av CDR-ene til det variable området med tungkjede er minst 80 %, 85 %, 90 % eller 95 % identisk med referanse-tungkjede CDR1-, CDR2- eller CDR3-aminosyresekvenser fra monoklonale Sp35-antistoffer som offentliggjøres i dette dokumentet. Som et alternativ er CDR1-, CDR2- og CDR3-området i VH minst 80 %, 85 %, 90 % eller 95 % identisk med referanse-tungkjede CDR1- CDR2- og CDR3- aminosyresekvensene fra de monoklonale Sp35-antistoffene som offentliggjøres i dette dokumentet. Dermed, i henhold til denne offentliggjøringen har variabelt område med tungkjede til offentliggjøringen CDR1-, CDR2- eller CDR3-polypeptidsekvenser som er relatert til polypeptidsekvensene som vises i tabell 4:

67 66 TABELL 4: Referanse VH CDR1-, CDR2- og CDR3-aminosyresekvenser* 5
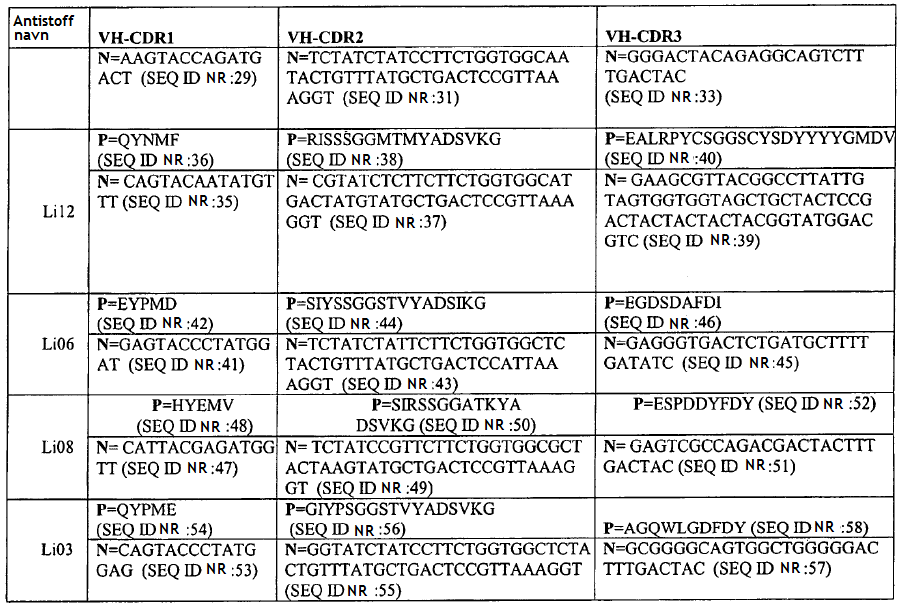
68 67 5
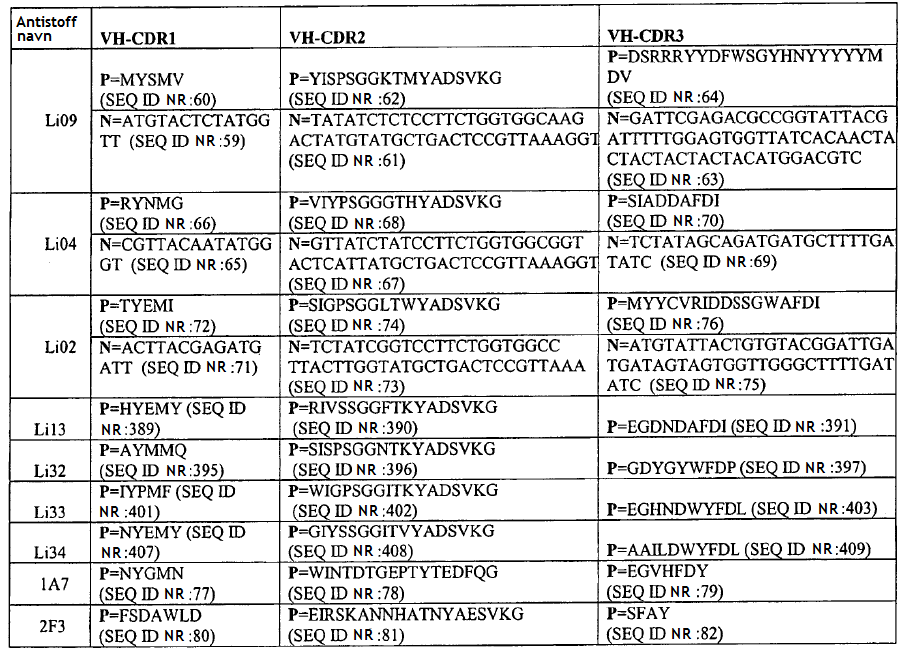
69 68 NO/EP

70 * Bestemmes med Kabat-systemet (se ovenfor). N=nukleotidsekvens P=polypeptidsekvens I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet som bindes spesifikt eller fortrinnsvis til Sp35. [0180] I et annet tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område

71 (VH) med tungkjede der CDR1-, CDR2- og CDR3-området har polypeptidsekvenser som er identiske med CDR1-, CDR2- og CDR3-gruppen som vises i tabell 4. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet, spesifikt eller fortrinnsvis til Sp35. [0181] I et annet tilfelle av offentliggjøringen, gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VH) med tungkjede der CDR1-, CDR2- og CDR3-området er kodet av nukleotidsekvenser som er minst 80 %, 85 %, 90 %, 95 % eller 100 % like VH CDR1-, CDR2- og CDR3- området til immunoglobulin tungkjede-polypeptidet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) eller VH CDR1-, CDR2- og CDR3-området til det immunoglobulin tungkjede-polypeptidet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA- 8107). [0182] I enda et annet tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VH) med tungkjede, der CDR1-, CDR2- og CDR3-området er kodet med polypeptidsekvenser som er identiske med nukleotidsekvensene som kodet CDR1-, CDR2- og CDR3- gruppen som vises i tabell 4. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet, spesifikt eller fortrinnsvis til Sp35. [0183] I et annet tilfelle av offentliggjøringen, gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VH) med tungkjede, der CDR1-, CDR2- og CDR3-området er kodet av nukleotidsekvenser som er minst 80 %, 85 %, 90 %, 95 % eller 100 % lik polynukleotidkodingen av VH CDR1-, CDR2- og CDR3-området til immunoglobulin tungkjede-polypeptidet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) eller VH CDR1-, CDR2- og CDR3-området til immunoglobulin tungkjede-polypeptidet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107). [0184] I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av eller består av et VH som er kodet av ett eller flere av polynukleotidene som beskrives ovenfor spesifikt eller fortrinnsvis til samme epitop som et monoklonalt antistoff som er valgt fra gruppen som består av: 201', 3A3, 3A6, 1A7, 1G7, 2B10, 7P1D5.1G9, 3B5.2 og Li81 og kompetitivt hemmer slikt et monoklonalt antistoff fra å bindes til Sp35. [0185] I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av eller består av et VH som er kodet av ett eller flere av polynukleotidene som beskrives
 som ikke er større enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 x 10-5 M, 10-5 M, 5 X 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X")
72 ovenfor og som spesifikt eller fortrinnsvis bindes til et Sp35-polypeptid eller fragment av det, eller et Sp35-variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 x 10-5 M, 10-5 M, 5 X 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X 10-9 M 10-9 M 5 X M M 5 X M M 5 X M M 5 X M, M, 5 X M, M, 5 X M eller M. [0186] I ett annet tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VL) med lettkjede der minst én av CDR-ene til det variable området med lettkjede eller minst to av CDR-ene til det variable området med lettkjede er minst 80 %, 85 %, 90 % eller 95 % likt referansetungkjede CDR1-, CDR2- eller CDR3-aminosyresekvenser fra monoklonale Sp35-antistoffer som offentliggjøres i dette dokumentet. Som et alternativ er CDR1-, CDR2- og CDR3-området i VL minst 80 %, 85 %, 90 % eller 95 % identisk med referanse-lettkjede CDR1- CDR2- og CDR3-aminosyresekvensene fra de monoklonale Sp35-antistoffene som offentliggjøres i dette dokumentet. Dermed, i henhold til denne offentliggjøringen har det variable området med lettkjede av offentliggjøringen CDR1-, CDR2- eller CDR3-polypeptidsekvensene som er relatert til polypeptidsekvensene som vises i tabell 5: TABELL 5: Referanse VL CDR1-, CDR2- og CDR3-aminosyresekvenser*
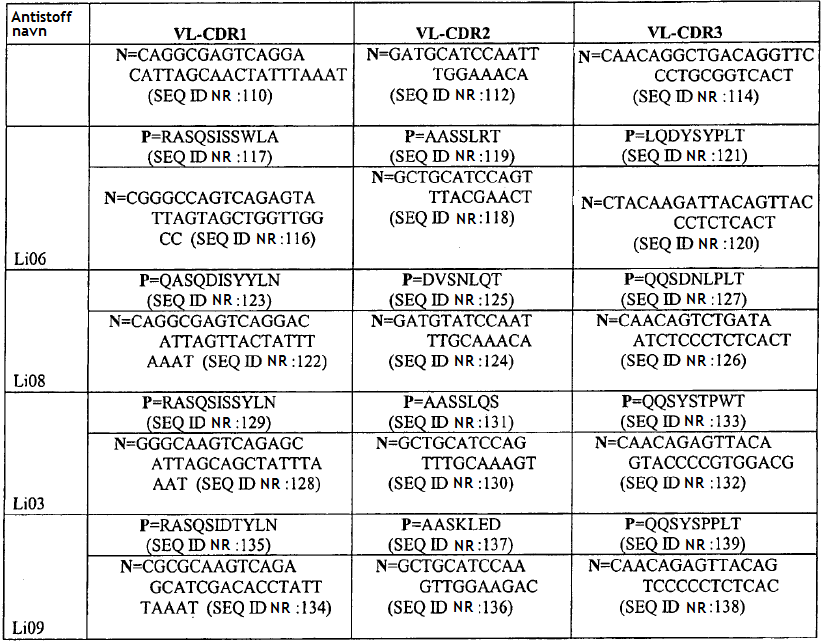
73 72 NO/EP
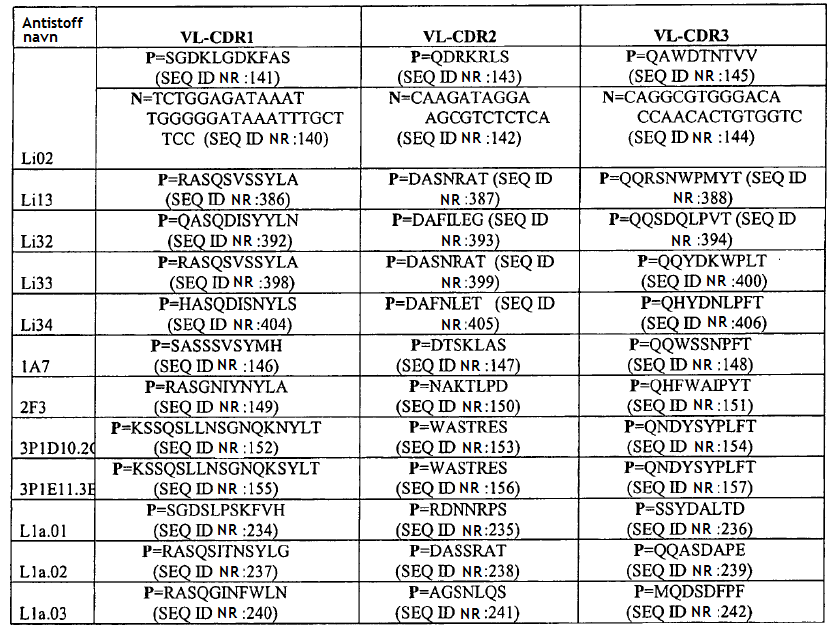
74 73 NO/EP
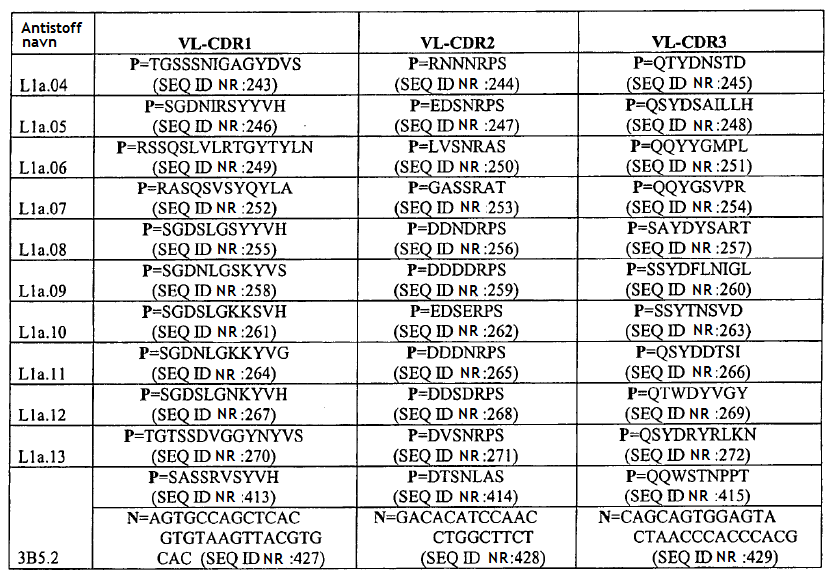
75 74 NO/EP
76 75 5 I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polynukleotidet som bindes spesifikt eller fortrinnsvis til Sp35. [0187] I et annet tilfelle av offentliggjøringen, gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VL) med lettkjede der CDR1-, CDR2- og CDR3-området er kodet av nukleotidsekvenser som er minst 80 %, 85 %, 90 %, 95 % eller 100 % like VL CDR1-, CDR2- og CDR3- området til det immunoglobulin lettkjede-polypeptidet som produseres av hybridom 2.P3B5.2 (ATCC

77 Deposit Designation PTA-8106) eller VL CDR1-, CDR2- og CDR3-området til det immunoglobulin tungkjede-polypeptidet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA- 8107). [0188] I et annet tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VL) med lettkjede der CDR1-, CDR2- og CDR3-området har polypeptidsekvenser som er identiske med CDR1-, CDR2- og CDR3-gruppen som vises i tabell 5. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VL som kodet av polynukleotidet, spesifikt eller fortrinnsvis til Sp35. [0189] I enda et annet tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VL) med lettkjede der CDR1-, CDR2- og CDR3-området er kodet med polypeptidsekvenser som er identiske med nukleotidsekvensene som kodet CDR1-, CDR2- og CDR3- gruppen som vises i tabell 5. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polynukleotidet, spesifikt eller fortrinnsvis til Sp35. [0190] I et annet tilfelle av offentliggjøringen, gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et immunoglobulin variabelt område (VL) med lettkjede der CDR1-, CDR2- og CDR3-området er kodet av nukleotidsekvenser som er minst 80 %, 85 %, 90 %, 95 % eller 100 % lik polynukleotidkodingen av VL CDR1-, CDR2- og CDR3-området til immunoglobulin lettkjede-polypeptidet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) eller VL CDR1-, CDR2- og CDR3-området til immunoglobulin lettkjede-polypeptidet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107). [0191] I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av eller består av et VL som er kodet av ett eller flere av polynukleotidene som beskrives ovenfor spesifikt eller fortrinnsvis til samme epitop som et monoklonalt antistoff som er valgt fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 2B10, 30 7P1D5.1G9, 3B5.2 og Li81 eller vil kompetitivt hemme et slikt monoklonalt antistoff fra å bindes til Sp35. [0192] I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av eller består av et VL som er kodet av ett eller flere av polynukleotidene som beskrives ovenfor, spesifikt eller fortrinnsvis til et Sp35-polypeptid eller fragment av det eller et Sp35-
78 variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 X 10-3 M, 10-3 M, 5 X 10-4 M, 10-4 M, 5 X 10-5 M, 10-5 M, 5 X 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X 10-9 M, 10-9 M, 5 X M, M, 5 X M M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M eller M. [0193] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VH som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse VH polypeptidsekvens valgt fra gruppen som består av SEQ ID NR: 158 til 172, 372, 376, 380, 384 og 416 som vises i tabell 6. I visse tilfeller binder et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet spesifikt eller fortrinnsvis til Sp35. [0194] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyresekvens som koder et VH som har et polypeptidsekvens som er valgt fra gruppen som består av SEQ ID NR: 158 til 172, 372, 376, 380, 384 og 416 som vises i tabell 6. I visse tilfeller binder et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet spesifikt eller fortrinnsvis til Sp35.

79 78 TABELL 6 -VH polypeptidsekvenser VH Sekvens SEQ ID NR

80 79 V H Sekvens SEQ ID NR
![80 VH Sekvens SEQ ID NR 5 10 [0195] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre](/docs-images/25/5927828/images/81-0.png "som koder et VH som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse VH polypeptidsekvens valgt fra gruppen som består av SEQ ID NR: 158-172, 372, 376, 380, 384 og 416.")
81 80 VH Sekvens SEQ ID NR 5 10 [0195] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VH som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse VH polypeptidsekvens valgt fra gruppen som består av SEQ ID NR: , 372, 376, 380, 384 og 416. I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet som bindes spesifikt eller fortrinnsvis til Sp35. [0196] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyresekvens som koder et VH av offentliggjøringen som er valgt fra gruppen som består av SEQ ID NR: ,372,376,380, 384 og 416. I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet som bindes spesifikt eller fortrinnsvis til Sp35.
![81 5 10 [0197] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VH som er](/docs-images/25/5927828/images/82-0.png "minst 80 %, 85 %, 90 % 95 % eller 100 % identisk med en referanse VH polypeptidsekvens valgt fra gruppen som består av immunoglobulin tungkjede-polypeptidet som produseres av hybridom 2.P3B5.")
82 [0197] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VH som er minst 80 %, 85 %, 90 % 95 % eller 100 % identisk med en referanse VH polypeptidsekvens valgt fra gruppen som består av immunoglobulin tungkjede-polypeptidet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) og immunoglobulin tungkjede-polypeptidet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107). [0198] I visse tilfeller bindes et antistoff eller antigenbindende fragment av det som er som omfatter, hovedsakelig består av eller består av et VH som er kodet av ett eller flere polynukleotider som beskrives ovenfor spesifikt eller fortrinnsvis til samme epitop som et monoklonalt antistoff som er valgt fra gruppen som består av (201') 3A3, 3A6, 1A7, 1G7, 2B10, P1D5.1G9 og 3B5.2 eller som vil kompetitivt hemme et slikt monoklonalt antistoff i å bindes til Sp35. [0199] I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av eller består av et VH som er kodet av ett eller flere av polynukleotidene som beskrives ovenfor og som spesifikt eller fortrinnsvis bindes til et Sp35-polypeptid eller fragment av det, eller et Sp35-variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 X 10-5 M, 10-5 M, 5 X 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X 10-9 M, 10-9 M 5 X 10-10, 5 X M, M, M, 5 X M, 5 X M, M, M, 5 X M, M, 5 X M, or M. [0200] I andre tilfeller inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder en tungkjede som er minst 80 %, 85 %, 90 %, 95 % eller 100 % identisk med polynukleotidet til SEQ ID NR: 420 som vist nedenfor. I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter det tunge kjedet kodet av polynukleotidet som bindes spesifikt eller fortrinnsvis til Sp35 eller samme epitop som det monoklonale antistoffet 3B5.2. [0201] Tungkjede-immunoglobulin polynukleotidsekvens til humant murint kimærisk monoklonalt antistoff 3B5.2:
 med tungkjede der polynukleotidet omfatter en VH-nukleinsyresekvens")
83 82 5 CTCTCCCTGTCTCCCGGTTGA (SEQ ID NR:420). [0202] I andre tilfeller inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som koder et variabelt område (V H ) med tungkjede der polynukleotidet omfatter en VH-nukleinsyresekvens som velges fra gruppen som er som omfatter SEQ ID NR: 173 til 184, 370, 374, 378, 382 og 422 som vist i tabell 7. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polynukleotidet, spesifikt eller fortrinnsvis til Sp35.

84 83 TABELL 7 -VH polynukleotidsekvenser VH Sekvens SEQ ID NR

85 84 VH Sekvens SEQ ID NR

86 85 VH Sekvens SEQ ID NR

87 86 VH Sekvens SEQ ID NR

88 87 VH Sekvens SEQ ID NR
![88 VH Sekvens SEQ ID NR 5 10 [0203] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en VH-kodende](/docs-images/25/5927828/images/89-0.png "nukleinsyre som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse-nukleinsyresekvens valgt fra gruppen som består av SEQ ID NR: 173-184, 370, 374, 378, 382 og 422 i tabell 7.")
89 88 VH Sekvens SEQ ID NR 5 10 [0203] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en VH-kodende nukleinsyre som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse-nukleinsyresekvens valgt fra gruppen som består av SEQ ID NR: , 370, 374, 378, 382 og 422 i tabell 7. I visse tilfeller koder polynukleotidet et VHpolypeptid som bindes spesifikt eller fortrinnsvis til Sp35. [0204] I visse tilfeller bindes et antistoff eller antigenbindende fragment av det som er som omfatter, hovedsakelig består av eller består av et VH som er kodet av ett eller flere polynukleotider som beskrives ovenfor spesifikt eller fortrinnsvis til samme epitop som et monoklonalt antistoff som er
90 89 valgt fra gruppen som består av (201 ') 3A3, 3A6, 1A7, 1G7, 2B B5.2 og Li81 eller som vil kompetitivt hemme et slikt monoklonalt antistoff i å bindes til Sp35. [0205] I visse tilfeller bindes et antistoff eller antigenbindende fragment som er som omfatter, hovedsakelig består av eller består av et VH som er kodet av ett eller flere av polynukleotidene som beskrives ovenfor, spesifikt eller fortrinnsvis til et Sp35-polypeptid eller fragment av det eller et Sp35-variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 X 10-5 M, 10-5 M, 5 X 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-9 M, S X 10-9, M, 10-9 M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M eller M. [0206] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VH som er minst 80 %, 85 %, 90 %, 95 % eller 100 % identisk med en referanse VH-polynukleotidsekvens valgt fra gruppen som består av polynukleotidet som koder det tunge immunoglobulin-kjedet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) og polynukleotidet som koder det tunge immunoglobulin-kjedet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA- 8107). [0207] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VL som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse VL polypeptidsekvens valgt fra gruppen som består av SEQ ID NR: 273 til 286, 373, 377, 381, 385 og 417 som vises i tabell 8. I visse tilfeller binder et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polynukleotidet spesifikt eller fortrinnsvis til Sp35. [0208] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyresekvens som koder et VL som har et polypeptidsekvens som er valgt fra gruppen som består av SEQ ID NR: 273 til 286, 373,377, og 417 som vises i tabell 8. I visse tilfeller binder et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polynukleotidet spesifikt eller fortrinnsvis til Sp35.

91 90 TABELL 8 -VL polypeptidsekvenser VL Sekvens SEQ ID NR
![91 VL Sekvens SEQ ID NR 5 10 [0209] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre](/docs-images/25/5927828/images/92-0.png "som koder et VL som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse VL polypeptidsekvens valgt fra gruppen som består av SEQ ID NR: 273 til 286, 373, 377, 381, 385 og 417 som vises i")
92 91 VL Sekvens SEQ ID NR 5 10 [0209] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VL som er minst 80 %, 85 %, 90 % eller 95 % identisk med en referanse VL polypeptidsekvens valgt fra gruppen som består av SEQ ID NR: 273 til 286, 373, 377, 381, 385 og 417 som vises i tabell 8. I visse tilfeller binder et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polynukleotidet spesifikt eller fortrinnsvis til Sp35. [0210] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyresekvens som koder et VL av

93 offentliggjøringen som er valgt fra gruppen som består av SEQ ID NR: 273 til 286, 373, 377, 381, 385 og 417. I visse tilfeller binder et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polynukleotidet spesifikt eller fortrinnsvis til Sp35. [0211] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VL som er minst 80 %, 85 %, 90 %,95 % eller 100 % identisk med en referanse-vl-polypeptidsekvens valgt fra gruppen som består av immunoglobulin lettkjede-polypeptid som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) og immunoglobulin lettkjede-polypeptid som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107). [0212] I visse tilfeller bindes et antistoff eller antigenbindende fragment av det, som er som omfatter, hovedsakelig består av eller består av et VL som er kodet av ett eller flere polynukleotider som beskrives ovenfor spesifikt eller fortrinnsvis til samme epitop som et monoklonalt antistoff som er valgt fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 2B P1D5.1G9, 3B5.2 og Li81 eller som vil kompetitivt hemme et slikt monoklonalt antistoff i å bindes til Sp35. [0213] I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av eller består av et VL som er kodet av ett eller flere av polynukleotidene som beskrives ovenfor og som spesifikt eller fortrinnsvis bindes til et Sp35-polypeptid eller fragment av det, eller et Sp35-variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 x 10-5 M, 10-5 M, 5 X 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X 10-9 M, 10-9 M, 5 x M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M eller M. [0214] I andre tilfeller inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som et lettkjede-immunoglobulin der polynukleotidet er et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et lettkjede som er minst 80 %,85 %,90 %,95 % eller 100 % identisk med polynukleotidet til SEQ ID NR: 421 som vist nedenfor.
![93 [0215] Lettkjede-immunoglobulin polynukleotidsekvens til humant murint kimærisk monoklonalt 5 [0216] I andre tilfeller inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som](/docs-images/25/5927828/images/94-0.png "koder et variabelt område (V L ) med lettkjede der polynukleotidet omfatter en V L -nukleinsyresekvens som velges fra gruppen som er som omfatter SEQ ID-NR 185 til 194, 371, 375, 379, 383 og 423 som")
94 93 [0215] Lettkjede-immunoglobulin polynukleotidsekvens til humant murint kimærisk monoklonalt 5 [0216] I andre tilfeller inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som koder et variabelt område (V L ) med lettkjede der polynukleotidet omfatter en V L -nukleinsyresekvens som velges fra gruppen som er som omfatter SEQ ID-NR 185 til 194, 371, 375, 379, 383 og 423 som vist i tabell 9. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polynukleotidet, spesifikt eller fortrinnsvis til Sp35.

95 94 TABELL 9 -VL polynukleotidsekvenser VL Sekvens SEQ ID NR

96 95 VL Sekvens SEQ ID NR
![96 VL Sekvens SEQ ID NR 5 [0217] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som](/docs-images/25/5927828/images/97-0.png "koder et VL som er minst 80 %, 85 %, 90 % eller 95 % identisk med et VL polynukleotid valgt fra gruppen som består av SEQ ID NR: 185-194,371,375,379,383 og 423 i tabell 9.")
97 96 VL Sekvens SEQ ID NR 5 [0217] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VL som er minst 80 %, 85 %, 90 % eller 95 % identisk med et VL polynukleotid valgt fra gruppen som består av SEQ ID NR: ,371,375,379,383 og 423 i tabell 9. I visse tilfeller koder polynukleotidet et VL polypeptid som bindes spesifikt eller fortrinnsvis til Sp35. [0218] I visse tilfeller bindes et antistoff eller antigenbindende fragment av det som er som omfatter, hovedsakelig består av eller består av et VL som er kodet av ett eller flere polynukleotider som

98 97 beskrives ovenfor spesifikt eller fortrinnsvis til samme epitop som et monoklonalt antistoff som er valgt fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 2B Li81 eller som vil kompetitivt hemme et slikt monoklonalt antistoff i å bindes til Sp35. [0219] I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av eller består av et VL som er kodet av ett eller flere av polynukleotidene som beskrives ovenfor og som spesifikt eller fortrinnsvis bindes til et Sp35-polypeptid eller fragment av det, eller et Sp35-variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 x 10-3 M, 10-3 M, 5 x 10-4 M, 10-4 M, 5 X 10-5 M, 10-5 M, 5 x 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 x 10-8 M, 10-8 M, 5 X 10-9 M, 10-9 M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M eller M. [0220] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av en nukleinsyre som koder et VL som er minst 80 %, 85 %, 90 %, 95 % eller 100 % identisk med en referanse VL polynukleotidsekvens valgt fra gruppen som består av polynukleotidet som koder immunoglobulin-lettkjedet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) og polynukleotidet som koder immunoglobulinlettkjeden som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107). [0221] Et hvilket som helst polynukleotid som beskrives ovenfor, kan i tillegg inkludere flere nukleinsyrer som f.eks. koder et signalpeptid for å styre utskillelsen av det kodede polypeptidet, antistoff-konstantområdene som beskrevet i dette dokumentet eller andre heterologe polypeptider som beskrives i dette dokumentet. [0222] Slik det beskrives i mer detalj et annet sted i dette dokumentet, inkludere den gjeldende offentliggjøringen sammensetninger som omfatter polynukleotidene er som omfatter ett eller flere polynukleotider som beskrives ovenfor. I ett tilfelle inkluderer offentliggjøringen sammensetninger som er som omfatter et første polynukleotid og et andre polynukleotid der det første polynukleotidet koder et VH-polypeptid som beskrevet i dette dokumentet og der det andre polynukleotidet koder en VL-polypeptid som beskrevet i dette dokumentet. Spesifikt, en sammensetning som omfatter, hovedsakelig består av eller består av et VH-polynukleotid som vist i tabell 7 og et VL-polynukleotid som vist i tabell 9 der VH-polynukleotidet og VL-polynukleotidet velges fra gruppen som består av: i) SEQ ID NR:173 og SEQ ID NR:185, ii) SEQ ID NR:174 og SEQ ID NR:186, iii) SEQ ID NR:175 og SEQ ID NR:187, iv) SEQ ID NR:176 og SEQ ID NR:188, v) SEQ ID NR: 178 og SEQ ID NR:189,
99 vi) SEQ ID NR: 179 og SEQ ID NR: 190, vii) SEQ ID NR: 180 og SEQ ID NR: 191, viii) SEQ ID NR:181 og SEQ ID NR:192, ix) SEQ ID NR:182 og SEQ ID NR:193, x) SEQ ID NR:183 og SEQ ID NR:194, xi) SEQ ID NR:370 og SEQ ID NR:371, xii) SEQ ID NR:374 og SEQ ID NR:375, xiii) SEQ ID NR:378 og SEQ ID NR:379, xiv) SEQ ID NR:382 og SEQ ID NR:385, xv) SEQ ID NR:422 og SEQ ID N0:423, xvi) SEQ ID NR: 448 og SEQ ID NR: 449, og xvii) SEQ ID NR:450 og SEQ ID NR:449. [0223] Den gjeldende offentliggjøringen inkluderer også fragmentene av polynukleotidene av offentliggjøringen som beskrevet et annet sted. I tillegg er det også tiltenkt at polynukleotider som koder fusjonspolynukleotidene, Fab-fragmentene og andre avledninger som beskrevet i dette dokumentet, i offentliggjøringen. [0224] Polynukleotidene kan produseres eller fremstilles ved bruk av en hvilken som helst metode som er kjent innen faget. Hvis f.eks. nukleotidsekvensen til antistoffet er kjent, kan som polynukleotidet som koder antistoff, settes sammen med kjemisk syntetiserte oligonukleotider (f.eks. som beskrevet i Kutmeier et al., BioTechniques 17:242 (1994)), som i korthet involverer syntese av overlappende oligonukleotider som inneholder deler av sekvensen som koder antistoffet, knytting (annealing)og ombinding av disse oligonukleotider og deretter forsterkning av de ombundne oligonukleotidene med PCR. [0225] Som et alternativ kan et polynukleotid som koder et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det, genereres fra nukleinsyre fra en egnet kilde. Hvis en klon som inneholder en nukleinsyre som koder et bestemt antistoff, er ikke tilgjengelig, men hvis sekvensen til antistoff-molekylet er kjent, kan koding av antistoffet syntetiseres kjemisk eller skaffes fra en egnet kilde (f.eks. et antistoff-cdna-bibliotek eller generert fra et cdna-bibliotek eller nukleinsyre, helst poly A+RNA, isolert fra et hvilket som helst vev eller celler som uttrykker antistoffet eller annet Sp35-antistoff som f.eks. hybridomceller som velges for å uttrykke et antistoff) med PCR-forsterkning og ved bruk av syntetiske primere som hybridiseres til 3'-enden og 5'-enden av sekvensen eller ved kloning og bruken av oligonukleotider som er probespesifikke for den bestemte gensekvensen for å identifisere f.eks. en cdna-klon fra et cdna-bibliotek som koder antistoffet eller annet Sp35-antistoff. Forsterkede nukleinsyrer som genereres av PCR, kan deretter klones til repliserbare kloningsvektorer ved bruk av en hvilken som helst metode som er velkjent innen faget. [0226] Etter at nukleotidsekvensen og tilsvarende aminosyresekvens til Sp35-antistoffet eller antigenbindende fragment, variant eller avledning av det er bestemt, kan nukleotidsekvensen manipuleres ved bruk av metoder som er velkjent innen faget for å manipulere nukleotidsekvensen, f.eks. rekombinante DNA-metoder, stedsrettet mutagenese, PCR, osv. (se f.eks. metodene som beskrives i Sambrook et al., Molecular Cloning, A Laboratory Manual, 2d Ed., Cold Spring Harbor
100 Laboratory, Cold Spring Harbor, NY, USA (1990) og Ausubel et al., eds., Current Protoeols in Molecular Biology, John Wiley & Sons, NY, USA (1998) for å generere antistoffer med forskjellige aminosyresekvenser, f.eks. for å opprette aminosyreerstatninger, slettinger og/eller innføringer. [0227] Et polynukleotid som koder et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det, kan omfatte et hvilket som helst polyribonukleotid eller polydeoksribonukleotid som kan være umodifisert RNA eller DNA eller modifisert RNA eller DNA. Et polynukleotidkodet Sp35- antistoff eller antigenbindende fragment, variant eller avledning av det, kan f.eks. omfatte enkel- og dobbeltrådet DNA, DNA som er en blanding av enkel- og dobbeltrådede områder, enkel- og dobbeltrådet RNA, og RNA som er en blanding av enkel- og dobbeltrådede områder, hybridmolekyler som omfatter DNA og RNA som kan være enkeltrådede eller mer vanlig, dobbeltrådede eller en blanding av av enkel- og dobbeltrådede områder. I tillegg kan et polynukleotid som koder et Sp35- antistoff eller antigenbindende fragment, variant eller avledning av det, omfatte trippeltrådede områder som omfatter RNA eller DNA eller både RNA og DNA. Et polynukleotid som koder et Sp35- antistoff eller antigenbindende fragment, variant eller avledning av det, kan også inneholde én eller flere modifiserte baser eller DNA- eller RNA-ryggmarg som er modifisert med tanke på stabilitet eller andre grunner. «Modifiserte» baser inkluderer f.eks. tritylerte baser og uvanlige baser som inosin. En rekke forskjellige modifikasjoner kan gjøres til DNA og RNA. Dermed dekker «polynukleotid» kjemiske, enzymatiske eller metabolske modifiserte former. [0228] Et isolert polynukleotid som koder en ikke-naturlig variant av et polypeptid som er avledet fra et immunoglobulin (f.eks. tungkjededel av immunoglobulinet eller lettkjededel), kan lages ved å føre én eller flere nukleotiderstatninger, tillegg eller slettinger inn i nukleotidsekvensen til immunoglobulinet slik at én eller flere aminosyreerstatninger, tillegg eller slettinger kan føres inn i det kodede proteinet. Mutasjoner kan føres inn med standard metoder, som f.eks. stedsrettet mutagenese og PCR-mediert mutagenese. Det foretrekkes at konservative aminosyreerstatninger lages ved én eller flere ikke-essensielle aminosyrerester. V. SP35-ANTISTOFF-POLYPEPTIDER [0229] I tillegg rettes den gjeldende offentliggjøringen mot isolerte polypeptider som utgjør Sp35- antistoffene, de antigenbindende fragmentene, variantene og avledningene av dem. Sp35-antistoffene til den gjeldende offentliggjøringen omfatter polypeptider, f.eks. aminosyresekvenser som koder Sp35 spesifikke antigenbindende områder som er avledet fra immunoglobulinmolekyler. Et polypeptid eller en aminosyresekvens som er «avledet fra» et utpekt protein, henviser til polypeptidets opphav. I visse tilfeller har polypeptidet eller aminosyresekvensen som er avledet fra et bestemt startpolypeptid eller aminosyresekvens en aminosyresekvens som er stort sett identisk med startsekvensen eller en del av den der delen består av minst aminosyrer, minst aminosyrer, minst aminosyrer eller som en personer med vanlige ferdigheter i faget, kan identifisere at den har sitt opphav i startsekvensen. [0230] I ett tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av et immunoglobulin variabelt område (VH) med tungkjede, der
101 minst én av CDR-ene til det variable området med tungkjede eller minst to av CDR-ene til det variable området med tungkjede er minst 80 %, 85 %, 90 % eller 95 % identisk med referanse-tungkjede CDR1-, CDR2- eller CDR3-aminosyresekvenser fra monoklonale Sp35-antistoffer som offentliggjøres i dette dokumentet. Som et alternativ er CDR1-, CDR2- og CDR3-området i VH minst 80 %, 85 %, 90 % eller 95 % identisk med referanse-tungkjede CDR1- CDR2- og CDR3-aminosyresekvensene fra de monoklonale Sp35-antistoffene som offentliggjøres i dette dokumentet. Dermed, i henhold til dette tilfellet, har det variable området med tungkjede i offentliggjøringen, CDR1-, CDR2- eller CDR3- polypeptidsekvenser som er relatert til gruppene i tabell 4 ovenfor. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VH-polypeptidet spesifikt eller fortrinnsvis til Sp35. [0231] I et annet tilfelle gir den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et immunoglobulin variabelt område (VH) med tungkjede, der CDR1-, CDR2- og CDR3-området har polypeptidsekvenser som er identiske med CDR1-, CDR2- og CDR3- gruppen som vises i tabell 4. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VH som er kodet av polypeptidet, spesifikt eller fortrinnsvis til Sp35. [0232] I ett tilfelle gir den gjeldende offentliggjøringen et isolert polynukleotid som omfatter, hovedsakelig består av eller består av et immunoglobulin variabelt område (VH) med tungkjede, der minst én av CDR-ene til det variable området med tungkjede eller minst to av CDR-ene til det variable området med tungkjede er minst 80 %, 85 %, 90%, 95 % eller 100 % identisk med referanse-tungkjede CDR1-, CDR2- og CDR3-aminosyresekvenser valgt fra gruppen som består av VH CDR1-, CDR2, og CDR3- aminosyresekvenser tungkjede-immunoglobulin produsert av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) og VH CDR1-, CDR2 og CDR3-aminosyresekvenser i tungkjede-immunoglobulin produsert av hybridom 7:P1D5.1.G9 (ATCC Deposit Designation PTA-8I07). [0233] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av en VH-polypeptid som er minst 80 %, 85 %, 90%, 95 % eller 100 %identisk med en VH-polypeptid-referansesekvens valgt fra gruppen som består av SEQ ID NR: 158 til 172, 372, 376, 380, 384, 416 og 433 som vist i tabell 6 og SEQ ID NR: 435. I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter VH-polypeptidet som bindes spesifikt eller fortrinnsvis til Sp35. [0234] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et VH-polypeptid valgt fra gruppen som består av SEQ ID NR: 158 til 172, 372, 376, 380, 384, 416 og 433 som vist i tabell 6 og SEQ ID NR:435. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VH-polypeptidet spesifikt eller fortrinnsvis til Sp35. [0235] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et VH-polypeptid som er minst 80 %, 85 %, 90%, 95 % eller 100 % identisk med en referanse VH-polypeptidsekvens valgt fra gruppen som består av immunoglobulin-tungkjedet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA- 8106) og immunoglobulin-tungkjedet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107).
![101 [0236] I visse tilfeller bindes et antistoff eller antigenbindende fragment av det som er som omfatter, hovedsakelig består av eller består av ett eller flere av VH-polypeptidene som beskrives](/docs-images/25/5927828/images/102-0.png "ovenfor, spesifikt eller fortrinnsvis til samme epitopen som et monoklonalt antistoff som velges fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 2B10, 2C11, 2F3, 5 10 15 20 25 30 og Li81 eller")
102 101 [0236] I visse tilfeller bindes et antistoff eller antigenbindende fragment av det som er som omfatter, hovedsakelig består av eller består av ett eller flere av VH-polypeptidene som beskrives ovenfor, spesifikt eller fortrinnsvis til samme epitopen som et monoklonalt antistoff som velges fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 2B10, 2C11, 2F3, og Li81 eller som vil kompetitivt hemme et slikt monoklonalt antistoff i å bindes til Sp35. [0237] I visse tilfeller bindes et antistoff eller antigenbindende fragment som er som omfatter, hovedsakelig består av eller består av ett eller flere av VH-polypeptider som beskrives ovenfor, spesifikt eller fortrinnsvis til et Sp35-polypeptid eller fragment av det eller et Sp35-variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 X 10-3 M, 10,3-3 M, 5 X 10-4 M, 5 X 10-4 M, 5 X 10-5 M, 10-5 M, 5 x 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X 10-9 M, 10-9 M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M eller M. [0238] I ett annet tilfelle gir den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et immunoglobulin variabelt område (VL) med lettkjede der minst én av CDR-ene til det variable området med lettkjede eller minst to av CDR-ene til det variable området med lettkjede er minst 80 %, 85 %, 90 % eller 95 % identisk med referanse-tungkjede CDR1-, CDR2- eller CDR3-aminosyresekvenser fra monoklonale Sp35-antistoffer som offentliggjøres i dette dokumentet. Som et alternativ er CDR1-, CDR2- og CDR3-området i VL minst 80 %, 85 %, 90 % eller 95 % identisk med referanse-lettkjede CDR1- CDR2- og CDR3-aminosyresekvensene fra de monoklonale Sp35-antistoffene som offentliggjøres i dette dokumentet. Dermed, i henhold til denne offentliggjøringen har et variabelt område med lettkjede av offentliggjøringen, CDR1-, CDR2- eller CDR3-polypeptidsekvenser som er relatert til polypeptidsekvensene som vises i tabell 5 ovenfor. I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter VL-polypeptidet som bindes spesifikt eller fortrinnsvis til Sp35. [0239] I et annet tilfelle gir den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et immunoglobulin variabelt område (VL) med lettkjede der CDR1-, CDR2- og CDR3-området har polypeptidsekvenser som er identiske med CDR1-, CDR2- og CDR3- gruppen som vises i tabell 5. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VL som er kodet av polypeptidet, spesifikt eller fortrinnsvis til Sp35. [0240] I ett tilfelle gir den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et immunoglobulin variabelt området (VL) med lettkjede der minst én av CDR-ene til det variable området med tungkjede eller minst to av CDR-ene til det variable området med lettkjede er minst 80 %, 85 %, 90 %, 95 % eller 100 %identisk med referanse-lettkjede
 og VL CDR1-, CDR2 og CDR3-aminosyresekvenser i lettkjede-immunoglobulin produsert av hybridom 7:P1D5.1.G9 (ATCC Deposit Designation PTA-8I07).")
103 CDR1-, CDR2- og CDR3-aminosyresekvenser valgt fra gruppen som består av VL CDR1-, CDR2, og CDR3- aminosyresekvenser lettkjede-immunoglobulin produsert av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) og VL CDR1-, CDR2 og CDR3-aminosyresekvenser i lettkjede-immunoglobulin produsert av hybridom 7:P1D5.1.G9 (ATCC Deposit Designation PTA-8I07). [0241] I enda et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et VL-polypeptid som er minst 80 %, 85 %, 90 % eller 95% identisk med en referanse-vl polypeptidsekvens som velges fra gruppen som består av SEQ ID NR:273 til 286, 373, 377, 381, 385, 417 og 434 som vist i tabell 8. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VL-polypeptidet spesifikt eller fortrinnsvis til Sp35. [0242] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et VL-polypeptid valgt fra gruppen som består av SEQ ID NR: 273 til 286, 373, 377, 381, 385, 417 og 434 som vises i tabell 8. I visse tilfeller bindes et antistoff eller antigenbindende fragment som omfatter VL-polypeptidet, spesifikt eller fortrinnsvis til Sp35. [0243] I et annet tilfelle inkluderer den gjeldende offentliggjøringen et isolert polypeptid som omfatter, hovedsakelig består av eller består av et VL-polypeptid som er minst 80 %, 85 %, 90%, 95 % eller 100 % identisk med en referanse VL-polypeptidsekvens valgt fra gruppen som består av VL til immunoglobulin-lettkjedet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA- 8106) og VL til immunoglobulin-lettkjedet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107). [0244] I visse tilfeller bindes et antistoff eller antigenbindende fragment av det som er som omfatter, hovedsakelig består av eller består av ett eller flere av VL-polypeptidene som beskrives ovenfor, spesifikt eller fortrinnsvis til samme epitopen som et monoklonalt antistoff som velges fra gruppen som består av 201', 3A3, 3A6, 1A7, 1G7, 2B10, 2C11, 2F3, 3P1D10.2C3, og Li81 eller som vil kompetitivt hemme et slikt monoklonalt antistoff i å bindes til Sp35. [0245] I visse tilfeller er et antistoff eller antigenbindende fragment som omfatter, hovedsakelig består av ett eller flere VL-polypeptider som beskrives ovenfor og som spesifikt eller fortrinnsvis bindes til et Sp35-polypeptid eller fragment av det, eller et Sp35-variantpolypeptid med affinitet som karakteriseres av en dissosiasjonskonstant (K D ) som ikke er større enn 5 x 10-2 M, 10-2 M, 5 X 10-3 M, 10-3 M, 5 X 10-4 M, 10-4 M, 5 X 10-5 M, 10-5 M, 5 x 10-6 M, 10-6 M, 5 X 10-7 M, 10-7 M, 5 X 10-8 M, 10-8 M, 5 X 10-9 M, 10-9 M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M, M, 5 X M eller M.
104 [0246] I andre tilfeller omfatter, hovedsakelig består eller består et antistoff eller antigenbindende fragment av et VH-polypeptid som vist i tabell 6 og et VL-polypeptid som vist i tabell 8, valgt fra gruppen som består av: i) SEQ ID NR: 170 og SEQ ID NR:283, ii) SEQ ID NR:171 og SEQ ID NR:284, iii) SEQ ID NR: 172 og SEQ ID NR:285, iv) SEQ ID NR:172 og SEQ ID NR:286, v) SEQ ID NR:158 og SEQ ID NR:273,. vi) SEQ ID NR:159 og SEQ ID NR:274, vii) SEQ ID NR:160 og SEQ ID NR:275, viii) SEQ ID NR:161 og SEQ ID NR:276, ix) SEQ ID NR:163 og SEQ ID NR:277, x) SEQ ID NR:164 og SEQ ID NR:278, xi) SEQ ID NR:165 og SEQ ID NR:279, xii) SEQ ID NR:166 og SEQ ID NR:280, xiii) SEQ ID NR:167 og SEQ ID NR:281, xiv) SEQ ID NR:168 og SEQ ID NR:282, xv) SEQ ID NR:372 og SEQ ID NR:373, xvi) SEQ ID NR:376 og SEQ ID NR:377, xvii) SEQ ID NR:380 og SEQ II) NR:381, xviii) SEQ ID NR:384 og SEQ ID NR:385, xix) SEQ ID NR:416 og SEQ ID NR:417, xx) SEQ ID NR:433 og SEQ ID NR:434, xxi) SEQ ID NR:435 og SEQ ID NR:434. [0247] I andre tilfeller omfatter, hovedsakelig består eller består et antistoff eller antigenbindende fragment av et VH-polypeptid eller et VL-polypeptid som er valgt fra gruppen som består av VHpolypeptidet og VL-polypeptidet som produseres av hybridom 2.P3B5.2 (ATCC Deposit Designation PTA-8106) og VH-polypeptidet og VL-polypeptidet som produseres av hybridom 7.P1D5.1.G9 (ATCC Deposit Designation PTA-8107). [0248] Et hvilket som helst polypeptid som beskrives ovenfor, kan i tillegg inkludere flere polypeptider f.eks. et signalpeptid for å styre utskillelsen av det kodede polypeptidet, antistoffkonstantområdene som beskrevet i dette dokumentet eller andre heterologe polypeptider som beskrives i dette dokumentet. I tillegg inkluderer polypeptidene i offentliggjøringen polypeptidfragmenter som beskrevet i annet sted. Andre polypeptider i offentliggjøringen er fusjonspeptid, Fab-fragmenter og andre avledninger som beskrevet i dette dokumentet. [0249] I tillegg, som beskrevet i mer detalj et annet sted i dette dokumentet, inkluderer offentliggjøringen sammensetninger som omfatter polypeptidene som beskrives ovenfor. [0250] Det vil også bli forstått av en person med vanlige ferdigheter i faget at Sp35-antistoffpolypeptidene som offentliggjøres i dette dokumentet, kan modifiseres slik at aminosyresekvensen varieres i forhold til det naturlige oppstående bindingspolypeptidet som det ble avledet fra. Et
105 polypeptid eller aminosyresekvens avledet fra et utpekt protein kan ligne, f.eks. ha en viss prosentvis identitet med startsekvensen, den kan f.eks. være 60 %, 70 %, 75 %, 80 %, 85 %, 90 % eller 95 % identisk med startsekvensen. [0251] I tillegg kan det lages nukleotid- eller aminosyreerstatninger, -slettinger eller -innsettinger som fører til konservative erstatninger eller endringer i «ikke-essensielle» aminosyreområder. F.eks. et polypeptid eller aminosyresekvens avledet fra et utpekt protein, kan være identisk med startsekvensen unntatt én eller flere individuelle aminosyreerstatninger, -innsettinger eller - slettinger, f.eks. én, to, tre, fire, fem, seks, sju, åtte, ni, ti, femten, tjue eller flere individuelle aminosyreerstatninger, innsettinger eller slettinger. I visse tilfeller har et polypeptid eller aminosyresekvens som er avledet fra et utpekt protein, én til fem, én til ti, én til femten eller én til tjue individuelle aminosyreerstatninger, -innsettinger, eller slettinger i forhold til startsekvensen. [0252] Visse Sp35-antistoff-polypeptider av den gjeldende offentliggjøringen er som omfatter, hovedsakelig består av eller består av et aminosyresekvens som er avledet fra en human aminosyresekvens. Visse Sp35-antistoff-polypeptider består imidlertid av ett eller flere sammenhengende aminosyrer som er avledet fra en annen pattedyrart. Et Sp35-antistoff av den gjeldende offentliggjøringen kan f.eks. inkludere en primat tungkjededel, hengseldel eller antigenbindende område. I et annet eksempel kan ett eller flere murinavledede aminosyrer finnes i et ikke-murint antistoff-polypeptid, f.eks. et antigenbindende stedt ved Sp35-antistoffet. I visse terapeutiske bruksområder utformes Sp35-spesifikke antistoffer eller antigenbindende fragmenter, varianter eller analoger av dem slik at de ikke er immunogeniske i dyr som antistoffet administreres. [0253] I visse tilfeller omfatter et Sp35-antistoffpolypeptid en aminosyresekvens eller én eller flere moieteter som ikke vanligvis forbindes med et antistoff. Eksempler på modifikasjoner beskrives i mer detalj nedenfor. Et enkeltkjedet fv-antistoff-fragment av offentliggjøringen kan omfatte en fleksibel koplingssekvens eller kan modifiseres for å legges til en funksjonell moietet (f.eks. PEG, legemiddel, toksin eller etikett). [0254] Et Sp35-antistoff-polypeptid kan omfatte, hovedsakelig bestå av eller bestå av et fusjonsprotein. Fusjonsproteiner er kimæriske molekyler som f.eks. omfatter et immunoglobulin antigenbindende domene med minst ett målbindingssted og minst én heterologdel, dvs. en del som ikke her naturlig koplet i naturen. Aminosyresekvensene kan vanligvis finnes i separate proteiner som bringes sammen i fusjonspolypeptidet eller at det vanligvis finnes i samme protein, men er plassert i et nytt arrangement i fusjonspolypeptidet. Fusjonsproteiner kan f.eks. lages med kjemisk syntese eller ved å lage og translatere et polynukleotid der peptidområdene kodes i det ønskede forholdet. [0255] Begrepet «heterolog» slik det gjelder et polynukleotid eller et polypeptid, betyr at polynukleotidet eller polypeptidet er avledet fra en atskilt enhet fra resten av enheten som den sammenlignes med. F. eks. Slik som brukt i dette dokumentet avledes «heterologt polypeptid» som kan fusjoneres til et Sp35-antistoffet eller et antigenbindende fragment, en variant eller analog av det fra et ikke-immunoglobulin-polypeptid av samme art eller et immunoglobulin- eller et ikkeimmunoglobulin-polypeptid til en annen art. [0256] En «konservativ aminosyreerstatning» er når aminosyreresten erstattes med en aminosyrerest med lignende sidekjede. Familier til aminosyrerester som har lignende sidekjeder og som er definert i
106 faget, inkluderer grunnleggende sidekjeder (f.eks. lysin, arginin, histidin), syreholdige sidekjeder (f.eks. aspartinsyre, glutaminsyre), uladede polarsidekjeder (f.eks. glysin, asparagin, glutamin, serin, treonin, tyrosin, cystein), ikke-polare sidekjeder (f.eks. alanin, valin, leucin, isoleucin, prolin, fenylalanin, metionin, tryptofan), betagrenede sidekjeder (f.eks. treonin, valin, isoleucin) og aromatiske sidekjeder (f.eks. tyrosin, fenylalanin, tryptofan, histidin). Dermed erstattes en ikkeessensiell aminosyrerest i et immunoglobulin-polypeptid helst med en annen aminosyrerest fra den samme sidekjedefamilien. I et annet tilfelle kan en streng med aminosyrer erstattes med en strukturellignende streng med annen rekkefølge og/eller sammensetning enn sidekjedefamiliemedlemmene. [0257] Som et alternativ kan mutasjoner i et annet tilfelle innføres uregelmessig langs hele eller deler av immunoglobulin-kodingssekvensen, som f.eks. med metningsmutagenese og de resulterende mutantene kan innlemmes i Sp35-antistoffene for å brukes i de diagnostiske og behandlingsmetodene som offentliggjøres i dette dokumentet og screenes for deres evne til å binde til det ønskede antigenet, f.eks. Sp35. VI. FUSJONSPROTEINER OG ANTISTOFF-KONJUGATER [0258] Som omtalt i mer detalj et annet sted i dette dokumentet, kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen i tillegg rekombinant fusjoneres til et heterologt polypeptid ved N- eller C-terminus eller kjemisk konjugeres (inkludert kovalente og ikke-kovalente konjugater) til polypeptidene eller andre sammensetninger. Sp35-spesifikke Sp35-antistoffer kan f.eks. rekombinant fusjoneres eller konjugeres til molekyler som er nyttige som markører i deteksjonsanalyser og effektormolekyler som f.eks. heterologe polypeptider, legemidler, radionuklider eller toksiner. Se f.eks. PCT-publikasjonene WO 92/08495, WO 91/14438, WO 89/12624, amerikansk patentnr. 5,314,995 og EP 396,387. [0259] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, inkluderer avledninger som er modifisert, dvs. ved kovalent festing av en hvilken som helst type molekyl til antistoffet slik at denne kovalente festingen ikke hindrer at antistoffet binder Sp35. F.eks., men ikke som begrensende, inkluderer antistoffavledninger antistoffer som er modifisert, f.eks. ved glykosylering, acetylering, pegylering, fosfylering, fosforylering, amidering, derivatisering av kjente beskyttende/blokkeringsgrupper, proteolytisk spalting, kopling til en celleligand eller annet protein, osv. En hvilken som helst av en rekke kjemiske modifikasjoner kan utføres med kjente metoder, inkludert, men ikke begrenset til spesifikk spalting, acetylering, formylering, metabolsk syntese av tunicamycin, osv. I tillegg kan avledningen inneholde én eller flere ikke-klassiske aminosyrer. [0260] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen kan omfatte aminosyrer som er koplet til hverandre med peptidbindinger eller modifiserte peptidbindinger, dvs. peptididostere og kan inneholde andre aminosyrer enn de 20 genkodede aminosyrene. Sp35-spesifikke antistoffer kan modifiseres med naturlige prosesser som f.eks. posttranslasjonsbehanding eller med kjemiske modifiseringsteknikker som er velkjent innen
107 faget. Slike modifikasjoner er godt beskrevet i grunntekster og i mer detaljerte monografer som i omfangsrikt forskningslitteratur. Modifikasjoner kan skje hvor som helst i det Sp35-spesifikke antistoffet inkludert peptid-ryggraden, aminosyre-sidekjedene og amino- eller karboksylterminus eller på slike moieteter som karbohydrater. Det vil bli satt pris på at samme type modifikasjon kan finnes i samme eller forskjellige grader på flere steder på et bestemt Sp35-spesifikt antistoff. I tillegg kan et bestemt Sp35-spesifikt antistoff inneholde mange typer modifikasjoner. Sp35-spesifikke antistoffer kan ha grener, f.eks. som et resultat av ubiquitinering, og de kan være sykliske, med og uten forgreninger. Sykliske, forgrenede og forgrenede sykliske Sp35-spesifikke antistoffer kan være et resultat av naturlige posttranslasjonsprosesser og kan gjøres med syntetiske metoder. Modifikasjoner inkluderer acetylering, acylering, ADP ribosylering, amidering, kovalent festing av flavin, kovalent festing av en hememoietet, kovalent festing av et nukleotid eller nukleotidavledning, kovalent festing av et lipid eller lipidavledning, kovalent festing av fosfotidylinositol, krysskopling, syklisering, disulfidbindingsformasjon, demetylering, dannelse av kovalente krysskoplinger, dannelse av cystein, dannelse av pyroglutamat, formylering, gamma-karboksylering, glykosylering, GPI-ankerformasjon, hydroksylering, jodering, metylering, myristoylering, oksidering, pegylering, proteolytisk prosessering, fosforylering, prenylering, racemisering, selenoylering, sulfatering, overførings-rna-mediert tilførsel av aminosyrer til proteiner som arginylering og ubiquitinering. (Se f.eks. Proteins - Structure And Molecular Properties, T. E. Creighton, W. H. Freeman and Company, New York 2nd Ed., (1993), Posttranslational Covalent Modification Of Proteins, B. C. Johnson, Ed., Academic Press, New York, pgs (1983); Seifter et al., Meth Enzymol 182: (1990), Rattan et al., Ann NY Acad Sci 663:48-62 (1992)). [0261] Den gjeldende offentliggjøringen sørger også for fusjonsproteiner som er som omfatter et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det og et heterologt polypeptid. Det heterologe polypeptidet som antistoffet er fusjonert med, kan være nyttig for funksjonen eller være nyttig for å rette mot Sp35-polypeptiduttrykkende celler. I ett tilfelle omfatter, hovedsakelig består eller består et fusjonsprotein av et polypeptid som har aminosyresekvensen til ett eller flere av hvilket som helst V H -område til et antistoff av offentliggjøringen eller aminosyresekvensen til ett eller flere av hvilket som helst V L -område til et antistoff av offentliggjøringen eller fragmenter eller varianter av dem og en heterolog polypeptidsekvens. I et annet tilfelle omfatter, hovedsakelig består eller består et fusjonsprotein til bruk i de diagnostiske og behandlingsmetoden som offentliggjøres i dette dokumentet av et polypeptid som har en aminosyresekvens hvilket som helst én, to, tre V H -CDR-er av et Sp35-spesifikt antistoff eller fragmenter, varianter eller avledninger av det eller aminosyresekvenser av hvilket som helst ett, to, tre V L -CDR-er til et Sp35-spesifikt antistoff eller fragmenter, varianter eller avledninger av det og et heterologt polypeptidsekvens. I ett tilfelle omfatter fusjonsproteinet et polypeptid med aminosyresekvensen til et V H -CDR3 av et Sp35-spesifikt antistoff av den gjeldende offentliggjøringen eller fragment, avledning eller variant av det og en heterolog polypeptidsekvens som fusjonsproteinet spesifikt binder minst én epitop av Sp35. I et annet tilfelle omfatter et fusjonsprotein et polypeptid som har aminosyresekvensen til ett V H -område til et Sp35-spesifikt antistoff av offentliggjøringen og aminosyresekvensen til minst ett V L -område til et Sp35-spesifikt antistoff av offentliggjøringen eller
108 fragmenter, avledninger eller varianter av det og en heterolog polypeptidsekvens. Det foretrekkes at V H - og V L -området til fusjonsproteinet korresponderer med et enkelkilde-antistoff (eller scfv- eller Fab-fragment) som spesifikt binder minst én epitop av Sp35. I enda et annet tilfelle av et fusjonsprotein til bruk i diagnostiske og behandlingsmetoder som offentliggjøres i dette dokumentet, omfatter et polypeptid med aminosyresekvens av hvilket som helst én, to, tre eller flere V H -CDR-er av et Sp35-spesifikt antistoff og aminosyresekvens av hvilket som helst én, to, tre eller flere V L -CDR-er av et Sp35-spesifikt antistoff eller fragmenter eller varianter av dem og en heterolog polypeptidsekvens. Det foretrekkes at to, tre, fire, fem, seks eller flere V H -CDR-er eller V L -CDR-er korresponderer med offentliggjøringens enkelkilde-antistoff (eller scfv- eller Fab-fragment). Nukleinsyre-molekyler som koder disse fusjonsproteinene, er også innlemmet i offentliggjøringen. [0262] Eksempler på fusjonsproteiner som rapporteres i litteraturen inkluderer fusjoner av T- cellereseptoren (Gascoigne et al., Proc. Natl. Acad. Sci. USA 84: (1987)), CD4 (Capon et al., Nature 337: (1989), Traunecker et al., Nature 339:68-70 (1989), Zettmeissl et al., DNA Cell Biol. USA 9: (1990) og Bym et al., Nature 344: (1990)), L selectin (homing receptor) (Watson et al., J. Cell. Biol. 110: (1990) og Watson et al., Nature 349: (1991)), CD44 (Aruffo et al., Cell 61: (1990)), CD28 and B7 (Linsleyet et al., J. Exp. Med. 173: (1991)), CTLA-4 (Lisley et al., J. Exp. Med. 174: (1991)), CD22 (Stamenkovic et al., Cell 66: (1991)), 1NF receptor (Ashkenazi et al., Proc. Natl. Acad. Sci. USA 88: (1991), Lesslauer et al., Eur. J. lmmunol. 27: (1991) og Peppel et al., J. Exp. Med. 174: (1991)) og IgE receptor a (Ridgway and Gorman, J. Cell. Biol. Vol. 115, Abstract No (1991)). [0263] I visse tilfeller omfatter Sp35-antistoffene, antistoff-fragmentene, avledningene og variantene av disse en målmoietet i tillegg. Målmoietetene inkluderer et protein eller et peptid som retter lokaliseringen til et viss del av kroppen, f.eks. til hjernen eller kamrene til den. I visse tilfeller er Sp35-antistoffene, antistoff-fragmentene, avledningene og variantene av dem, festet eller fusjonert til en hjernemålmoietet. Hjernemålmoietetene er festet kovalent (f.eks. direkte, overføringsfusjon eller kjemisk kopling direkte eller gjennom et koplingsmolekyl som kan være spaltbar som alternativ) eller ikke-kovalent festet (f.eks. via reverserbare samhandlinger som avidin, biotin, protein A, IgG, osv.). I andre tilfeller er Sp35-antistoffene, antistoff-fragmentene, avledningene og variantene av dem, festet til én hjernemålmoietet til. I enda andre tilfeller er hjernemålmoieteten festet til flere Sp35-antistoffer, antistoff-fragmenter, avledninger og varianter av dem. [0264] En hjernemålmoietet forbundet med et Sp35-antistoff, antistoff-fragment, avledning eller variant av det, forbedrer hjernetilførselen av slike Sp35-antistoffer, antistoff-fragmenter, avledninger og varianter av dem. En rekke polypeptider er beskrevet som når det fusjoneres til et protein eller et terapeutisk middel, leverer proteinet eller det terapeutiske midlet gjennom blodhjernebarrieren (BBB). Ikke-begrensende eksempler inkluderer et enkelt domene-antistoff FC5 (Abulrob et al. (2005) J. Neurachem. 95, ), mab 83-14, et monoklonalt antistoff til den humane insulinreseptoren (Pardridge et al. (1995) Pharmacal. Res. 12, ), B2-, B6- og B8-peptidene som bindes til den humane transferrinreseptoren (htfr) (Xia et al. (2000) J. Virol. 74, ), OX26 monoklonale
109 antistoff til transferrinreseptoren (Pardridge et al. (1991) J. Pharmacol. Exp. Ther. 259, 66-70) og SEQ ID NR: 1-18 i det amerikanske patentnr. 6,306,365. [0265] forbedret hjernetilførelse av Sp35-antistoff, antistoff-fragment, avledning eller variant av det bestemmes på en rekke måter som er godt etablert innen faget. F.eks., administrering til et dyr, radioaktivt, enzymatisk eller fluorescensmerket Sp35-antistoff, antistoff-fragment, avledning og variant av det, som er koplet til en hjernemålmoietet, bestemmelse av hjernelokalisering og sammenligning av lokaliseringen med et ekvivalent radioaktivt, enzymatisk eller fluorescensmerket Sp35-antistoff, antistoff-fragment, avledning eller variant av det som ikke er forbundet med en en hjernemålmoietet. Andre måter å bestemme forbedret innretting som beskrevet i henvisningene ovenfor. [0266] Som omtalt et annet sted i dette dokumentet, kan Sp35-antistoffer eller antigenbindende fragmenter eller varianter av dem av offentliggjøringen, fusjoneres til heterologe polypeptider for å øke in vivo halveringstiden til polypeptidene eller til bruk i immunoanalyser ved bruk av metoder som er kjent innen faget. I ett tilfelle kan f.eks. PEG konjugeres til Sp35-antistoffene til offentliggjøringen for å øke in vivo halveringstiden. Leong, S.R., et al., Cytokine 16:106 (2001), Adv. in Drug Deliv. Rev. 54:531 (2002) eller Weir et al., Biochem. Soc. Transactions 30:512 (2002). [0267] Dessuten kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledning av dem av offentliggjøringen, fusjoneres til markørsekvenser som f.eks. et peptid for å fremme rensing og deteksjon. I det foretrukne tilfellet er markør-aminosyresekvensen et heksahistidin-peptid som f.eks. en tagg som finnes blant annet i pqe-vektoren (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, CA, 91311, USA). Mange av disse er kommersielt tilgjengelige. Slik som f.eks. beskrevet i Gentz et al., Proc. Natl. Acad. Sci. USA 86: (1989), gir heksahistidin konvensjonell rensing av fusjonsproteinet. Andre peptidtagger som er nyttige ved rensing inkluderer, men er ikke begrenset til «HA»-taggen som korresponderer til en epitop som er avledet fra influensa hemagglutininprotein (Wilson et al., Cell 37:767 (1984)) og «flagg»-taggen. [0268] Fusjonsproteinene kan prepareres ved bruk av metoder som er velkjent innen faget (se f.eks. de amerikanske patentnr. 5,116,964 og 5,225,538). Det nøyaktige stedet der fusjonen lages, kan velges empirisk for å optimalisere utskillelses- eller bindingsegenskapene til fusjonsproteinet. DNA som koder fusjonsproteinet transfekteres deretter inn i en vertscelle til uttrykk. [0269] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem ved den gjeldende offentliggjøringen, kan brukes i ikke-konjugert form eller kan konjugeres til minst én av en rekke forskjellige molekyler, f.eks. for å forbedre de terapeutiske egenskapene til molekylet for å fremme måldeteksjon eller til pasientavbildning eller terapi. Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, kan merkes eller konjugeres enten før eller etter rensing når rensing utføres. [0270] Helt spesifikt kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen konjugeres til terapeutiske midler, prekursorer, peptider, proteiner, enzymer, viruser, lipider, biologiske responsmodifikatorer, farmasøytiske midler eller PEG. [0271] Personer med ferdigheter i faget vil sette pris på at konjugater kan også settes sammen ved bruk av en rekke forskjellige metoder avhengig av det valgte midlet som skal konjugeres. F.eks.
110 prepareres konjugater med biotin blant annet ved å reagere et bindingspolypeptid med en aktivert ester av biotin som f.eks. biotin N-hydroksysukkinimidesteren. På lignende måte kan konjugater med en fluorescensmarkør prepareres i tilstedeværelse av et koplingsmiddel, f.eks. de som står oppført i dette dokumentet eller ved en reaksjon med et isotiocyanat, helse fluorescein-isotiocyanat. Konjugater av Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen prepareres på en analog måte. [0272] I tillegg omfatter den gjeldende offentliggjøringen Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledning av dem av offentliggjøringen som konjugeres til et et diagnostisk eller terapeutisk middel. Sp35-antistoffer kan brukes diagnostisk f.eks. for å overvåke utviklingen eller progresjonen av en nevrologisk sykdom som en del av en klinisk testingprosedyre f.eks. for å bestemme effektiviteten til en bestemt behandlingsplan og/eller en forebyggende plan. Deteksjon kan fremmes ved å kople Sp35-antistoffet eller det antigenbindende fragmentet, varianten eller avledningen av det til et detekterbart stoff. Eksempler på detekterbare stoffer inkluderer forskjellige enzymer, prostetiske grupper, fluorescens-materialer, luminescens-materialer, bioluminescens-materialer, radioaktive materialer, positronemitterende metaller ved bruk av forskjellige positronemitterende tomografier og ikke-radioaktive paramagnetiske metallioner. Se f.eks. det amerikanske patentnr. 4,741,900 for metallioner som kan konjugeres til antistoffer til bruk ved diagnostikk iht. den gjeldende offentliggjøringen. Eksempler på egnede enzymer inkluderer pepperrot-peroksidase, alkalisk fosfatase, β-galaktosidase eller acetylkolinesterase, eksempler på egnede prostetiske gruppekomplekser inkludert streptavidinlbiotin og avidin/biotin, eksempler på egnede fluorescens-materialer inkludert umbelliferon, fluorescein, fluorescein-isotiocyanat, rhodamin, diklorotriazinylamin-fluorescein, dansylklorid eller fycoerytrin, et eksempel på et luminescens-materiale inkluderer luminol, eksempler på bioluminescens- materialer inkluderer luciferase, luciferin og aequorin, og eksempler på egnet radioaktiv materiale inkluderer 125 I, 131 I, 111 In eller 99 Tc. [0273] Et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det kan også detekterbart merkes ved å kople det til en kjemiluminescent sammensetning. Tilstedeværelsen av det kjemiluminescent-tagget Sp35-antistoffet bestemmes deretter ved å påvise tilstedeværelsen av luminescens som oppstår i løpet av en kjemisk reaksjon. Eksempler på spesielt nyttige kjemiluminescent-merkingssammensetning er luminol, isoluminol, teromatisk akridiniumester, imidazol, akridiniumsalt og oksalat ester. [0274] Én av måtene som Sp35-antistoffet eller det antigenbindende fragmentet, varianten eller avledningen av det kan detekterbart merkes på er å kople det til et enzym og å bruke det koplede produktet i en enzymimmunoanalyse (EIA) (Voller, A., «The Enzyme Linked Immunosorbent Assay (ELISA)» Microbiological Associates Quarterly Publication, Walkersville, MD, USA, Diagnostic Horizons 2:1-7 (1978), Voller et al., J. Clin. Pathol. 31: (1978), Butler, J. E., Meth. Enrymol. 73: (1981), Maggio, E. (ed.), Enzyme Immunoassay, CRC Press, Boca Raton, Fla., (1980), Ishikawa, E. et al., (eds.), Enzyme Immunoassay, Kgaku Shoin, Tokyo (1981). Enzymet som bindes til Sp35- antistoffet, vil reagere med et egnet substrat, helst et kromogenisk substrat på en slik måte at det
111 produseres en kjemisk moietet som kan påvises, f.eks. med spektrofotometriske, fluorimetriske eller visuelle metoder. Enzymer som kan brukes for å detekterbart merke antistoffet inkluderer, men er ikke begrenset til malatdehydrogenase, stafylokkal nuklease, delta-5-steroidisomerase, gjæralkoholdehydrogenase, alfa-glyserofosfat, dehydrogenase, triosefosfatisomerase, pepperrot-peroksidase, alkalisk fosfatase, asparaginase, glukoseoksidase, beta-galaktosidase, ribonuklease, urease, katalase, glukose-6-fosfatdehydrogenase, glukoamylase og acetylkolinesterase. I tillegg kan påvisning oppnås med kolorimetriske metoder som anvender et kromogenisk substrat til enzymet. Påvisning kan også oppnås med visuell sammenligning av utstrekningen av den enzymatiske reaksjonen til et substrat i forhold til lignende preparerte standarder. [0275] Påvisning kan også oppnås ved å bruke en rekke andre immunoanalyser. Med radioaktiv merking av Sp35-antistoffet eller antigenbindende fragment, variant eller avledning av det, er det f.eks. mulig å påvise antistoffet ved bruk av en radioimmunoanalyse (RIA) (se f.eks. Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, (March, 1986)). Den radioaktive isotopen påvises med metoder som inkluderer, men er ikke begrenset til en gammateller, en scintillasjonsteller eller autoradiografi. [0276] Et Sp35-antistoff eller antigenbindende fragment, variant eller avledning kan detekterbar merkes ved bruk av fluorescensemitterende metaller som 152Eu eller andre i lantanidserien. Disse metallene kan festes til antistoffet ved bruk av slike metallchelaterende grupper som dietylenetriaminpent-eddiksyre (DTPA) eller etylenediaminetetra-eddiksyre (EDTA). [0277] Metoder for å konjugere forskjellige moieteter til et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det er velkjent, se f.eks. Arnon et al., «Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy,» in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp (Alan R. Liss, Inc. (1985); Hellstrom et al., «Antibodies For Drug Delivery,» in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), Marcel Dekker, Inc., pp (1987), Thorpe, «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review,» in Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al. (eds.), pp (1985), «Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy,» i Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), Academic Press pp (1985), and Thorpe et al., «The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates,» Immunol. Rev. 62: (1982). VII. UTTRYKK AV ANTISTOFF-POLYPEPTIDER [0278] Det er velkjent at RNA kan isoleres fra de opprinnelige hybridomcellene eller fra andre transformerte celler med standard metoder som guanidiniumisotiocyanat-ekstrahering og utfelling og påfølgende sentrifugering eller kromatografi. Der det er ønskelig kan mrna isoleres fra total RNA med standard metoder som kromatografi på oligo dt-cellulose. Egnede metoder er kjent innen faget. [0279] I ett tilfelle kan cdna-er som koder de lette og tunge kjedene til antistoffet, gjøres enten samtidig eller separat ved bruk av revers transkriptase og DNA-polymerase iht. velkjente metoder.
112 PCR kan settes i gang med konsensus konstantområde-primere eller mer spesifikke primere basert på den publiserte tung- eller lettkjede-dna og aminosyresekvensen. Som omtalt ovenfor, kan PCR også brukes til å isolere DNA-kloner som koder antistoffets lette og tunge kjeder. I dette tilfellet kan biblioteket screenes med konsensusprimere eller større homologe prober som f.eks. museprober i det konstante området. [0280] DNA, vanligvis plasmid DNA, kan isoleres fra cellene ved bruk av metoder kjent innen faget, begrensninger kan kartlegges og sekvenseres iht. standarden, velkjente metoder fremlegges i detalj, f.eks. i ovennevnte henvisninger i forbindelse med rekombinante DNA-metoder. DNA-en kan selvsagt være syntetiske på et hvilket som helst punkt under isoleringsprosessene eller påfølgende analyse. [0281] Etter manipulering av det isolerte genetiske materialet for å gi Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, settes polynukleotidene som koder Sp35-antistoffene, vanligvis inn en uttrykksvektor til innføring i vertscellen som kan brukes til å produsere den ønskelige mengden Sp35-antistoff. [0282] Rekombinant uttrykk av et antistoff eller fragment, avledning eller analog av det, f.eks. et tung- eller lettkjede av et antistoff som bindes til et målmolekyl som beskrives i dette dokumentet, f.eks. Sp35, krever en konstruksjon av et uttrykksvektor som inneholder et polynukleotid som koder antistoffet. Etter at et polynukleotid koder et antistoff-molekyl eller en tung- eller lettkjede til antistoffet eller en del av det (helst som inneholder det variable domenet med tung- eller lettkjede) av offentliggjøringen, er skaffet, kan vektoren til produksjon av antistoff-molekylet produsere med rekombinant DNA-teknologi ved bruk av metoder som er velkjent innen faget. Dermed beskrives metoder for å preparere et protein ved å uttrykke et polynukleotid som inneholder et antistoff som koder nukleotidsekvensen, i dette dokumentet. Metodene som er velkjent hos personer med ferdigheter i faget, kan brukes til å konstruere uttrykksvektorer som inneholder antistoff-kodede sekvenser og egnede transkripsjonelle og overføringskontrollsignaler. Disse metodene inkluderer f.eks. in vitro rekombinante DNA-metoder, syntetiske metoder og in vivo genetisk rekombinasjon. Dermed gir offentliggjøringen reproduserbare vektorer som er som omfatter en nukleotidsekvens som koder et antistoff-molekyl av offentliggjøringen eller en tung- eller lettkjede av det, eller et variabelt domene med tung- eller lettkjede som er operativt koblet til en promoter. Slike vektorer kan inneholde nukleotidsekvensen som koder det konstante området til antistoff-molekylet (se f.eks. PCTpublikasjon WO 86/05807, PCT-publikasjon WO 89/01036 og det amerikanske patentnr. 5,122,464), og det variable domenet til antistoffet kan klones til en slik vektor for å uttrykke hele den tunge eller lette kjeden. [0283] Vertscellen kan kotransfekteres med to uttrykksvektorer av offentliggjøringen, den første vektoren koder et polypeptid som er avledet fra en tungkjede og en andre vektor som koder et polypeptid som er avledet fra et lettkjede. De to vektorene kan inneholde like valgbare markører som muliggjør likt uttrykk av tung- og lettkjede-polypeptid. Som et alternativ kan en enkel vektor brukes som koder både tung- og lettkjede-polypeptid. I slike situasjoner plasseres det lette kjedet med fordel før det tunge kjedet for å unngå eksess av toksisk fritt tungkjeden(proudfoot, Nature 322:52 (1986), Kohler, Proc. Natl. Acad. Sci. USA 77:2197 (1980)). Kodesekvensen til de tunge og lette kjedene kan omfatte cdna eller genomisk DNA.
113 [0284) Slik begrepet «vektor» eller «uttrykksvektor» brukes i dette dokumentet, betyr det vektorer som brukes iht. den gjeldende offentliggjøringen som et vehikkel for å føre inn eller uttrykke et ønsket gen i en vertscelle. Som kjent hos personer med ferdigheter i faget, kan slike vektorer lett velges fra en gruppe som består av plasmider, fager, viruser og retroviruser. Generelt vil vektorer som er kompatible med offentliggjøringen, omfatte en valgmarkør, egnede begrensningssteder som en hjelp ved kloning av det ønskede genet og evnen til å gå inn og/eller replisere i eukaryotiske eller prokaryotiske celler. [0285] Med tanke på denne offentliggjøringen kan en rekke forskjellige uttrykksvektorsystemer brukes. Én vektorklasse bruker f.eks. DNA-elementer som er avledet fra dyreviruser som bovinpapillomvirus, polyomvirus, adenovirus, vaccinia-virus, baculovirus, retroviruser (RSV, MMTV eller MOMLV) eller SV40-virus. Andre innbefatter bruken av polycistroniske systemer med interne ribosombindende steder. I tillegg kan celler som har integrert DNA-en i deres kromosomer, velges ved å introduseres én eller flere markører som muliggjør valg av transfekterte vertsceller. Markøren kan sørge for prototrofi til en auxotrofisk vert, biosidmotstand (f.eks. antibiotikum) eller motstand mot tunge metaller som kopper. Det valgbare markørgenet kan enten koples direkte til DNA-sekvensene som skal uttrykkes eller føres inn i samme celle med kotransformasjon. Det er mulig at det trenges flere elementer ved optimal syntese av mrna. Disse elementene kan inkludere signalsekvenser, spleisesignaler samt transkripsjonelle promotere, enhancere og avslutningssignaler. [0286] I spesielt foretrukne tilfeller settes klonede variable områdegener i en uttrykksvektor langs med tung- og lettkjede-konstantområdegener (helst humane) som er syntetiske og som omtalt ovenfor. I ett tilfelle gjøres dette ved bruk av en en proprietær uttrykksvektor fra Biogen IDEC, Inc. Som kalles NEOSPLA (amerikansk patentnr. 6,159,730). Denne vektoren inneholder cytomegaloviruspromoter/enhancer, musbetaglobin hovedpromoter, SV40-replikeringsopphav, bovint veksthormon, polyadenyleringesekvens, neomycinfosfotransferase ekson 1 og ekson 2, dihydrofolatreduktasegen og ledesekvens. Man har funnet at denne vektoren fører til svært høyt uttrykksnivå ved antistoffene ved innlemmelse av variable og konstante områdegener, transfeksjon i CHO-celler, med påfølgende valg i G418 som inneholder medium og metotreksatforsterkning. Selvsagt kan en hvilken som helst uttrykksvektor som er i stand til å fremkalle uttrykk i eukaryotiske celler, brukes i den gjeldende offentliggjøringen. Eksempler på egnede vektorer inkluderer, men er ikke begrenset til plasmidene pcdna3, phcmv/zeo, pcr3.1, pef1/his, pind/gs, prc/hcmv2, psv40/zeo2, ptracer-hcmv, pub6/v5-his, pvax1 og pzeosv2 (tilgjengelige hos Invitrogen, San Diego, CA, USA) og plasmidet pci (tilgjengelig hos Promega, Madison, WI, USA). Generelt er screening av et stor antall transformerte celler som uttrykker passende høye nivåer med immunoglobulin tunge og lette kjeder, rutinemessige forsøk som kan f.eks. utføres med robotsystemer. Det instrueres også om vektorsystemer i de amerikanske patentnr. 5,736,137 og 5,658,570. Disse systemene gir høye uttrykksnivåer, f.eks. > 30 pg/celle/dag. And eksempler på vektorsystemer offentliggjøres f.eks. i det amerikanske patentet 6,413,777. [0287] I andre foretrukket tilfeller kan Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledningene av dem av offentliggjøringen, uttrykkes ved å bruke polycistroniske konstruksjoner som f.eks. dem som offentliggjøres i den amerikanske patentsøknaden publikasjonsnr.
114 A1, innlevert 18. november I disse hittil ukjente uttrykkssystemene kan flere genprodukter av interesse som f.eks. tunge og lette kjeder av antistoffer, produseres fra en enkel polycistronisk konstruksjon. Disse systemene bruker et internt ribosominngangssted (IRES) med fordel for å gi relative høye Sp35-antistoffnivåer, f.eks. binde polypeptider, f.eks. Sp35-spesifikke antistoffer eller immunospesifikke fragmenter av dem i eukaryotiske vertsceller. Kompatible IRES-sekvenser offentliggjøres i det amerikanske patentnr. 6,193,980. Personer med ferdigheter i faget vil sette pris på at slike uttrykkssystemer kan brukes til effektiv produksjon av en komplett serie med Sp35- antistoffer som offentliggjøres i den «instant» søknaden. [0288] Mer generelt, etter at vektoren eller DNA-sekvensen som koder en monomerisk subenhet av Sp35-antistoffet, er preparert, kan uttrykksvektoren føres inn i en egnet vertscelle. Innføring av plasmidet i vertscellen kan utføres med forskjellige metoder som er velkjent blant personer med ferdigheter i faget. Disse inkluderer, men ikke begrenset til transfeksjon (inkludert elektroforese og elektroporasjon), protoplastfusjon, kalsiumfosfatutfelling, cellefusjon med innhyllet DNA, mikroinjeksjon og infeksjon med intakt virus. Se Ridgway, A. A. G. «Mammalian Expression Vectors» Vectors, Rodriguez and Denhardt, Eds., Burterworths, Boston, Mass., Chapter 24.2, pp (1988). Vanligvis skjer plasmidinnføring i verten med elektroporasjon. Vertscellene som rommer uttrykkskonstruksjonen, dyrkes under forhold som er egnet til produksjonen av lette kjeder og tunge kjeder og analysert for tung- og/eller lettkjede-proteinsyntese. Eksempler på analysemetoder inkluderer enzymkoplede immunosorbentanalyse (ELISA), radioimmunoanalyse (RIA) eller fluorescensaktivert cellesorteringsanalyser (FAC-er) immunohistokjemi og lignende. [0289] Uttrykksvektoren overføres til en vertscelle med konvensjonelle metoder og de transfekterte cellene dyrkes deretter med konvensjonelle metoder for å produsere et antistoff til bruk i metodene som beskrives i dette dokumentet. Dermed inkluderer offentliggjøringen vertsceller som inneholder et polynukleotid som koder et antistoff til offentliggjøringen eller en tung- eller lettkjede av det som er operativt koplet til en heterolog promoter. I det fortrukne tilfellet ved uttrykk av dobbeltkjedede antistoffer, kan vektorer som koder både de tunge og lette kjede, samuttrykkes i vertscellen til uttrykk av hele immunoglobulinmolekylet som beskrevet nedenfor. [0290] Slik som brukt i dette dokumentet, henviser «vertsceller» til celler som rommer vektorer som er konstruert ved bruk av rekombinante DNA-metoder og som koder minst et heterologt gen. I beskrivelsene av prosessene for å isolere antistoffer fra rekombinante verter, brukes begrepene «celle» og «cellekultur» om hverandre for å benevne antistoffkilden med mindre det tydelig spesifiseres noe annet. Med andre ord kan gjenvinning av polypeptid fra «cellene» betyr enten fra nedspunnede hele celler eller fra cellekulturen som inneholder både mediet og de suspenderte cellene. [0291] En rekke forskjellige vertsuttrykks-vektorsystemer kan brukes til å uttrykke antistoffmolekyler til bruk i metodene som beskrives i dette dokumentet. Slike vertsuttrykkssystemer representer vehikler som kan brukes for å kode sekvenser av interesse og deretter rense dem, men representerer også celler som kan, når de omdannes eller transfekteres med de passende nukleotidkodingssekvensen, uttrykke et antistoffmolekyl til offentliggjøringen in situ. Disse inkluderer, men er ikke begrenset til mikroorganismer som bakterier (f.eks. E. coli, B. subtilis) som er
115 omdannet med rekombinante bakteriofage- DNA-, plasmid-dna eller cosmid DNA-uttrykksvektorer som inneholder antistoff-kodesekvenser, gjær (f.eks. Saccharomyces, Pichia) som er omdannet med rekombinante gjæruttrykksvektorer som inneholder antistoff-kodesekvenser, insektcellesystemer som er infisert med rekombinante virusuttrykksvektorer (e.g., baculovirus) som inneholder antistoffkodesekvenser, plantecellesystemer som er infisert av rekombinante virusuttrykksvektorer (f.eks. blomkål-mosaikkvirus, CaMV, tobakk-mosaikkvirus, TMV eller omdannet med rekombinante plasmiduttrykksvektorer (f.eks. Ti-plasmid) som inneholder antistoff-kodesekvenser eller pattedyrcellesystemer (f.eks. COS-, CHO-, BLK-, 293-, 3T3-celler) som rommer rekombinante uttrykkskonstruksjoner som inneholder promotere som er avledet fra genomet til pattedyrceller (f.eks. metallotionein-promoter) eller fra pattedyrviruser (f.eks. adenovirus senpromoter, vaccinia virus 7,5K-promoter). I beskrivelsene av prosessene for å isolere antistoffer fra rekombinante verter, brukes begrepene «celle» og «cellekultur» om hverandre for å benevne antistoffkilden med mindre det tydelig spesifiseres noe annet. Pattedyrsceller som f.eks. ovarieceller fra kinesisk hamster (CHO), i forbindelse med en vektor som f.eks. hoved intermediat tidlig genpromoterelement fra human cytomegalovirus er f.eks. et effektivt uttrykkssystem for antistoffer (Foecking et al., Gene 45:101 (1986); Cockett et al., Bio/Technology 8:2 (1990)). [0292] Vertscellelinjen som brukes til proteinuttrykk har ofte pattedyropphav. Det anerkjennes at personer med ferdigheter i faget har evnen til å foreta fortrinnsvis bestemmelse av bestemte vertscellelinjer som er best egnet til det ønskede genproduktet som skal uttrykkes i den. Eksempler på vertscellelinjer inkluderer, men er ikke begrenset til CHO (ovarieceller fra kinesisk hamster), DG44 og DUXB11 (ovariecellelinjer fra kinesisk hamster, DHFR-minus), HELA (human cervikal karsinom), CVI (apekattnyre-linje), COS (en avledning av CVI med SV40 T-antigen), VERY, BHK (hamsterunge-nyre), MDCK, 293, WI38, R1610 (kinesisk hamsterfibroblast) BALBC/3T3 (musfibroblast), HAK (hamsternyrelinje), SP2/O (musmyelom), P3x63-Ag3.653 (musmyelom), BFA-1c1BPT (bovine endoteliale celler), RAJI (humant lymfocytt) og 293 (human nyre). CHO-celler er spesielt fortrukket. Vertscellelinjer er vanligvis tilgjengelige fra kommersielle tjenester, American Tissue Culture Collection eller fra publisert litteratur. [0293] I tillegg kan en vertscellestamme velges som modulerer uttrykket til de innsatte sekvensens eller modifiserer eller prosesserer genproduktet på den spesielle måten som ønskes. Slike modifikasjoner(f.eks. glykosylering) og prosessering (f.eks. spalting) av proteinproduktene kan være nyttig når det gjelder proteinets funksjon. Forskjellige vertsceller har karakteristiske og spesifikke mekanismer til posttranslasjonsprosessering og modifikasjon av proteinene og genproduktene. Egnede cellelinjer eller vertssystemer kan velges for å sikre riktig modifikasjon og prosessering av det fremmede proteinet som uttrykkes. Med dette i tankene, kan eukaryotiske vertsceller som inneholder cellemaskinen til riktig prosessering av det primære transkriptet, glykosylering og fosforylering av genproduktet, brukes. [0294] Til langtids, høyytelsesproduksjon av rekombinante proteiner foretrekkes stabilt uttrykk. Cellelinjer som stabilt uttrykker antistoff-molekylet, kan f.eks. konstrueres. Istedenfor å bruke uttrykksvektorer som inneholder virusopphav av replikasjonen, kan vertsceller omdannes med DNA som er kontrollert av egnede uttrykkskontrollelementer (f.eks. promotere, enhancere, sekvenser,
116 transkripsjonsterminatorer, polyadenyleringssteder, osv. og en valgbar markør. Etter innføring av det fremmede DNA, kan konstruerte celler få lov til å vokse i 1 til 2 dager i et beriket medium og deretter byttes til et selektivt medium. Den valgbare markøren i det rekombinante plasmidet overfører motstand til valget og gjør at cellene stabilt integrerer plasmidet inn i deres kromosomer og vokser og danner foci som deretter kan klones og utvides inn i cellelinjene. Denne metoden kan med fordel brukes for å konstruere cellelinjer som kan stabilt uttrykke antistoff-molekylet. [0295) En rekke seleksjonssystemer kan brukes inkludert, men ikke begrenset til herpes simplex virus tymidinkinase (Wigler et al., Cell 11:223 (1977)), hypoxantinguanin fosforibosyltransferase (Szybalska & Szybalski, Proc. Natl. Acad. Sci. USA 48:202 (1992)) og adenin-fosforibosyltransferase(lowy et al., Cell 22: )-gener kan brukes i henholdsvis tk-, hgprt- eller aprt-celler. I tillegg kan antimetabolitt-motstand brukes som grunnlag for valg blant følgende gener: dhfr, som overfører motstand på metotreksat (Wigler et al., Natl. Acad. Sci. USA 77:357 (1980), O'Hare et al., Proc. Natl. Acad. Sci. USA 78:1527 (1981)), gpt, som overfører motstand på mykofenolsyre (Mulligan & Berg, Proc. Natl. Acad. Sci. USA 78:2072 (1981)), neo, som overfører motstand på aminoglykosid G-418 Clinical Pharmacy 12: , Wu and Wu, Biotherapy 3:87-95 (1991), Tolstoshev, Ann. Rev. Pharmacol. Toxicol. 32: (1993), Mulligan, Science 260: (1993) og Morgan and Anderson, Ann. Rev. Biochem. 62: (1993),, TIB TECH 11(5): (May, 1993) og hygro, som overfører motstand på hygromycin (Santerre et al., Gene 30:147 (1984). Metoder som er allment kjent innen faget i rekombinant DNA-teknologi som kan brukes, beskrives i Ausubel et al. (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, NY (1993); Kriegler, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY (1990) og i kapittel 12 og 13, Dracopoli et al. (eds), Current Prolocols in Human Genetics, John Wiley & Sons, NY (1994), Colberre-Garapin et al., J. Mol. Biol. 150:1 (1981). [0296] Uttrykksnivåene til et antistoff-molekyl kan økes med vektorforsterkning (se gjennomgangen Bebbington and Hentschel, The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells in DNA cloning, Academic Press, New York, Vol. 3. (1987)). Når en markør i vektorsystemet som uttrykker antistoffet forsterkes, vil økninger i nivået til hemmeren som finnes i kulturen til vertscellen, øke antall kopier av markørgenet. Fordi det forsterkede området er forbundet med antistoffgenet, øker også produksjonen av antistoffet (Crouse et al., Mol. Cell. Biol. 3:257 (1983)). [0297] In vitro produksjon gjør at oppskaleringen gir store mengder av de ønskede polypeptidene. Metoder for dyrking av pattedyrceller under vevskulturforhold er kjent innen faget og inkluderer homogen suspensjonskultur, f.eks. i en airlift-reaktor eller en kontinuerlig rørereaktor, eller immobilisert eller innkapslet cellekultur, f.eks. i hule fibre, mikrokapsler eller agarose mikrokuler eller keramiske kassetter. Om nødvendig og/eller hvis ønskelig, kan polypeptidløsningen renses med vanlige kromatografimetoder, f.eks. gelfiltrering, ionbyttekromatografi, kromatografi over DEAEcellulose eller (immuno-)affinitetskromatografi, f.eks. etter foretrukket biosyntese av et syntetisk hengelsområdepolypeptid eller før eller etter HIC-kromatografitrinnet som beskrives i dette dokumentet.
117 [0298] Gener som koder Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, kan også uttrykke ikke-pattedyrceller som f.eks. bakterier eller gjær eller planteceller. Bakterier som lett opptar nukleinsyrer, inkluderer medlemmer av enterobacteriaceae som stammer med Escherichia coli eller Salmonella, bacillaceae som Bacillus subtilis, Pneumococcus, Streptococcus og Haemophilus influenzae. I tillegg vil det bli satt pris på at når det uttrykkes i bakterier, blir de heterologe polypeptidene vanligvis en del av inklusjonskroppene. Heterologe polypeptider må isoleres, renses og deretter settes sammen i funksjonelle molekyler. Der tetravalente former av antistoffene ønskes, blir subenheten undermontert i tetravalente antistoffer (WO02/096948A2). [0299] I bakterielle systemer, kan det være en fordel at uttrykksvektorene velges avhengig av hva antistoffet er tenkt å uttrykke. Når f.eks. en stor mengde av et slikt protein skal produseres, for å generere farmasøytiske sammensetninger av et antistoff-molekyl, vektorer som retter uttrykket med høye nivåer av fusjonsproteinprodukter som er lett å rense, kan være ønskelig. Slike vektorer inkluderer, men er ikke begrenset til E. coli uttrykksvektoren pur278 (Ruther et al., EMBO J. 2:1791 (1983)) der den antistoffkodede sekvensen kan ligeres enkeltvis inn i vektoren i rammen med 1acZkodingsområdet slik at fusjonsproteinet produseres, pin-vektorer (Inouye & Inouye, Nucleic Acids Res. 13: (1985), Van Heeke & Schuster, J. Biol. Chem. 24: (1989)) og lignende. pgexvektorer kan også brukes til å uttrykke fremmede polypeptider som fusjonsproteiner med glutation-stransferase (GST). Generelt er slike fusjonsproteiner oppløselige og kan lett renses fra lyserte celler med adsorpsjon og binding til matrise glutationagarose-kuler med etterfølgende eluering i tilstedeværelse av fritt glutation. pgex-vektorene er utformet til å inkludere trombin eller faktor-xa proteasespaltingssteder slik at det klonede målgenproduktet kan frigis fra GST-moieteten. [0300] Eukaryotiske mikrober kan brukes i tillegg til prokaryoter. Saccharomyces cerevisiae eller vanlig bakegjær, er det som er mest vanlig å bruke blant de eukaryotiske mikroorganismene selv om en rekke andre stammer er lett tilgjengelige, f.eks. Pichia pastoris. [0301] Ved uttrykk i Saccharomyces er plasmid YRp7 f.eks. (Stinchcomb et al., Nature 282:39 (1979), Kingsman et al., Gene 7:141 (1979); Tschemper et al., Gene 10:157 (1980)) i vanlig bruk. Dette plasmidet inneholder allerede TRP1-genet som gir et seleksjonsmarkør for en mutant gjærstamme som mangler evnen til å vokse i tryptofan, f.eks. ATCC No eller PEP4-1 (Jones, Genetics 85:12 (1977)). Tilstedeværelse av trp1-lesjon som et karaktertrekk til gjærvertcelle-genomet gir deretter et effektivt miljø for å detektere omdanning ved vekst i uteblivelsen av tryptofan. [0302] I et insektsystem brukes Autographa californica nukleær polyhedrosevirus (AcNPV) vanligvis som vektor for å uttrykke fremmede gener. Viruset vokser i Spodoptera frugiperda-celler. Den antistoffkodende sekvensen kan klones enkeltvis inn i ikke-essensielle områder (f.eks. polyhedringenet) av viruset og plassert under kontroll av en AcNPV-promoteren (f.eks. polyhedrinpromoteren). [0303] Etter at et antistoff-molekyl av offentliggjøringen er blitt rekombinant uttrykt, kan det renses med en hvilken som helst metode som er kjent innen faget for å rense et immunoglobulinmolekyl, f.eks. ved kromatografi (f.eks. ionbytte, affinitet, spesielt ved affinitet med det spesifikke antigenet etter Protein A og graderende kolonnekromatografi), sentrifugering, differensial oppløselighet eller
118 117 ved bruk at en hvilken som helst annen standard metode for å rense proteinene. Som alternativ offentliggjøres en foretrukket metode for å øke affiniteten til offentliggjøringens antistoffer i US A1. 5 VIII. BEHANDLINGSMETODER SOM ANVENDER TERAPEUTISKE Sp35-ANTISTOFFER [0304] Som beskrevet i dette dokumentet, kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse av offentliggjøringen lette NgR1-mediert hemming av aksonal forlengelse som vanligvis skjer i CNS-nevroner. Dette er en fordel i situasjoner der aksonal forlengelse eller nevrittspiring trengs i hjernen eller ryggraden. Ryggradsskade inkludert delvis eller fullstendig knusing eller løsriving, er eksempler på en situasjon der det trengs aksonal forlengelse, men som vanligvis hemmes ved bruk av «Nogo»-banen. Eksempler på sykdommer eller lidelser der aksonal forlengelse og/eller nevrittspiring i hjernen ville være fordelaktig inkludert slag, multippel sklerose og andre nervedegenererende sykdommer og lidelser som multippel sklerose (MS), progressiv multifokal leukoencefalopati (PML), encefalomyelitt (EPL), sentral pontinmyelolyse (CPM), adrenoleukodystrofi, Alexanders sykdom, Pelizaeus Merzbacher-sykdom (PMZ), globoid celleleukodystrofi (Krabbes sykdom) og Wallerisk degenerering, optikusnevritt, transvers myelitt, amylotrofisk-lateralsklerose (ALS), Huntingtons sykdom, Alzheimers sykdom, Parkinsons sykdommer, ryggradsskade, traumatisk hjerneskade, poststrålingsskade, nevrologiske komplikasjoner av kjemoterapi, slag, nevropati, akutt iskemisk optisk nevropati, vitamin E-mangel, isolert vitamin E-mangelsyndrom, AR, Bassen-Kornzweigsyndrom, Marchiafava-Bignami-syndrom, metakromatisk leukodystrofi, trigeminal nevralgi, Bells parese, ryggradsskade og alle nevrologiske sykdommer som er forbundet med nevronal celledød. [0305] I tillegg har oppfinnerne oppdaget at Sp35 uttrykkes i oligodendrocytter og bidrar til oligodendrocyttbiologi. Oppløselige avledninger av Sp35, visse polynukleotider (f.eks. RNAi) samt visse antistoffer som spesifikt bindes til Sp35 som beskrives i dette dokumentet, fungerer som antagonister til Sp35-funksjonen i oligodendrocytter, fremmer spredning, differensiering og overlevelse av oligodendrocytter og fremmer myelinering av nevroner in vitro og in vivo. Dette er en fordel ved sykdommer, lidelser eller tilstander som involverer demyelinering og dysmyelinering. Eksempler på sykdommer og lidelser der oligodendrocyttspredning, differensiering og overlevelse og/eller myelinering eller remyelinering ville være en fordel inkluderer multippel sklerose (MS), progressiv multifokal leukoencefalopati (PML), enkefalomyelitt (EPL), sentral pontin myelolyse (CPM), adrenoleukodystrofi, Alexanders sykdom, Pelizaeus Merzbacher-sykdom (PMZ), globoid celleleukodystrofi (Krabbes sykdom) og Wallerisk degenerasjon, optikusnevritt, transversell myelitt, amyotrofisk-lateralsklerose (ALS), Huntingtons sykdom, Alzheimers sykdom, Parkinsons sykdom, ryggmargsskade, traumatisk hjerneskade, skader etter stråling, nevrologiske komplikasjoner ved kjemoterapi, slag, akutt iskemisk optisk nevropati, vitamin E-mangel, isolert vitamin E-mangel syndrom, AR, Bassen-Kornzweig-syndrom, Marchiafava Bignami-syndrom, metakromatisk leukodystrofi, trigeminal nevralgi og Bells parese. [0306] Følgelig gir ett tilfelle av den gjeldende offentliggjøringen metoder for å behandle ryggradsskade, sykdommer eller lidelser som er forbundet med hemming av nevronal vekst i CNS,
119 sykdommer eller lidelser som er forbundet med hemming av oligodendrocyttvekst eller differensiering og sykdommer som involverer demyelinering eller dysmyelinering av CNS-nevroner i et dyr som lider som et resultat av en slik skade eller sykdom eller predisponert for å få en slik sykdom, der metoden er som omfatter, hovedsakelig består av eller består av å administrere en effektiv mengde Sp35- antistoff eller antigenbindende fragment, variant eller avledning av dem til et dyr. Antistoffene til offentliggjøringen beskrives i dette dokumentet og inkluderer de monoklonale antistoffene som står oppført i tabell 3A og 3B, antistoffer som spesifikt bindes til samme epitop som de monoklonale antistoffene som står oppført i tabell 3A og 3B, antistoffer som kompetitivt hemmer binding av monoklonale antistoffer til Sp35 og som står oppført i tabell 3A og 3B og antistoffer som omfatter polypeptider som er avledet fra monoklonale antistoffer som står oppført i tabell 3A og 3B. [0307] Et terapeutisk Sp35-antistoff som skal brukes ved behandlingsmetoder som offentliggjøres i dette dokumentet, kan prepareres og brukes som terapeutisk middel som fremmer CNS-nevrittutvekt, nevronal overlevelse, aksonstyring og aksonregenerering som fremmer oligodendrocytt-overlevelse, - vekst og/eller differensiering og som fremmer myelinering eller remyelinering av CNS-nevroner. Egenskapene til egnede terapeutiske Sp35-antistoffer inkluderer: binding til Sp35-epitoper som fører til blokkering av Sp35-aktivitet, binding til Sp35 med tilstrekkelig affinitet til å fremkalle en terapeutisk effekt og binding til Sp35, helst til normale bindingspartnere, f.eks. «Nogo»-reseptor. [0308] Terapeutiske Sp35-antistoffer kan være monoklonale, kimæriske eller humaniserte antistoffer eller fragmenter av antistoffer som bindes spesifikt til Sp35. Anstistoffene kan være monovalente, bivalente, polyvalente eller bifunksjonelle antistoffer. Antistoff-fragmenter inkluderer uten begrensning Fab F(ab') 2 - og Fv-fragmenter. [0309] Terapeutiske Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse iht. offentliggjøringen, kan brukes i umerket eller ukonjugert form eller kan koples eller lenkes til legemidler, etiketter eller stabiliseringsmidler som utøver flere terapeutiske effekter eller ikke. [0310] En bestemt dose eller behandlingsplan for en hvilken som helst bestemt pasient vil være avhengig av en rekke faktorer som inkluderer det bestemte Sp35-antistoffet eller antigenbindende fragmentet, variant eller avledning av disse som brukes, pasientens alder, kroppsvekt, generell helsetilstand, kjønn og kosthold og administreringstidspunkt, ekskresjonsraten, legemiddelkombinasjon og alvorlighetsgraden til den bestemte sykdommen som behandles. Vurdering av slike faktorer av medisinske omsorgsytere er innenfor vanlige ferdigheter i faget. Mengden vil også være avhengig av den enkelte pasienten som skal behandles, administreringsrutinen, type formulering, egenskapene til sammensetning som brukes og sykdommens alvorlighetsgrad og den ønskede effekten. Mengden kan bestemmes med farmakologiske og farmakokinetiske prinsipper som er velkjent innen faget. [0311] I metodene til offentliggjøringen, kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse, administreres direkte i nervesystemet, intracerebroventrikulært eller intratekalt, f.eks. inn i en kronisk lesjon av MS som omtalt i mer detalj nedenfor. [0312] I forskjellige tilfeller er Sp35-antistoffet som beskrives ovenfor, en antagonist av Sp35- aktivitet. I visse tilfeller, f. eks. binding av et antagonist Sp35-antistoff til Sp35, om uttrykt på nevroner, blokkerer myelinforbundet hemming av nevrittutvekst eller nevronal celledød. I andre
120 tilfeller blokkerer binding av Sp35-antistoffet til Sp35 slik som uttrykt på oligodendrocytter, hemming av oligodendrocyttvekst eller differensiering eller blokkerer demyelinering eller dysmyelinering av CNS-nevroner. [0313] I metoder av den gjeldende offentliggjøringen av et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det, kan helt bestemt Sp35-antistoffene som beskrives i dette dokumentet, administreres direkte som et utført polypeptid, eller indirekte gjennom en nukleinsyrevektor for å muliggjøre fordelaktig aksonal utvekst, fremme oligodendrocyttspredning, differensiering og overlevelse og/eller fremme myelinering og remyelinering. [0314] I visse tilfeller kan et forsøksobjekt behandles med et nukleinsyre-molekyl som koder et Sp35- antistoff eller antigenbindende fragment, variant eller analog av det, f.eks. i en vektor. Nukleinsyredoser som koder polypeptider er på mellom omtrent 10 ng til 1 g, 100 ng til 100 mg, 1 μg til 10 mg eller μg DNA per pasient. Doser til smittevirus-vektorer varierer fra eller mer, virioner per dose. [0315] I noen tilfeller av den gjeldende offentliggjøringen, administreres et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det i en behandlingsmetode som inkluderer: (1) omdanning eller transfeksjon av en implanterbar vertscelle med en nukleinsyre, f.eks. en vektor som uttrykker et Sp35-antistoff eller et antigenbindende fragment, variant eller avledning av det og (2) implantering av den omdannede vertscellen i et pattedyr på sykdoms-, lidelses- eller skadestedet. Den omdannede vertscellen kan implanteres på stedet til en ryggradsskade eller et dysmyelineringssted. I noen tilfeller av offentliggjøringen, fjernes den implanterbare vertscellen fra et pattedyr, dyrkes midlertidig, omdannes eller transfekteres med en isolert nukleinsyre som koder et Sp35-antistoff og som implanteres tilbake i samme pattedyret den ble fjernet fra. Cellen kan, men trenger ikke, fjernes fra samme stedet som den ble implantert. Slike tilfeller som noen ganger blir kalt ex vivo genbehandling, kan gi kontinuerlig tilførsel av Sp35-polypeptidet som er lokalisert på handlingsstedet i en begrenset tidsperiode. [0316] Metodene for å behandle ryggradsskade, sykdommer eller lidelser forbundet med hemming av nevronal vekst i CNS, sykdommer eller lidelser forbundet med hemming av oligodendrocytt-vekst eller differensiering og sykdommer som involverer demyelinering eller dysmyelinering av CNS-nevroner som omfatter administrering av et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det, testes vanligvis in vitro og deretter in vivo i en akseptabel dyremodell med den ønskede terapeutiske eller profylaktiske aktiviteten før det brukes på mennesker. Egnede dyremodeller inkludert transgeniske dyr, er velkjent hos personer med vanlige ferdigheter i faget, In vitro analyser som f.eks. skal demonstrere den terapeutiske nytten til Sp35-antistoffet som beskrives i dette dokumentet, inkluderer effekten som et Sp35-antistoff har på en cellelinje eller en pasientvevsprøve. Effekten som Sp35-antistoffet har på cellelinjen og/eller vevsprøven kan bestemmes ved bruk av metoder som er kjent hos personer med ferdigheter i faget som f.eks. analysene som offentliggjøres andres steder i dette dokumentet. I henhold til offentliggjøringen, inkluderer in vitro analyser som kan brukes til å bestemme om administrering av et bestemt Sp35-antistoff er indisert, in vitro cellekulturanalyser der en vevsprøve til en pasient dyrkes i kultur og eksponeres for eller at en
121 sammensetning administreres på en annen måte og effekten til en slik sammensetning på vevsprøven observeres. [0317] Supplementære aktive sammensetninger kan også innlemmes i sammensetningene til offentliggjøringen. Et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen, kan f.eks. formuleres sammen og/eller administreres sammen med ett eller flere midler i tillegg. [0318] Offentliggjøringen omfatter alle egnede tilførselsmetoder for et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen til et valgt målvev, inkludert bolusinjisering av en vannholdig løsning eller implantering av et CR (controlled-release)- system. Bruken av et CR-implantat reduserer behovet for gjentatte injeksjoner. IX. FARMASØYTISKE SAMMENSETNINGER OG ADMINISTRERINGSMETODER [0319] Metoder for å preparere og administrere Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, til et forsøksobjekt som trenger dem er velkjent eller lett å bestemme av personer med ferdigheter i faget. Administreringsruten til Sp35- antistoffet eller det antigenbindende fragmentet, variant eller avledning av dem kan f.eks. være oralt, parenteralt ved inhalering eller topisk. Slik begrepet «parenteralt» brukes i dette dokumentet, inkluderes blant annet intravenøs, intraarterial, intraperitoneal, intramuskulær, subkutan, rektal eller vaginal administrering. Selv om alle disse administreringsformene er tydelig overveid som innenfor omfanget av offentliggjøringen, ville en administreringsform være en oppløsning til injisering, helt spesifikt til intravenøs eller intraarteriell injisering eller drypp. Vanligvis kan en egnet farmasøytisk sammensetning til injisering omfatte en buffer (f.eks. acetat, fosfat eller citratbuffer), en surfaktant (f.eks. polysorbat), som alternativ et stabiliserende middel (f.eks. humant albumin), osv. I andre sammenlignbare metoder som det opplyses om i dette dokumentet, kan imidlertid Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem, leveres direkte til stedet til den negative cellepopulasjonen og dermed øke eksponeringen av det syke vevet til det terapeutiske midlet. [0320] Som omtalt tidligere, kan Sp35 antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem, administreres i en farmasøytisk effektiv mengde til in vivo behandling ryggradsskade, sykdommer eller lidelser forbundet med hemming av nevronal vekst i CNS, sykdommer eller lidelser forbundet med hemming av oligodendrocytt-vekst eller differensiering og sykdommer som involverer demyelinering eller dysmyelinering av CNS hos pattedyr. I denne forbindelsen vil det bli satt pris på at de offentliggjorte antistoffene blir formulert slik at de hjelper ved administrering og fremmer stabilitet av det aktive midlet. Det foretrekkes at de farmasøytiske sammensetningene iht. den gjeldende offentliggjøringen, omfatter en farmasøytisk akseptabel, ikke toksisk, steril bærer som f.eks. fysiologisk saltløsning, ikke-toksiske buffere, konserveringsmidler eller lignende. Med tanke på den «instant» søknaden skal den farmasøytiske effektive mengden Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det, konjugert eller ukonjugert, bety en mengde
122 som er tilstrekkelig for å oppnå effektiv binding til et mål og for å oppnå en fordel, f.eks. når det gjelder å bedre symptomene til en sykdom eller lidelse eller for å påvise et stoff eller en celle. [0321] De farmasøytiske sammensetningene som brukes i denne offentliggjøringen omfatter farmasøytisk akseptable bærere inkludert f.eks. ionbyttere, alumina, aluminiumstearat, lecitin, serumproteiner som f.eks. humant serumalbumin, bufferstoffer som f.eks. fosfater, glysin, sorbinsyre, kaliumsorbat, delvise glyseridblandinger av mettede vegetabilske-fettsyrer, vann, salter eller elektrolytter som f.eks. protaminsulfat, dinatriumhydrogenfosfat, kaliumhydrogenfosfat, natriumklorid, sinksalter, kolloidalsilika, magnesiumtrisilikat, polyvinyl pyrrolidon, cellulosebaserte stoffer, polyetylenglykol, natriumkarboksymetylcellulose, polyakrylater, vokser, polyetylenpolyoksypropylen-blokkpolymerer, polyetylenglykol og ullfett. [0322] Preparater til parenteral administrering inkludert sterile vannholdige eller ikke-vannholdige oppløsninger, suspensjoner og emulsjoner. Eksempler på ikke-vannholdige løsemidler er propylenglykol, polyetylenglykol, vegetabilske oljer som olivenolje og injiserbare organiske estere som etyloleat. Vannholdige bærer inkluderer vann, alkoholholdige/vannholdige oppløsninger, emulsjoner eller suspensjoner inkludert saltløsning og bufrede medier. Den gjeldende offentliggjøringen med farmasøytisk akseptable bærere inkluderer, men er ikke begrenset til 0,01 0,1 M og fortrinnsvis 0,05 M fosfatbuffer eller 0,8 % saltløsning. Andre vanlige parenterale vehikler inkluderer natriumfosfatløsninger, Ringers dekstrose, dekstrose og natriumklorid, Ringers laktatoppløsning eller faste oljer. Intravenøse vehikler inkluderer væske- og næringsetterfylling, elektrolyttetterfylling som de som er basert på Ringers dekstrose og lignende. Det kan også finnes konserveringsmidler og andre tilsetningsstoffer som f.eks. antimikrober, antioksidanter, chelateringsmidler og inerte gasser og lignende. [0323] Mer spesifikt inkluderer farmasøytiske sammensetninger som er egnet til injiserbart bruk inkluderer sterile vannholdige oppløsninger (som er vannoppløselige) eller dispergerings- eller sterile pulver til esktemporanøs preparering av sterile injiserbare oppløsninger eller dispergeringer. I slike tilfeller må sammensetningen være steril og skal være flytende i den utstrekningen som lett bruk av sprøyte finnes. Den skal være stabil under produksjons- og oppbevaringsforhold og det foretrekkes at den konserveres mot kontaminerende handlinger av mikroorganismer som bakterier og sopp. Bæreren kan være et løsemiddel eller et dispengeringsmedium som f.eks. inneholder vann, etanol, polyol (f.eks. glyserol, propylenglykol og flytende polyetylenglykol og lignende) og egnede blandinger av dem. Riktig flytegrad kan f.eks. opprettholdes ved å bruke et belegg som f.eks. lecitin, ved å opprettholde den påkrevde partikkelstørrelse når det gjelder dispergering og ved bruk av surfaktanter. Egnede formuleringer til bruk i terapeutiske metoder som offentliggjøres i dette dokumentet beskrives i Remington's Pharmaceutical Sciences, Mack Publishing Co., 16th ed. (1980). [0324] Virkningen av mikroorganismene kan hindres med forskjellige antibakterielle og antifungale midler, f.eks. parabener, klorobutanol, fenol, askorbinsyre, timerosal og lignende. I mange tilfeller vil det være foretrukket å inkludere isotoniske midler, f.eks. sukker, polyalkoholer som mannitol, sorbitol eller natriumklorid i sammensetningen. Forlenget absorpsjon av de injiserbare sammensetningene kan skje ved å inkludere et middel i sammensetningen som forsinker absorpsjon, f.eks. aluminiummonostearat og gelatin.
123 [0325] Uansett kan de sterile injiserbare oppløsningene prepareres ved å innlemme en aktiv sammensetning (f.eks. et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det, alene eller sammen med andre aktive midler) i den påkrevde mengden i et egnet løsemiddel med ett eller en kombinasjon av ingredienser som forklares i dette dokumentet, med påfølgende filtrert sterilisering. Generelt prepareres dispengeringer ved å innlemme den aktive sammensetningen i et sterilt vehikkel som inneholder et grunnleggende dispengeringsmedium og de andre nødvendige ingrediensene fra dem som nevnes ovenfor. Når det gjelder sterile pulver fra preparering av sterile injiserbare oppløsninger, er de fortrukne prepareringsmetodene vakuumtørking og frysetørking som gir et pulver til den aktive ingrediensen samt eventuelt annen ingrediens som ønskes, fra en oppløsning av den som tidligere ble sterilfiltrert. Injiseringspreparatene prosesseres, fylles i beholdere som f.eks. ampuller, poser, flasker, sprøyter eller flasker, og forsegles under aseptiske forhold iht. metoder som er kjent innen faget. I tillegg kan preparatene emballeres og selges som et sett som dem som beskrives i «co-pending» U.S.S.N. 09/259,337 (US A1). Slike produksjonsdeler skal helst ha etiketter eller emballasjevedlegg som indikerer at de assosierte sammensetningene er nyttige ved behandling av et forsøksobjekt som lider av eller er predisponert for autoimmune eller neoplastiske lidelser. [0326] Parenterale formulering kan være en enkel bolusdose, en infusjon eller en start-bolusdose med påfølgende vedlikeholdsdose. Disse sammensetningene kan administreres ved bestemte faste eller variable intervaller, f.eks. én gang hver dag eller etter behov. [0327] Visse farmasøytiske sammensetninger som brukes i denne offentliggjøringen, kan administreres oralt med en akseptabel doseringsform inkludert f.eks. kapsler, tabletter, vannholdige suspensjoner eller oppløsninger. Visse farmasøytiske sammensetninger kan også administreres med nesespray eller inhaleres. Slike sammensetninger kan prepareres som oppløsninger i saltløsninger der benzylalkohol brukes eller andre egnede konserveringsmidler, absorpsjonsfremmere for å forbedre biologisk tilgjengelighet og/eller andre konvensjonelle løseliggjørende eller dispengeringsmidler. [0328] Mengden Sp35-antistoff eller fragment, variant eller avledning av det, som kan kombineres med bærematerialer for å produsere en enkeltdoseform, vil variere avhengig av verten som behandles og den bestemte administreringsmodusen. Sammensetningen kan administrere som en enkeltdose, flere doser eller i en fastslått tidsperiode i en infusjon. Doseringsplaner kan også justeres for å gi den optimale ønskede responsen (f.eks. en terapeutisk eller profylaktisk respons). [0329] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledning av dem, kan administreres til et menneske eller andre dyr iht. de ovennevnte behandlingsmetodene i en mengde som er tilstrekkelig for å produsere en terapeutisk effekt. Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledningene av dem, kan administreres til et slikt menneske eller annet dyr i en konvensjonell doseringsform ved å kombinere antistoffet med en konvensjonell farmasøytisk akseptabel bærer eller fortynner iht. kjente metoder. Det vil bli gjenkjent av en person med ferdigheter i faget at formen og egenskapene til den farmasøytisk aksepterbare bæreren eller fortynneren bestemmes av mengde aktiv ingrediens som den kombineres med og administreringsruten og andre velkjente variabler. Personer med ferdigheter i faget vil i tillegg sette pris på at en
124 «cocktail» som omfatter én eller flere arter av Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem, kan vise seg å være spesielt effektive. [0330] Effektive doser av sammensetningene til den gjeldende offentliggjøringen til behandling av ryggradsskade, sykdommer eller lidelser forbundet med hemming av nevronal vekst i CNS, sykdommer eller lidelser forbundet med hemming av oligodendrocytt-vekst eller differensiering og sykdommer som involverer demyelinering eller dysmyelinering av CNS, varierer avhengig av mange forskjellige faktorer inkludert administreringsmåte, målsted, pasientens fysiologisk tilstand, om pasienten er et menneske eller et dyr, andre medikamenter som er administrert og om behandlingen er profylaktisk eller terapeutisk. Vanligvis er pasienten et menneske, men ikke-humane pattedyr inkludert transgeniske pattedyr, kan også behandles. Behandlingsdosene kan titreres ved bruk av rutinemessige metoder som er kjent hos personer med ferdigheter i faget for å optimalisere sikkerhet og effekt. [0331] Behandling av ryggradsskade, sykdommer eller lidelser forbundet med hemming av nevronal vekst i CNS, sykdommer eller lidelser forbundet med hemming av oligodendrocytt-vekst eller differensiering og sykdommer som involverer demyelinering eller dysmyelinering av CNS med et Sp35- antistoff eller antigenbindende fragment, variant eller avledning av det, doseringen kan f.eks. være mellom 0,0001 til 100 mg/kg og mer vanlig 0,01 til 5 mg/kg (f.eks. 0,02 mg/kg, 0,25 mg/kg, 0,5 mg/kg, 0,75 mg/kg, 1 mg/kg, 2 mg/kg, osv.), av vertens kroppsvekt. Doseringer kan f.eks. være 1 mg/kg kroppsvekt eller 10 mg/kg kroppsvekt eller innen området 1-10 mg/kg, helst minst 1 mg/kg. Doseringer som intermedierer i områdene ovenfor, er også tenkt å være innen omfanget til offentliggjøringen. Forsøksobjekter kan administreres slike doser hver dag, annen hver dag, hver uke eller iht. en hvilken som helst plan som bestemmes med empiriske analyser. Et eksempel på behandling innbefatter administrering av flere doser i en lenger periode, f.eks. i minst 6 måneder. Andre eksempler på behandlingsplaner innbefatter administrering én gang hver to uker eller én gang i måneden eller én gang hver 3. eller 6. måned. Eksempler på doseringsplaner inkluderer 1-10 mg/kg eller 15 mg/kg på påfølgende dager, 30 mg/kg annenhver dag eller 60 mg/kg hver uke. I noen metoder administreres to eller flere monoklonale antistoffer med forskjellige bindingsspesifisiteter samtidig. I slike tilfeller faller doseringen av hvert antistoff som administreres, innenfor de indikerte områdene. [0332] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem kan administreres ved flere anledninger. Intervallene mellom enkeltdoser kan være daglig, ukentlig, månedlig eller årlig. Intervallene kan også være uregelmessige som indikert ved å måle blodnivåene i målpeptidet eller målmolekylet i pasienten. I noen metoder justeres doseringen for å oppnå en plasmapolypeptid-konsentrasjon på μg/ml og i noen metoder μg/ml. Som et alternativ kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem administreres som depotformulering. I et slikt tilfelle kreves mindre hyppig administrering. Dosering og hyppighet varierer avhengig av halveringstiden til antistoffet i pasienten. Halveringstiden til et Sp35-antistoff kan også forlenges ved å fusjonere til et stabilt polypeptid eller moeitet, f.eks. albumin eller PEG. Generelt utviser humaniserte antistoffer lengst halveringstid og deretter kimæriske antistoffer og ikke-humane antistoffer. I ett tilfelle kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem, administreres i ukonjugert form. I et annet tilfelle
125 kan Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledningene av dem av offentliggjøringen, administreres flere ganger i konjugert form. I enda et annet tilfelle kan Sp35- antistoffene eller de antigenbindende fragmentene, variantene eller avledningene av dem av offentliggjøringen, administreres i ukonjugert form og deretter i konjugert form, eller motsatt. [0333] Sammensetningene i den gjeldende offentliggjøringen kan administreres med en hvilken som helst egnet metode, f.eks. parenteralt, intraventrikulært, oralt, med inhaleringsspray, topisk, rektalt, nasalt, bukkalt, vaginalt eller via en implantert beholder. Slik begrepet «parenteralt» brukes i dette dokumentet, inkluderes subkutane, intravenøse, intramuskulære, intraartikulære, intrasynoviale, intrasternale, intratekale, intrahepatiske, intralesjonale og intrakranial injeksjons- eller infusjonsmetoder. Som beskrevet tidligere, virker Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem, i nervesystemet for å fremme overlevelse, spredning og differensiering av oligodendrocytter og myelinering av nevroner og nevronal overlevelse, aksonregenerering og aksonstyring. Dermed administreres Sp35-antistoffene eller der antigenbindende fragmentene, variantene eller avledningene av dem på en slik måte at de krysser blod-hjerne-barrieren i offentliggjøringens metoder. Denne kryssingen kan være et resultat av de fysisk kjemisk-egenskapene knyttet til selve Sp35-antistoff-molekylet, fra andre komponenter i en farmasøytisk formulering eller fra bruken av en mekanisk anordning som f.eks. en nål, en kanyle eller kirurgiske instrumenter for å bryte blod-hjernebarrieren. Der Sp35-antistoffet er et molekyl som ikke naturlig krysser blod-hjerne-barrieren, f.eks. en fusjon til en moietet som bistår ved kryssingen, er f.eks. intratekalt eller intrakranialt, f.eks. inn i en kronisk lesjon ved MS, egnede administreringsruter. Der Sp35-antistoffet er et molekyl som naturlig krysser blod-hjerne-barrieren, kan administreringsruten være én eller flere av de forskjellige rutene som beskrives nedenfor. I noen metoder administreres antistoffene som depotsammensetning eller anordning som f.eks. en Medipad -anordning. Tilførsel over blod-hjerne-barrieren kan forbedres med et bæremolekyl som f.eks. anti-fc-reseptor, transferrin, antiinsulin-reseptor or et toksinkonjugat eller penetreringsenhancer. [0334] Sp35-antistoffene eller de antigenbindende fragmentene, variantene eller avledningene av dem som brukes i offentliggjøringens metoder, kan sprøytes inn i hjernen. Forskjellige implantater til direkte hjerneinfusjon av sammensetninger er kjent og er effektive i å tilføre terapeutiske sammensetninger til humane pasienter som lider av nevrologiske lidelser. Disse inkluderer kronisk infusjon i hjernen ved bruk av en pump, stereotaktisk implantat, midlertidige interstitiale katetre, permanente intrakraniale kateterimplantater og kirurgisk implanterte biologisk nedbrytbare implantater. Se f.eks. et al., «Direct brain infusion of glial cell line-derived neurotrophic factor in Parkinson disease», Nature Med. 9: (2003); Scharfen et al., «High Activity Iodine-12S Interstitial Implant For Gliomas,» Int. J. Radiation Oncology Biol. Phys. 24(4): (1992), Gaspar et al., «Pennanent 125 I Implants for Recurrent Malignant Gliomas,» Int. J. Radiation Oncology Biol. Phys. 43(5): (1999), chapter 66, pages , Bellezza et al., «Stereotactic Interstitial Brachytherapy,» i Gildenberg et al., Textbook of Stereotactic and Functional Neurosurgery, McGraw- Hill (1998) og Brem et al., «The Safety of Interstitial Chemotherapy with BCNU-Loaded Polymer
126 Followed by Radiation Therapy in the Treatment of Newly Diagnosed Malignant Gliomas: Phase I Trial,» J. Neuro-Oncology 26: (1995). [0335] Sammensetningene kan også omfatte et Sp35-antistoff som er dispergert i et biokompatibelt bæremateriale som fungerer som et egnet tilførsels- eller støttesystem til sammensetningene. Egnede eksempler på depotbærere inkluderer semipermeable polymermatriser som er utformet som f.eks. suppositorium eller kapsler. Implanterbare eller mikrokapsulære depotmatriser inkludert polylaktider (amerikansk patentnr. 3,773,319, EP 58,481), kopolymerer av L-glutaminsyre og gamma-etyl-lglutamat (Sidman et al., Biopolymers 22: (1985)); poly(2-hydroksyetyl-metakrylat), etylenvinylacetat (Langer et al., J. Biomed. Mater. Res. 15: (1981), Langer, Chem. Tech. 12: (1982)) eller poly-d-(-)-3hydroksybutyrsyre (EP 133,988). [0336] I noen tilfeller av offentliggjøringen, administreres et Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det av offentliggjøringen, til en pasient med direkte infusjon inn i det aktuelle området i hjernen. Se f.eks. Gill et al., ovenfor. Alternative metoder er tilgjengelige og kan brukes for å administrere et Sp35-antistoff iht. offentliggjøringen. Stereotaktisk plassering av et kateter eller implantat kan f.eks. gjøres ved bruk av Riechert-Mundinger-enheten og ZD (Zamorano- Dujovny) flerbruks-lokaliseringsenheten. Kontrastforbedret computertomografi (CT)-skanning, injisering av 120 ml omnipakk, 350 mg jod/ml med 2 mm snittykkelse kan muliggjøre tredimensjonal multiplan behandlingsplanlegging (STP, Fischer, Freiburg, Tyskland). Dette utstyret muliggjør planlegging basert på magnetisk resonansavbildnings-studier, fletter sammen CT- og MRImålinformasjon til tydelig målbekreftelse. [0337] Leksell stereotaktisk system (Downs Surgical, Inc., Decatur, GA, USA) modifisert til bruk med en GE CT-skanner (General Electric Company, Milwaukee, WI, USA) samt Brown Roberts-Wells (BR W) stereotaktisk system (Radionics, Burlington, MA, USA) kan brukes til dette. Dermed kan den runde bunnringen på BRW stereotaktiske ramme festes til pasientens skalle på morgenen til implantatet. Serie-CT-deler kan skaffes ved 3 mm intervaller gjennom (målvev)-området med en grafittstanglokalisator som spennes til bunnplaten. Et dataprogram til behandlingsplanlegging kan kjøres et en VAX 11/780-datamaskin (Digital Equipment Corporation, Maynard, MA, USA) ved bruk av CTkoordinatorene til grafittstang-avbildninger for å kartlegge mellom CT-mellomrommet og BRWmellomrommet. [0338] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, kan som et alternativ administreres i kombinasjon med andre midler som er effektive i å behandle lidelsen eller tilstanden som krever behandling (f.eks. profylaktisk eller terapeutisk). 35 X. DIAGNOSTIKK [0339] I tillegg gir offentliggjøringen en diagnostisk metode som er nyttig ved diagnose av nevronale lidelser eller skader som involverer måling av uttrykksnivået til Sp35-proteinet eller transkript i vev eller andre celler eller kroppsvæske fra et forsøksobjekt eller sammenligning av målt uttrykksnivå
127 med standard Sp35-uttrykksnivåer i normalt vev eller normal kroppsvæske der en økning i uttrykksnivået sammenlignet med standarden, er en indikasjon på en lidelse. [0340] Sp35-spesifikke antistoffer kan brukes til å analysere proteinnivåer i en biologisk prøve ved bruk av klassiske immunohistologiske metoder som er kjent hos personer med ferdigheter i faget (f.eks. se Jalkanen, et al., J. Cell. Biol. 101: (1985), Jalkanen, et al., J. Cell Biol. 105: (1987)). Andre antistoff-baserte metoder som er nyttig ved påvisning av proteinuttrykk inkluderer immunoanalyser som f.eks. en enzymkoblet immunosorbentanalyse (ELISA), immunopresipitering eller Western blot-metoden. Egnede analyser beskrives i mer detalj et annet sted i dette dokumentet. [0341] «Analysere uttrykksnivået til Sp35-polypeptid» er ment å bety kvalitativt eller kvantitativt måling eller estimering av Sp35-polypeptidnivået i en første biologisk prøve enten direkte (f.eks. ved å bestemme eller estimere absolutt proteinnivå) eller relativt (f.eks. ved å sammenligne med det kreftassosierte polypeptidnivået i en andre biologisk prøve). Det foretrekkes at Sp35-polypeptiduttrykksnivået i den første biologiske prøven måles eller estimeres og sammenlignes med et standard Sp35-polypeptidnivå, der standarden kommer fra en andre biologisk prøve som skaffes fra et forsøksobjekt uten lidelsen eller som bestemmes basert på å finne middelverdien til nivåene fra en førsøkspopulasjon som ikke har lidelsen. Det vil også bli satt pris på innen faget at etter at «standard» Sp35-polypeptidnivået er kjent, kan det brukes gjentatte ganger som sammenligningsstandard. [0342] «Biologisk prøve» er ment å bety en hvilken som helst biologisk prøve som skaffes fra et forsøksobjekt, cellelinje, vevskultur eller annen cellekilde som muligens uttrykker Sp35. Metoder for å skaffe vevsbiopsier og kroppsvæsker fra pattedyr, er velkjent innen faget. [0343] Sp35-antistoffer til bruk i diagnostiske metoder som beskrives ovenfor, inkluder alle Sp35- antistoffer som spesifikt bindes til et Sp35-genprodukt som beskrevet et annet sted i dette dokumentet. XI. IMMUNOANALYSER [0344] Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av disse av offentliggjøringen, kan analyseres for immunospesifikk binding med en hvilken som helst metode som er er kjent innen faget. Immunoanalysene som kan brukes, kan også inkludere, men er ikke begrenset til kompetitive og ikke kompetitive analysesystemer ved bruk av slike metoder som Western blot, radioimmunoanalyser, ELISA (enzymkoplet immunosorbentanalyse), «sandwich» immunoanalyser, immunopresipiteringsanalyser, presipitinreaksjoner, geldiffusjons-presipitinreaksjoner, immunodiffusjonsanalyser, agglutinasjonsanalyser, komplement-fikseringsanalyser, immunoradiometriske analyser, fluorescens-immunoanalyser, protein A-immunoanalyser for å nevne noen. Slike analyser er rutinemessige og velkjent innen faget (se f.eks. Ausubel et al., eds, Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, Vol. 1 (1994). Eksempler på immunoanalyser beskrives i korthet nedenfor (men er ikke tent å være begrensende). [0345] Immunopresipiteringsprotokoller omfatter generelt lysering av en cellepopulasjon i en lysisbuffer som f.eks. RIPA-buffer (1 % NP-40 eller Triton X-100, 1 % natriumdeoksykolat, 0,1 % SDS,
128 ,15 M NaCl, 0,01 M natriumfosfat ved ph 7,2, 1 % Trasylol) supplementert med proteinfosfatase og/eller proteasehemmere (f.eks. EDTA, PMSF, aprotinin, natriumvanadat), tilsetting av antistoffet av interesse til cellelysatet, inkubering i en tidsperiode (f.eks. 1-4 timer) ved 4 C, tilsetting av protein A og/eller protein G-sefarosekuler til cellelysatet, inkubering i omtrent en time eller med ved 4 C, vasking av kulene i lysebuffer og resuspendering av kuler i SDS/prøvebuffer. Evnen som antistoffet av interesse har for å immunopresipitere et bestemt antigen, kan f.eks. analyseres med en Western blotanalyse. En person med ferdigheter i faget vil kjenne til hvilke parametere som kan modifiseres for å øke bindingen av antistoffet til et antigent og redusere bakgrunnen (før fjerning av cellelysat med sefarosekuler). For mer omtale i forbindelse med immunopresipiteringsprotokoller se f.eks. Ausubel et al., eds, Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, Vol. 1 (1994) at [0346] Western blot-analyse omfatter generelt preparering av proteinprøver, elektroforese av proteinprøver i et polyakrylamidgel (f.eks. 8 %-20 % SDS-PAGE avhengig av antigenets molekylvekt), overføring av proteinprøven fra polyakrylamidgelet til en membran som f.eks. nitrocellulose, PVDF eller nylon, blokkering av membranen i blokkeringsoppløsningen (f.eks. PBS med 3 % BSA eller fettfri melk), vasking av membranen i vaskebuffer (f.eks. PBS-Tween 20), blokkering av membranen med primært antistoff (antistoffet av interesse) fortynnet i blokkeringsbuffer, vasking av membranen i vaskebufferen, blokkering av membranen med et sekundært antistoff (som gjenkjenner det primære antistoffet, f.eks. et antihumant antistoff) som er konjugert til et enzymatisk substrat (f.eks. pepperrot-peroksidase eller alkalisk fosfatase) eller radioaktivt molekyl (f.eks. 32p eller 1251) som er fortynnet i blokkeringsbuffer, vasking av membranen i vaskebuffer og påvisning av antigen. En person med ferdighet i faget vil kjenne til parametrene som kan modifisere for å øke det påviste signalet og for å redusere bakgrunnsstøy. For mer omtale i forbindelse med Western blot-protokoller se f.eks. Ausubel et al., eds, Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York Vol. 1 (1994) at [0347] ELISA-er omfatter å preparere antigen, belegge brønnen til en 96-brønnmikrotiterplate med antigenet, tilsette antistoffet av interesse som er konjugert til en detekterbar sammensetning som f.eks. et enzymatisk substrat (f.eks. pepperrot-peroksidase eller alkalisk fosfatase), til en brønn og inkuberes i en tidsperiode, og påvise antigenet. I ELISA-er trenger antistoffet av interesse ikke å konjugeres til en detekterbar sammensetning, i stedet kan et andre antistoff (som gjenkjenner antistoffet av interesse) som konjugeres til en detekterbar sammensetning, tilsettes til brønnen. I tillegg, istedenfor å belegge brønnen med antigenet, kan brønnen belegges med antistoffet. I dette tilfellet kan et andre antistoff som konjugeres til en detekterbar sammensetning, tilsettes etter at antigenet av interesse tilsettes til den belagte brønnen. En person med ferdigheter i faget vil kjenne til parametrene som kan modifiseres for å øke signalet som detekteres samt andre variasjoner av ELISA-er som er kjent innen faget. For videre omtale om ELISA-er se Ausubel et al., eds, Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, Vol. 1 (1994) at [0348] Bindingsaffiniteten til et antistoff til et antigen og av-raten til antistoff-antigen-interaksjonen, kan bestemmes med kompetitive bindingsanalyser. Ett eksempel på en kompetitiv bindingsanalyse er en radioimmunoanalyse som omfatter inkubering av et merket antigen (f.eks. 3 H or 125 I) med
129 antistoffet av interesse der det finnes økende mengder umerket antigent, og påvisning av antistoff som er bundet til det merkede antigenet. Affiniteten til antistoffet av interesse for et bestemt antigen og binding av av-rater, kan bestemmes fra data med Scatchard-plottanalyse. Konkurranse med et andre antistoff kan også bestemmes ved å bruke radioimmuonoanalyser. I dette tilfellet inkuberes antigenet med antistoff av interesse som konjugeres til en merket sammensetning (f.eks. 3 H or 125 I) der det finnes økende mengder av et umerket andre antistoff. [0349] I tillegg kan Sp35-antistoffer eller antigenbindende fragmenter, varianter eller avledninger av dem av offentliggjøringen, brukes histologisk som i immunofluorescens, immunoelektronmikroskopi eller ikke-immunologiske analyser til in situ påvisning av kreftantigengen-produkter eller konserverte varianter eller peptidfragmenter av disse. In situ påvisning kan utføres ved å fjerne en histologisk prøve fra en pasient og påføre den på et merket Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det, og helst påføre med overlegg av det merkede antistoffet (eller fragment) på den biologiske prøven. Ved bruk av en slik prosedyre er det mulig å bestemme ikke bare om det finnes Sp35-protein eller konserverte varianter eller peptidfragmenter, men også om fordelingen i det undersøkte vevet. Ved bruk av den gjeldende offentliggjøringen, kan personer med vanlige ferdigheter i faget lett forstå at en rekke forskjellige histologiske metoder (som f.eks. fargingsprosedyrer) kan modifiseres for å oppnå slik in situ påvisning. [0350] Immunoanalyser og ikke-immunoanalyser av Sp35-genprodukter eller konserverte varianter eller peptidfragmenter av disse, vil vanligvis omfatte inkubering av en prøve som f.eks. en biologisk væske, vevsekstrakt, friske høstede celler eller lysater av celler som er inkubert i cellekultur der det finnes merket antistoff som kan bindes til Sp35 eller konserverte varianter eller peptidfragmenter av disse og detektere det bundne antistoffet med en hvilken som helst av de mange metodene som er kjent innen faget. [0351] Den biologiske prøven kan anbringes i kontakt med og immobilisert på et fastfase-støtte eller bærer som f.eks. nitrocellulose eller annen fast støtte som kan immobilisere celler, cellepartikler eller oppløselige proteiner. Deretter kan støtten vaskes med egnede buffere med påfølgende behandling med detekterbart merket Sp35-antistoff eller antigenbindende fragment, variant eller avledning av det. Fastfase-støtten kan deretter vaskes med bufferne for andre gang for å fjerne ubundet antistoff. Deretter kan man velge å merke antistoffet. Mengden bundet merke på fast støtte kan deretter påvises på konvensjonell måte. [0352] «Fastfase-støtte eller bærer» er ment å bety en hvilken som helst støtte som er i stand til å binde et antigen eller et antistoff. Velkjente støtter eller bærere inkluderer glass, polystyren, polypropylen, polyetylen, dekstran, nylon, amylaser, naturlige og modifiserte celluloser, polyakrylamider, gabbro og magnetitt. Av natur kan bæreren være enten oppløselig en en viss grad eller uoppløselig når det gjelder den gjeldende offentliggjøringen. Støttematerialet kan ha nesten en hvilken som helst mulig strukturell konfigurasjon så lenge som det koplede molekylet kan bindes til et antigen eller antistoff. Dermed kan støttekonfigurasjonen være sfærisk som en kule, eller sylindrisk som innsiden av et prøverør eller den utvendige overflaten på en stang. Som alternativ kan flaten være flatt som et ark, et prøvepapir, osv. Foretrukne støtter inkluderer polystyrenkuler. Personer
130 med ferdigheter i faget vil kjenne til andre egnede bærer for å binde antistoff eller antigen, eller vil kunne fastslå dette ved å bruke rutinemessig forsøk. [0353] Bindingsaktiviteten til et bestemt parti med Sp35-antistoff eller antigenbindende fragment, variant aller avledning av det kan bestemmes iht. velkjente metoder. Personer med ferdigheter i faget vil kunne bestemme operative og optimale analyseforhold for hver bestemmelsen ved å bruke rutinemessig forsøk. [0354] Det finnes en rekke forskjellige metoder om er tilgjengelig, for å måle affiniteten til en antistoff-antigen-interaksjon, men relativ få for å bestemme hastighetskonstanter. De fleste metodene er avhengige enten av merket antistoff eller antigen. Det er uunngåelig at dette kompliserer rutinemessige målinger og innfører uvissheter i de målte kvantitetene. [0355] Overflate-plasmonreasonans (SPR) som utført i BIAcore gir en rekke fordeler i forhold til konvensjonelle metoder for å måle affiniteten til antistoff-antigen-interaksjoner: (i) ingen krav om å merke verken antistoffet eller antigenet, (ii) trenger ikke å rense antistoffene på forhånd, cellekultursupernatant kan brukes direkte, (iii) sanntidsmålinger muliggjør rask semikvantitativ sammenligning av forskjellige monoklonale antistoff-interaksjoner, aktiveres og er tilstrekkelig ved mange evalueringsoppgaver, (iv) biospesifikk flate kan regenereres slik at en rekke forskjellige monoklonale antistoffer kan lett sammenlignes under like forhold, (v) analytiske prosedyrer er helautomatiske og en utstrakt rekke med målinger kan utføres uten brukerintervensjon. BIAapplications Handbook, version AB (reprinted 1998), BIACORE code No. BR ; BIAtechnology Handbook, version AB (reprinted 1998), BIACORE code No. BR [0356] SPR-baserte bindingsstudier krever at ett medlem av et bindingspar immobiliseres på en sensorflate. Den immobliserte bindingpartneren kalles liganden. Bindingpartneren i oppløsningen kalles analytten. I noen tilfeller er liganden festet indirekte til flaten via binding til et annet immobilisert molekyl som kalles fangemolekyl. SPR-respons reflekterer en endring i massekonsentrasjonen på detektorflaten i det analyttene binder eller dissosieres. [0357] Sanntids BIAcore-målinger overvåker interaksjoner direkte i det de skjer basert på SPR. Metoden er godt egnet til bestemmelse av kinetiske parametere. Komparativ affinitetsrangering er ekstremt enkelt å utføre, og både kinetiske og affinitetskonstanter kan avledes fra sensorgramdataene. [0358] Når analytt injiseres i en diskret puls over en ligandflate, and det resulterende sensorgrammet deles inn i tre essensielle faser: (i) assosiasjon av analytt med ligand under prøveinjisering, (ii) likevekt eller stø tilstand under prøveinjiseringen, der analyttbindingsraten balanseres med dissosiasjon fra komplekset, (iii) dissosiasjon av analytt fra flaten under bufferstrømning. [0359] Assosiasjons- og dissosiasjonsfasen gir informasjon om kinetikken til analytt-ligandinteraksjonen (k a og k d, ratene til kompleksformasjon og dissosiasjon, k d /k a = K D ). Likevektsfasen gir informasjon om affiniteten til analytt-ligand-interaksjonen (K D ). [0360] BIAevaluation-programvaren gir omfattende hjelp ved kurvetilpasning ved bruk av både numerisk integrasjon og globale tilpasningsalgoritmer. Med egnet analyse av dataene, kan separate hastighets- og affinitetskonstanter skaffes fra enkle BIAcore-undersøkelser. Det målbare affinitetsområdet med denne metoden er et svært stort område som strekker seg fra mm til pm.
131 [0361] Epitopspesifisitet er en viktig egenskap til et monoklonalt antistoff. Epitopkartlegging med BIAcore, i motsetning til konvensjonelle metoder med bruk av radioimmunoanalyse, ELISA elle andre flateadsorpsjonsmetoder, krever ikke merking eller rensing av antistoffer, og muliggjør spesifisitetstesting på flere steder ved bruk av en sekvens til flere monoklonale antistoffer. I tillegg kan et stort antall analyser behandles automatisk. [0362] Forsøk med parvis binding tester evnen som to MAb-er har til å bindes samtidig til samme antigen. MAb-er som rettes mot separate epitoper, bindes hver for seg, mens MAb-er som rettes mot identiske eller nært relaterte epitoper, forstyrrer hverandres binding. Disse bindingsforsøkene med BIAcore er rettfrem å utføre. [0363] Et fangemolekyl kan f.eks. brukes til å binde den første Mab-en og deretter tilsette antigen og den andre Mab-en sekvensielt. Sensorgrammene åpenbarer: 1. hvor mye antigent bindes til første Mab-en, 2. i hvilken utstrekning den andre Mab-en bindes til den overflatefestede antigenet, 3. hvis den andre Mab-en ikke bindes, om reversering av rekkefølgen til de parvise testene endrer resultatene. [0364] Peptidhemming er en annen metode som brukes ved epitopkartlegging. Denne metoden kan komplementere studier av parvis antistoffbinding og kan relaterer funksjonelle epitoper til strukturelle funksjoner når den primære sekvensen til antigenet er kjent. Peptider eller antigenfragmenter testes for hemming av bindingen av forskjellige Mab-er til immobilisert antigen. Peptider som forstyrrer bindingen til en bestemt Mab, antas å være strukturelt forbundet med epitopen som defineres av den Mab-en. [0365] Praksisen som den gjeldende offentliggjøringen vil bruke, med mindre noe annet er indisert, konvensjonelle cellebiologi-metoder, cellekultur, molekylbiologi, transgenisk biologi, mikrobiologi, rekombinant DNA og immunologi som er innenfor ferdigheten til faget. Slike metoder forklares fullt ut i litteraturen. Se f.eks. Molecular Cloning A Laboratory Manual, 2nd Ed., Sambrook et al., ed., Cold Spring Harbor Laboratory Press: (1989); Molecular Cloning: A Laboratory Manual, Sambrook et al., ed., Cold Springs Harbor Laboratory, New York (1992), DNA Cloning, D. N. Glover ed., Volumes I and II (1985); Oligonucleotide Synthesis, M. J. Gait ed., (1984), Mullis et al. amerikansk patentnr.: 4,683,195; Nucleic Acid Hybridization, B. D. Hames & S. J. Higgins eds. (1984), Transcription And Translation, B. D. Hames & S. J. Higgins eds. (1984); Culture of Animal Cells, R.I. Freshney, Alan R. Liss, Inc., (1987); Immobilized Cells And Enzymes, IRL Press, (1986), B. Perbal, A Practical Guide To Molecular Cloning (1984), thesis, Methods In Enzymology, Academic Press, Inc., N.Y., Gene Transfer Vectors For Mammalian Cells, J. H. Miller and M. P. Calos eds., Cold Spring Harbor Laboratory (1987), Methods In Enzymology, Vols. 154 and 155 (Wu et al. eds.), Immunochemical Methods In Cell And Molecular Biology, Mayer and Walker, eds., Academic Press, London (1987), Handbook of Experimental Immunology, Volumes I-IV, D. M. Weir and C. C. Blackwell, eds., (1986), Manipulating the Mouse Embryo, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., (1986) og i Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1989). [0366] Generelle prinsipper ved antistoff-engineering fremlegges i Antibody Engineering, 2nd edition, C.A.K. Borrebaeck, Ed., Oxford Univ. Press (1995). Generelle prinsipper ved protein-engineering fremlegges i Protein Engineering, A Practical Approach, Rickwood, D., et al., Eds., IRL Press at Oxford
132 Univ. Press, Oxford, Eng. (1995). Generelle prinsipper ved antistoffer og antistoff-haptenbinding fremlegges i: Nisonoff, A, Molecular Immunology, 2nd ed., Sinauer Associates, Sunderland, MA (1984) og Steward, M.W., Antibodies, Their Structure and Function, Chapman and Hall, New York, NY (1984). I tillegg følges standard metoder i immunologi som er kjent innen faget og som ikke spesifikt beskrives, generelt som i Current Protocols in Immunology, John Wiley & Sons, New York; Stites et al. (eds), Basic and Clinical-Immunology (8th ed.), Appleton & Lange, Norwalk, CT (1994) og Mishell and Shiigi (eds), Selected Methods in Cellular Immunology, W.H. Freeman and Co., New York (1980). [0367] Standard referanselitteratur som fremlegger generelle prinsipper ved immunologi inkluderer Current Protocols in Immunology, John Wiley & Sons, New York; Klein. J., Immunology: The Science of Self-Nonself Discrimination, John Wiley & Sons, New York (1982), Kennett, R., et al., eds., Monoclonal Antibodies, Hybridoma: A New Dimension in Biological Analyses, Plenum Press, New York (1980), Campbell, A., «Monoclonal Antibody Technology» i Burden, R., et al., eds., Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 13, Elsevere, Amsterdam (1984), Kuby Immunnology 4 th ed. Ed. Richard A. Goldsby, Thomas J. Kindt and Barbara A Osborne, H. Freemand & Co. (2000), Roitt, I., Brostoff. J. og Male D., Immunology 6 th ed. London: Mosby (2001), Abbas A., Abul, A and Lichtman, A., Cellular and Molecular Immunology Ed. 5, Elsevier Health Sciences Division (2005), Kontermann and Dubel, Antibody Engineering, Springer Verlan (2001), Sambrook and Russell, Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Press (2001), Lewin, Genes VIII, Prentice Hall (2003), Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Press (1988), Dieffenbach and Dveksler, PCR Primer Cold Spring Harbor Press (2003). EKSEMPLER 25 EKSEMPEL 1 Sp35 er involvert i oligodendrocyttbiologi [0369] Oligodendrocytter modner gjennom flere utviklingsstadier fra A2B5-progenitorceller (som uttrykker A2B5), differensiering til premyelinerings- oligodendrocytter (som uttrykker O1 og O4) og til slutt til modne myelinerings-oligodendrocytter (som uttrykker O1, O4 og MBP). Dermed, ved å overvåke tilstedeværelsen eller uteblivelsen av A2B5-, O1-, O4- og MBP-markører, er det mulig å bestemme utviklingsstadiet til en bestemt celle og å evaluere rollen til Sp35-Fc i oligodendrocyttbiologi. For å finne en generell gjennomgang om oligodendrocyttbiologi se f.eks. Baumann and Pham-Dinh, Physiol. Rev. 81: (2001). [0370] Monoklonale antistoffer mot O4, MBP og CNPase var fra Sternberger Monoklonaler, antistoff til APC (klon CC-1, ref. 29) var fra Calbiochem. Andre antistoffer var βiii tubulin (Covance), Sp35 (Biogen Idec), Fyn (Santa Cruz Biotechnology) og Phospho-Fyn (Biosource). Monoklonale antistoffer mot A2B5 er tilgjengelige hos Chemicon. 40 Sp35 uttrykkes i oligodendrocytter
133 [0371] Uttrykk av Sp35 i renset P13 CG-nevron fra rotte, P2-oligodendrocytter og P4-astrocyttkulturer ble analysert med polymerasekjedereaksjon etter revers transkripsjon (RT PCR). Et sett fra Ambion, Inc. ble brukt til å ekstrahere mrna fra rottehjerneceller iht. produsentens instruksjoner. Semikvantitativ RT-PCR som ble utført ved bruk av primer i starten 5' AGAGACATGCGATTGGTGA 3' (SEQ ID NR:344) og primer på slutten 5' AGAGATGTAGACGAGGTCATT 3' (SEQ ID NR:345), utviste høyt uttrykk i nevroner, lavt uttrykk i oligodendrocyttet og igjen uttrykk i astrocytter. [0372] Uttrykk av Sp35 i oligodendrocytter ble bekreftet med in situ hybridisering i seksjoner som var avledet fra synsnerven til en voksen rotte. Synsnerveseksjoner fra rotter ble preparert og behandlet som beskrevet i Mi et al., «Sp35 is a component of the Nogo-66 receptor/p75 signaling complex,» Nat. Neurosci. 7: (2004) og sondert med digoksigeninmerket Sp35-antisense- eller sense- RNA-er ved bruk av de første 500 nukleotidene til Sp35-kodesekvensen. Seksjonene ble farget iht. produsentens instruksjoner ved bruk av settet Tyrarnide Signal Amplification (Amersham Biosciences) og et fluorescent anti-digoksigeninkonjugert antistoffsett (Perkin Elmer). Ved kombinerte in situ og immunofluorescens-analyser ble seksjonene først sondert med digoksigenin-merkede RNA-er og deretter med antistoffer, f.eks. CC1-antistoff (Calbiochem, en markør til modne oligodendrocytter) or anti-sp35-antistoff. Vi observerte at oligodendrocytter som hybridisert til en antisense-sp35-probe også ble kofarget med et antistoff til CC1 (dataene vises ikke). Ingen spesifikk merking ble observert ved bruk av sense Sp35-probe. Sp35-uttrykk i oligodendrocytter ble også bekreftet med immunohistokjemi-studier av vevsseksjoner fra det laterale ventrikkelområdet til P7-rottekorteks. Flertallet av de kortikale cellene som ble merket med CC1-antistoffet, ble også merket med anti- Sp35-antistoffet. Dataene vises ikke. Spesifisiteten til interaksjonen ble bekreftet med preadsorpsjon av anti-sp35-antistoffet med Sp35-Fc (se eksempel 2) som eliminerte signalet Sp35-spesifikk RNAi-«knockdown» av Sp35-uttrykk fremmer oligodendrocytt-vekst og differensiering. [0373] Sp35-spesifikk RNAi ble brukt til å fjerne Sp35-uttrykk i oligodendrocytt-prekursorceller for å undersøke hvordan Sp35 bidrar til oligodendrocytt-vekst og differensiering A2B5- oligodendrocytt-prekursorceller ble infisert med lentivirusbærende Sp35-spesifikk RNAi-sekvens eller kontroll-rnai preparert på følgende måte. [0374] Murine og Sp35-DNA-sekvenser fra rotter ble sammenlignet for å finne homologe områder til bruk som kandidater ved sh-rna-er(shrna). CH324, til lentivirus-uttrykk av Sp35 RNAi, ble konstruert ved festing av oligonukleotidene LV1-035 og LV1-036 og ligering til HpaI- og XhoI-nedbrutt pll3.7. pll3.7-vektoren, tilleggs-metodologi og virusproduksjon var slik som beskrevet i Rubinson et al., Nat. Genet. 33, (2003). Sp35 RNAi-oligonukleotider ble kjøpt fra MWG og har følgende sekvenser: LV1 035 (sense-oligo) 5'-TGA TCG TCA TCC TGC TAG ACT TCA AGA GAG TCT AGC AGG ATG ACG ATC TTT TTT C-3' (SEQ 1D NR:346) og LV1-036 (antisense-oligo) 5' TCG AGA AAA AAG ATC GTC ATC CTG CTA GAC TCT CTT GAA GTC TAG CAG GAT GAC GAT CA-3' (SEQ ID NR:347). [0375] Kontroll-RNAi ble utformet med samme oligonukleotid-sekvenser unntatt for nukleotidendringer som indikeres med små bokstaver: 5'-TGA TCc TCA TcC ttc Tat ACT TCA AGA GAG
134 TgT AGC AGG ATG AcG ATC TTT ITT CTC GA - 3' (SEQ ID NR:348) og 5' TCG AGA AAA AAG ATC GTC ATe CTG CTA GAC TCT CIT GAA GTa TAG aag GAT GAC GAT CA-3'. (SEQ IDNO:349). [0376] Før lentiviruset produseres, ble DNA fra pll3.7 eller kandidat-shrna i pll3.7 kotransfektert med Sp35-HA-merket plasmid i forholdet 5 til 1 inn i CHO-celler i et 6-brønns format. «Knockdown» ble analysert med Western blot-deteksjon av Sp35-HA-tagg fra transfekterte CHO-cellelysater samt med Northern blot av total RNA som var preparert fra duplikate brønner. Blot-en ble sondert med et Sp35 cdna-fragment. Analyser ble utført 48 timer etter transfeksjon. Som forventet var det en 10-fold reduksjon i Sp35 mrna i CH324 RNAi-behandlede CHO-celler i forhold til kontrollbehandlede celler. Dataene vises ikke. RNAi-lentiviruser som bærer grønt fluorescensprotein (GFP), ble generert som beskrevet i Rubinson et al. I kulturer behandlet enten med kontroll eller Sp35 RNAi, var omtrent 80 % av oligodendrocyttene GFP-positive. Det totale celleantall ble ikke endre av RNAi behandlingene. For å kvantitifisere effektene av RNAi på differensiering ble kun GFP-uttrykkende oligodenrodcytter telt. [0377] Berikede oligodendrocyttpopulasjoner ble dyrket fra Long Evans P2-hunnrotter som beskrevet av Conn, Meth. Neurosci. 2:1-4 (Academic Press; 1990) med følgende modifikasjoner. I korthet ble forhjernen dissekert og plassert i Hank's buffered salt solution (HBSS; Invitrogen). Vevet ble skjært inn i 1 mm fragmenter og inkubert ved 37 C i 15 minutter i 0,01 % trypsin og 10 μg/ml Dnase. Dissosierte celler ble overtrukket på poly-l-lysin belagt T75-vevskulturflasker og dyrket ved 37 C i 10 dager i et DMEM-medium med 20 % fetalt kalveserum (Invitrogen). Oligodendrocytt-prekursorer (A2B5 + ) ble samlet ved å riste flaske over natten ved 200 rotasjoner per minutt ved 37 C som førte til en 95 % ren populasjon. Kulturene ble holdt i et høyglukose DMEM (Dulbecco's modified Eagle's medium) med FGF/PDGF (10 ng/ml, Peprotech) i én uke. Fjerning av FGF/PDGF gjorde at A2B5 + - cellene kunne differensiere inn i O4 + -premyelinerings-oligodendrocytter etter 3-7 dager og differensiere inn i O4 + og MBP + modne oligodendrocytter etter 7-10 dager. Disse differensieringstilstandene er svært opplagte fra endringene i morfologi: A2B5 + celler har bipolar fasong, O4 + premyelinerings-oligodendrocytter har lengre og mer forgrenede prosesser og MBP + modne oligodendrocytter inneholder myelinskjedekonstruksjoner mellom prosessene. [0378] A2B5-oligodendrocytt-prekursorceller ble infisert med lentiviruset som inneholdt CH324 RNAi. De resulterende cellene ble dyrket i 3 dager og antall O4-positive (en markør for oligodendrocyttdifferensiering) oligodendrocytter ble telt. Endogen Sp35-uttrykk ble redusert med infeksjonen med Sp35 RNAi-lentiviruset og ble bekreftet med RT-PCR. Reduksjonen av Sp35 førte til mer differensierte, modne oligodendrocytter i forhold til kontrollinfiserte celler. Dette var tydelig på grunn av økningen i lengden på celleprosessene og at det fantes rikelig med myelinskjedekonstruksjoner (dataene vises ikke). I cellene som uttrykker Sp35 RNAi, fantes det tre ganger så mange modne (O4-positive) oligodendrocytter som i kontrollkulturene. Disse dataene indikerer at Sp35 kan regulere oligodendrocytt-differensieringen negativt. Dominant-negative Sp35 fremmer oligodendrocytt-vekst og differensiering. 40 [0379] Lentivirus-vektorer som uttrykker vill-type og en dominant-negativ form av Sp35 ble konstruert. DNA-sekvensenkodende mus Sp35 med full lengde(fl-sp35, aminosyrerester av SEQ
135 ID NR:2) ble forsterket med PCR ved bruk av primere 5' -GAG GAT CTC GAC GCG GCC GCA TGG AGA CAG ACA CAC TCC TG-3' (SEQ ID NR:350) og 5' GGG GCG GAA TTG GAT CCT CAC AGA TCC TCT TCT GAG ATG AG-3' (SEQ ID NR:351) og satt inn i HRST-IRESeGFP-lentivirus-vektoren på NotI- og BamHIstedene. På lignende måte, DNA-sekvensendekodende dominant negativt Sp35 (DN-Sp35, aminosyrerester til SEQ ID NR:2) ble forsterket av PCT av primere 5' -GAG GAT CTC GAC GCG GCC GCA TGG AGA CAG ACA CAC TCC TG -3' (SEQ ID NR:352) and 5' -GAT ACG GAT CCT CAG CCT TTG CCC CGG CTC CAT AGA AAC AGC -3' (SEQ ID NR:353). FL-Sp35- og DN-Sp35-plasmidet ble transfektert inn i 293-cellene for å produsere lentivirus som beskrevet i Rubinson et al., «A lentivirus-based system to functionally silence genes in primary mammalian cells, stem cells and transgenic mice by RNA interference,» Nat. Genet. 33: (2003). Oligodendrocyttene ble infisert med lentivirus ved 2 MOI per celle og bekreftet uttrykk til FL-Sp35 og DN-Sp35 med Western blot. [0380] DN-Sp35 fremmet oligodendrocytt-differensiering som produserte en økning i antall modne oligodendrocytter. I motsetning til overuttrykk av Sp35 med full lengde(fl-sp35) som hadde den motsatte effekten og hemmet differensiering som var tydelig på grunn av reduksjonen i antall modne oligodendrocytter i forhold til kontrollen (dataene vises ikke). EKSEMPEL Konstruksjon og rensing av Sp35-Fc-fusjonsprotein [0381] Det ble laget en konstruksjon ved å fusjonere den ekstracellulære delen av humant Sp35 (restene 1 532) til hengslet og Fc-området av human IgG1 for å studere den biologiske funksjonen til Sp35. En delvis kodesekvens for humant Sp35 ble skaffet med PCR fra klone ved bruk av primer i starten 5' -CAG CAG GTC GAC GCG GCC GCA TGC TGG CGG GGG GCG T 3' (SEQ ID NR:354) og primer på slutten 5' -CAG CAG GTC GAC CTC GCC CGG CTG GTT GGC CAA CCA GCC GGG CGA GGT CGA CCT CGA GG -3' (SEQ ID NR:355). [0382] PCR-produktet med butt ende ble subklonet inn i SrfI-stedet på PCR SCRIPT AMP-vektoren (Stratagene) for å danne PCR SCRIPT AMP-Sp35. Et SalI-fragment ble isolert fra PCR SCRIPT AMP-Sp35 og subklonet inn i PCRCAMP Ig-vektoren (avledet fra Stratagene-vektoren PCR SCRIPT AMP). I PCRCAMP Ig-vektoren subklones hengselet og Fc gamma-sekvensen som et SalI(5')- til et NotI(3')- fragment. SalI Sp35-fragmentet ble subklonet inn i SalI-stedet til PCRCAMP Ig-vektoren og dermed fusjonertes Sp35-signalsekvensen og det ekstracellulære domenet (kodonene 1-532) i rammen med sekvenser som koder hengselet og Fc-området til human Ig1. Riktige isolater ble identifisert, og et NotI-fragment som omslutter Sp35 Fc-fragmenter ble subklonet inn i det enkle NotI-klonestedet til CHO-uttrykksvektoren, PV90 (Biogen Idec). Det resulterende plasmidet ble bekreftet med DNAsekvensering og kalt GT123. [0383] Stabile cellelinjer som uttrykker Sp35-Fc-fusjonsproteinet, ble generert med elektroporering av CHO-vertscellene DG44 med plasmid GT123. Transfekterte CHO-celler ble dyrket i alfa minus MEM der det fantes 10 % dialysert serum og 4 mm glutamin til valg ved neukleotiduavhengig vekst. Fjorten dager etter transfeksjon, ble cellene matet ferske medier. CHO-celler med merket med fykoerytrin
136 (PE)-merket anti-humant IgG fra geit (Jackson Labs) og utsatt for flowcytometri-sortering ved høy hastighet i en FACS Mo-Flo (Cytomation) for å screene celler som uttrykte Sp35-Fc. Cellene som uttrykte høye Sp35 Fc-nivåer, ble valgt. Disse cellene ble utvidet i kultur i 7 dager og deretter merket på nytt og sortert på nytt. Cellene som uttrykte høye Sp35 Fc-nivåer, ble isolert som individuelle kloner i 96-brønns plater. Disse klonene ble dyrket i to uker og deretter matet med ferske medier én dag før FACS-analyse for å kontrollere uttrykksnivåene. Cellene som uttrykte høye Sp35 Fc-nivåer ble utvidet og frosne cellebanker ble opprettet. Cellelinjene ble tilpasset for å vokse i suspensjonskultur i det serumfrie mediet BCM16. Titreringen fra Sp35-Fc som ble produsert fra disse klonene, ble bestemt ved å dyrke cellelinjene ved 37 C i 4-5 passeringer og deretter dyrke cellene til 50 % maksimal celletetthet og dyrke dem i dager ved 28 C helt til den variable celletettheten hadde falt til 75 %. På dette tidspunktet ble kulturmediene høstet, cellene og avfall ble fjernet ved sentrifugering og kultur-supernatantene ble titrert for Sp35-Fc-nivåer med en Western blot-analyse med bruk av antihumant Ig-antistoff (Jackson Lab) som proben. [0384] Sp35-Fc-fusjonsproteinet ble renset fra det rensede medium på følgende måte: 9 ml 1M HEPES ph 7,5 ble tilsatt til 900 ml klargjort medium. Mediet ble satt i som batch i 3 timer ved 4 C på 3 ml Protein A Sepharose (Amersharn Bioscience). Harpiksen ble samlet i en 1,5 cm (innvendig diameter) søyle og vasket fire ganger med 3 ml PBS, to ganger med 4 ml PBS som inneholdt 800 mm NaCl og deretter igjen med 3 ml PBS. Sp35-Fc ble eluert fra søylen med 25 mm NaH 2 PO 4, ph 2,8 og 100 mm NaCl i 1,5 ml fraksjoner og nøytralisert ved å tilsette 75 μl 0, 5 M NaH 2 PO 4, ph 8,6. Topp proteininnholdende fraksjoner ble identifisert med absorbans ved 280 nm, samlet sammen og utsatt for videre rensing på en 1 ml Protein A-søyle. Før innlasting ble NaCl tilsatt til 600 mm og HEPES, ph 7,5 til 5,0 mm. Søylen ble vasket to ganger med 600 μl 10 mm HEPES, ph 7,5 and 1 M NaCl og deretter med 1 ml PBS. Sp35-Fc ble eluert fra søylen med 25 mm NaH 2 PO 4, ph 2,8 og 100 mm NaCl, samlet i 0,5 ml fraksjoner og nøytralisert ved å tilsette 25 μl 0,5 M NaH 2 PO 4, ph 8,6. Topp proteininnholdende fraksjoner ble identifisert med absorbans ved 280 nm og samlet sammen. Sp35-Fc-proteinet migrerte som et enkelt bånd (> 95 % rent) med en tilsynelatende masse på 90 kda ved å redusere SDS-PAGE. Under ikke-reduserende forhold, ble proteinet kjørt som en dimer med en omtrentlig masse på 180 kda. Det rensede Sp35-Fc-proteinet ble alikvotert og oppbevart ved -70 C. 30 EKSEMPEL 3 Produksjon av Sp35-Spesifikke monoklonale antistoffer 35 [0385] Anti-Sp35-antistoffer som spesifikt binder et Sp35-polypeptid ble laget ved bruk av følgende metoder og prosedyrer. A. Antistoff-screeninganalyser ELISA-analyse
137 [0386] Sp35-Fc (0,5 μg i 50 μl 0,1 M natriumbikarbonat-buffer, ph 9,0) ble tilsatt til hver brønn med 96-brønns MaxiSorp TM -plater (Nunc TM ). Deretter ble platene inkubert ved 37 C i 1 time eller ved 4 C i 16 timer. Ikke-spesifikke bindingssteder på platene ble blokkert ved bruk av 25 mm HEPES, ph 7,4 som inneholdt 0,1 % BSA, 0,1 % ovalbumin, 0,1 % (5% (v/v) fettfri tørrmelk i 150 mm NACE) og 0,001 % azid. Fortynninger av serum eller hybridom-supernatanter (f.eks. serielle trefolds fortynninger) ble tilsatt over hver platerad og inkubert ved 25 C i 1 time. Etter vasking tre ganger med PBS ble 50 μl 1: fortynning med pepperrot-peroksidase-konjugert anti-mus sekundært antistoff fra geit (Jackson lmmunoresearch Inc.) tilsatt til hver brønn og inkubert i 1 time til. Etter tre vaskingen ble farge utviklet med TMB (punktering) og stoppet med 2 M sulfonsyre. Fargens intensitet ble overvåket i et spektrofotometer ved 450 nm. 2. FACS-analyse [0387] COS-7-celler eller CHO-celler ble merket med 0,1 μm CellTracker Green CMFDA (Molecular Probes, Eugene, OR) som beskrevet av leverandøren. Like volumer av CellTracker -merkede kontrollceller ble blandet med vaskede Sp35-COS-7-celler eller Sp35-CHO-celler (produsert med transient transfeksjon av Sp35-uttrykksvektoren) før inkubering med anti-sp35-testsera eller hybridom-supernatanter. Femti mikroliter av celleblandingen ble dispengert inn i hver brønn ved 96- brønns polystyrenplater med V-bunn (Costar 3877, Coming, NY, USA) og 100 μl museserum, hybridom-supernatant eller kontroll-anti-sp35-antistoff ble tilsatt. Etter inkubering ved 4 C i 30 minutter, ble cellene vasket og inkubert med 50 μl fykoerytrin konjugert affinitetsren F(ab') 2 - fragment med anti-mus IgG Fc gammaspesifikt andre antistoff fra geit (1:200, Jackson ImmunoResearch Laboratory, West Grove, PA, USA) i PBS. På slutten av inkuberingen ble cellene vasket to ganger med PBS og suspendert i 200 μl PBS som inneholdt 1 % fetalt bovinserum (FBS) og FACS-analyseringer. Som et alternativ ble Sp35 COS-7-celler eller Sp35-CHO-celler blandet med museserum eller hybridom-supernatant og deretter behandlet med R-fykoerytrin-konjugert anti-mus sekundært antistoff fra geit og gjennomgikk direkte standard FACS-analyseringer B. Hybridomproduksjon av murine monoklonale anti-sp35-antistoffer [0388] Åtte ukers gamle RBF-hunnmus (Jackson Labs, Bar Harbor, ME, USA) ble immunisert intraperitonealt med emulsjon som inneholdt 50 μg Sp35-Fc (aminosyrene 34 til 532 ved SEQ ID NR:2 fusjonert til hengselet og Fc-området med IgG1), produsert som beskrevet i eksempel 2 eller immunisert intraperitonealt med en emulsjon som inneholder 50 μg human Sp35-Fc, and 50 μl Complete Freund's adjuvant (Sigma Chemical Co., St. Louis, MO, USA) én gang hver andre uke. Sera fra de immuniserte musene ble samlet før den første immuniseringen og 1 uke etter den andre og tredje immuniseringen, og anti-sp35-antistoff-titreringer ble målt av FACS-analysen på Sp35- uttrykkende COS-7-celler som beskrevet ovenfor. En siste booster-dosering ble gitt etter den tredje immuniseringen og tre dager før hybridomfusjonene ble satt i gang. [0389] Sera fra mus immunisert med forskjellige Sp35-peptider, ble screenet med ELISA som beskrevet ovenfor. Mus som var positive for antistoffer som spesifikt bandt Sp35-uttrykkede COS-7-
138 celler, ble identifisert med flowcytometri (FACS) som beskrevet ovenfor og ble avlivet. Splenocytter ble isolert fra musene og fusjonert til FL653-myelom (en APRT-avledning av et Ig-/HGPRT-Balb/cmusemyelom, holdt i DMEM som inneholdt 10 % FBS, 4500 mg/l glukose, 4 mm L-glutamin og 20 mg/ml 8 azaguanin) som beskrevet i Monoclonal Antibodies. Hybridomas: A New Dimension in Biological Analyses, ed. Kennett, R.H., McKearn, T.J. and Bechtol, K.B. New York: Plenum Press (1982). Fusjonerte celler ble lagt på 24- eller 48-brønns plater (Corning Glass Works, Corning, NY, USA) og matet med adenin, aminopterin og tymidin (AAT, tilgjengelig hos Sigma Chemical Co., St. Louis, MO, USA) som inneholdt kulturmedium. AAT-resistante kulturer ble screenet med ELISA eller flowcytometri som beskrevet ovenfor til binding enten til Sp35 COS-7-celler eller Sp35-Fc. Positive hybridomer ble videre subklonet med begrensningsfortynning. [0390] Sytten hybridom-cellelinjer som produserte monoklonale antistoffer produsert fra mus og ble immunisert med Sp35-Fc, ble isolert. Egenskapene til de hybridomavledede monoklonale antistoffene vises i tabell 3A og 3B. [0391] Polynukleotider som koder de variable domenene (V H og V L til de monoklonale antistoffene 1A7, 2F3, 3P1D10.2C3 og 3P1E11.3B7, ble isolert med PCR, klonet og fikk gjennomgå sekvensanalyse med følgende metode. Total RNA ble ekstrahert fra hybridomcellene ved bruk av Qiagen RNeasy minisett, og cdna ble generert fra den isolerte RNA med RT PCR ved bruk av standard betingelser. En «cocktail» med primere ble brukt til RT-PCR. Et foretrukket primersett inkluderte en primer med 5' av primerhybridisering til signalsekvensen og 3'-enden av primerhybridisering til det konstante domenet 3' til FR4/konstantdomene-forbindelsespunktet. Dette muliggjør forsterking av et intakt variabelt domene uten tvetydigheter om det monoklonale antistoff N-terminus og V/C-forbindelsespunktet. En person med ferdigheter i faget vil se at primersettet må modifiseres for å forsterke forskjellige mal og de forskjellige PCR-betingelsene. Av og tid kan tilstedeværelsen av et stort antall ikke-produktive meldinger (f.eks. CDR3-FR4 rammeforskyvet ikke-produktiv lettkjede fra fusjonspartneren) eller ikkespesifikke produktive meldinger produseres og kan komplisere kloningen av variable kjeder. Én løsning er å bruke N-terminalsekvensdata fra autentisk renset antistoff for å utforme en degenerert primer for å aktivere kloning. Som et alternativ kan primere med «universalt rammeverk» brukes, som f.eks. de som beskrives i Orlandi et al, PNAS 86:3833 (1989), som «fastsetter» N- og C-terminus til de variable domenene (dvs. N-terminus til FR1 og C-terminus til FR4 er primerbestemt) [0392] I tillegg kan sekvensdata til utforming av mer effektive primere skaffes fra RT-PCRbulkprodukter som er gelrenset og deretter sekvensert. PCR-produktet kan også subklones ved bruk av f.eks. TOPO Cloning Kit (Invitrogen) og deretter sekvenseres. Deretter skaffes sekvensdata fra flere uavhengige subkloner eller gelrensede fragmenter til fast etablering av konsensussekvens. [0393] Lettkjedesekvensen til P1E11.3B7 ble bestemt ved å bruke en «cocktail» består av 5' murine kappa lettkjede-signalsekvensprimere: (i) 5' GGG GAT ATC CAC CAT GGA TTT TCA GGT GCA GAT TTT CAG 3' (SEQ ID NR:356), (ii) 5' GGG GAT ATC CAC CAT GRA GTC ACA KAC YCA GGT CTT YRT A 3' (SEQ ID NR:357), (iii) 5' GGG GAT ATC CAC CAT GAA GTT GCC TGT TAG GCT GTT G 3' (SEQ ID NR:358), og (iv) 5' GGG GAT ATC CAC CAT GAG GKC CCC WGC TCA GYT YCT KGG A 3' (SEQ ID NR:359), med en enkel 3' murin kappa konstantdomeneprimer: 5' GCG TCT AGA ACT GGA TGG TGG GAG ATG GA 3' (SEQ ID NR:4) der K=G/T, R=A/G, W=A/T og Y=C/T. Det resulterende PCR-produktet ble subklonet og flere
139 uavhengige subkloner ble sekvensert. Den utledede konsensussekvensen var i samsvar med Edmandegraderings-sekvenseringsdata. Sekvenseringen indikerte at degenererings-signalsekvensen 5' primeren 5' GGG GAT ATC CAC CAT GRA GTC ACA KAC YCA GGT CTT YRT A 3' (SEQ ID NR:357) var den somhadde gitt 3P1E11.3B7 variabelt domene med lettkjede under forsterkningen. [0394] 3P1E11.3B7-sekvensen med tungkjede ble bestemt med en «cocktail» består av murine tungkjede-signalsekvens 5' PCR-primere: (i) 5' GGG GAT ATC CAC CAT GGR ATG SAG CTG KGT MAT SCT CTT 3', (SEQ ID NR:360) (ii) 5' GGG GAT ATC CAC CAT GRA CTT CGG GYT GAG CTK GGT TTT 3' (SEQ ID NR:361) og (iii) 5' GGG GAT ATC CAC CAT GGC TGT CTT GGG GCT GCT CIT CT 3' (SEQ ID NR:362) med degenerert murint IgG CH1-konstantdomene 3' primer 5' AGG TCT AGA AYC TCC ACA CAC AGG RRC CAG TGG ATA GAC 3' (SEQ ID NR:363) der K=G/T, M= A/C, R=A/G og Y=C/T. PCR med bruk av denne «cocktail» av primere med en rekke forskjellige kretsprosessforhold, klarte ikke å gi en variabel domenesekvens med tungkjede der det utledede N-terminus var i samsvar med det som ble bestemt av Edman-degraderingssekvensen til det rensede 3P1E11.3B7-antistoffet. Derfor brukte vi universale primere med tungkjede: FRI 5' AGG TSM ARC TGC AGS AGT CWG G 3' (SEQ ID NR:364) og FR4 5' TGA GGA GAC GGT GAC CGT GGT CCC TTG GCC CCA G 3' (SEQ ID NR:365) der M= A/C,R=A/G, S=C/G og W=A/T. Dette settet ga en murint variabelt domene med tungkjede der den utledede sekvensen var i samsvar med de empiriske 3P1E11.3B7-dataene. [0395] For å kunne verifisere at de variable domenene N- og C-terminus med tungkjede var autentiske og ikke primer-bestemt, ble en annen PCR-reaksjon utført med en degenerert signalsekvensprimer 5' ATG GAR TGY AAY TGG ATH CTN CCN TTY A 3' (SEQ ID NR:366) og det ovennevnte konstante domenet 3' primeren 5' AGG TCT AGA AYC TCC ACA CAC AGG RRC CAG TGG ATA GAC 3' (SEQ ID NR:367), der H=A/C/T, N=A/C/G/T og R=A/G og Y=C/T. Utformingen av den degenererte signalsekvensprimeren var basert på signalsekvensene til de beste treffene avledet fra et TFASTA-søk av sekvenser i Genbank-gnagerdatabasen forespurt om 3P1E11.3B7-konsensusutledet FR1 sekvens fra PCR-reaksjonen med «universalprimeren» som beskrives ovenfor. Denne PCR ga et produkt med et komplett murint variabelt domene med tungkjede. [0396] De komplette 3P1E11.3B7 murine variable domenene ble brukt (med stille mutagenese etter behov til innføring i begrensede steder) i forbindelse med human IgG1 og kappa konstant domene cdna-er for å konstruere kimæriske cdna-er med henholdsvis tung- og lettkjede. ImmunoglobulincDNA-ene med full lengde, ble subklonet inn i en uttrykksvektor som kalles pne001, en avledning av den kommersielle EBV-pattedyr-celleepisomal-uttrykksvektoren pcep4. Uttrykksvektorene med tungog lettkjede (kalles henholdsvis pxw372 og pxw363) ble kotransfektert inn i 293-EBNA-cellene. Western blot-analyse (sondert fra humane IgG-spesifikke reagenser) med klargjort medium fra transiente transfekterte celler med bekreftet uttrykk av kimærisk 3P1E11.3B7-huIgG1, kappa mab. De resulterende 3P1E11.3B7 VH- og VL-polypeptidsekvensene vises i tabell 6 og 8 og er henholdsvis SEQ ID NR: 173 og 209. Sekvensene med tung- og lettkjede til 1A7, 2F3 og 3P1D10.2C3 monoklonale antistoffer med bestemt med lignende metoder. 40 C. Identifisering av anti-sp35 monoklonale antistoffer med fagedisplay
140 [0397] Anti-Sp35 monoklonale antistoff-fab-fragmenter ble identifisert og isolert fra fagedisplaybiblioteker som beskrevet i Hoet et al., Nat. Biotech. 23: (2005), Rauchenberger, et al., J. Biol. Chem. 278: (2003) og Knappik, et al., J. Mol. Biol. 296:57-86 (2000). [0398] MorphoSys Fab-fagedisplay-biblioteket HuCAL GOLD («Fagedisplay-bibliotek-2» i tabell 3B) som omfatter humaniserte syntetiske antistoff variable områder, ble screenet mot rekombinant humant oppløselig Sp35-Fc-protein med standard ELISA OG IHC-screeningmetoder. Se f.eks. Ostendorp, R., Frisch, C. and Urban M, «Generation, engineering and production of human antibodies using HuCAL.» Antibodies, Volume 2 Novel Technologies and Therapeutic Use. New York: Kluwer Academic/Plenum (2004). Fab-fager som spesifikt bindes til Sp35, ble renset og karakterisert. Egenskapene til disse fagedisplayavledede monoklonale antistoff-fab-fragmentene vises i tabell 3B som «fagedisplay-bibliotek-2-avledede monoklonale Fab-fragmenter.» Isolert Fab-fage 1968 ble valgt for videre analyse. 15 EKSEMPEL 4 Immunopresipitering av Sp35 av anti-sp35 monoklonale antistoffer [0399] For å utføre immunopresipiteringen ble COS-1-celler som uttrykte Sp35 fusjonert til en hemaglutinin (HA)-tagg på N-terminus, produsert med transient transfektering av COS-1-celler med en DNA-konstruksjon som uttrykker Sp35-proteinet med full lengde med en HA-tagg. Cellene ble høstet 48 timer etter transfeksjonen og ble lysert i 1 ml lysisbuffer (50 mm HEPES, ph 7,5, 150 mm NaCl, 1,5 mm MgCl 2, 1mM EGTA, 1 % Triton X-100 og 10 % glyserol) i 30 minutter ved 4 C. Etter sentrifugering ved xg i 15 minutter ble supernatantene inkubert med Protein A/G-Sepharose-kuler (Santa Cruz) ved 4 C i 1 time og deretter inkubert ved 4 C i 1 time enten med 1A7- eller 2F3-anti-Sp35 murine monoklonale antistoffer. Kulene ble vasket 3 ganger med lysisbuffer, kokt i Laemmliprøvebuffer, utsatt for 4-20 % SDS PAGE og analysert med Western blot ved bruk av et antistoff som gjenkjenner HA-tagging. Som vist på SDS-PAGE-gel, monoklonale antistoffer 1A7 og 2F3, immunopresipitert humant og murint Sp35 (fig. 1). Som vist i fig. 1, immunopresipiterte monoklonalt antistoff 2F3 sterkt immunopresipiterte både humant og murint Sp35 mens monoklonalt antistoff 1A7 som sterkt immunopresipiterte humant Sp35, kun gjenkjente murint Sp35-protein svakt. På lignende måte immunopresipiterte monoklonale antistoffene 1G7, 2B10, 2F3, 3P4C2.2D2, 3P4C8.2G9, Li01, Li03, Li05, Li06, Li07, Li08, Li11, Li12, 7PID5.1G9 og 3B5.2 humant eller muse- eller humant og muse- Sp35 (se tabell 3B og 3C). I tillegg immunopresipiterte Li08 AP Sp35 og monoklonale antistoffer 1B6.4 og 3E3.1 immunopresipiterte endogent Sp35 (se tabell 3B). EKSEMPEL 5 Anti-Sp35-antistoff-binding spesifikt til Sp35 bestemt med ELISA 40 [0400] For å bestemme hvilke områder av Sp35-polypeptidet som ble bundet til de forskjellige hybridom- og fagedisplayavledede monoklonale antistoffer produsert i eksempel 2, ble en ELISA-
141 analyse utført ved bruk av et panel med avkortede Sp35-polypeptider der hvert ble fusjonert til hengselet og Fc-områdene til IgG1 med metodene som beskrives i eksempel 1. Panelet bestod av følgende Sp35-fragmenter: aminosyrene ved SEQ ID NR:2, aminosyrene ved SEQ ID NR:2, aminosyrene ved SEQ ID NR:2 og aminosyrene ved SEQ ID NR:2. Ovalbumin og BSA ble brukt som kontroller. Som vist i tabell 3B tyder de hybridomavledede mab-ene 2F3, 2B10, 3A3, 3P4c2.2d2 og 3P4c8.2g9 og de Fab-fageavledede mab-ene 3383, 3563, 3564, 3565, 3568, 3569, 3570 og 3582 som alle er spesifikt bundet til og Sp35-fragmentene, på at disse antistoffene bindes til epitoper i LRR-området til Sp35. Hybridomavledede Mab-ene 1A7, 3P1B11F9, 3P1D10.2C3, 3P1E11.3B7, 3P2C63G10.2H7, 2P2C9.2G4, 3P4A61D9 og 394C51D8 og Fab-fageavledede Mab-ene 3495,3566,3567 og 1968 bindes spesifikt til Sp35-fragmentet og bindes svakt til Sp35 som tyder på at disse antistoffene sannsynligvis bindes til epitoper som minst inkluderer en del av Sp35 C-terminalen til LRR-området. I lignende forsøk bandt disse sistnevnte antistoffene også spesifikt til et Sp35-polypeptid som bestod av aminosyrene med humant Sp35 og med lav affinitet til mus- og rotte-sp35. Affiniteten til disse sistnevnte antistoffene for mus- og rotte-sp35, ble gjenopprettet til nivået som sees, ved bruk av humant Sp35 når aminosyren 419 til mus- eller rotte-sp35 endres fra histidin (H) til arginin (R). Arginin er aminosyren i posisjon 419 i humant Sp35. K D for monoklonalt antistoff 1A7 ble bestemt til 10 nm (1 x 10-9 M) for binding av humant Sp35 og 20 μm (2 x 10-5 M) til binding av murint Sp35. For at Ap-Sp35 ELISA skal påvise antistoffene som bindes til 417 til 532 området, ble ELISA utført på følgende måte: Mab-ene ble belagt på ELISA-platene og deretter inkubert enten med et Sp35-AP-fusjonsprotein ved 4 C over natten med påfølgende APkoplet anti-human (H+L) (1:5000, Jackson ImmunoResearch) ved romtemperatur i 1 time eller med AP-fusjonsproteiner ved 4 C over natten. AP-substrat ble deretter utviklet med 10 mg/ml 4NPP i 0,1 M glysin, 1 mm MgCl 2, 1 mm ZnCl 2, ph 10,5 og lest ved utvendig diameter 405. [0401] I lignende forsøk ble de monoklonale antistoffene 3B5.2 og 7P1D5.1G9 samt Fab-fragmentet til 7P1D5.1G9-antistoffet testet i ELISA-analyser for å finne deres evne til å binde humant Sp35, hele LRR-området til Sp35, Ig-området til Sp35 og murint Sp35. Som vist i tabell 3C, binder 3BS.2, 7P1DS.lG9 og 7P1DS.1G9 Fab-fragmentet humant og murint Sp3S. 3BS.2 og 7P1D5.1G9 monoklonale antistoffer binder også LRR-området til Sp35. Se tabell 3C. [0402] I en Sp35-bindingsanalyse med 3B5.5 som var monoklonalt fiksert til bunnen av en vevskulturbrønn, blokkerte ikke følgende antistoffer 3BS.2-binding av Sp35: Li03, Li05, Li08, Li011 og Fab-fragment av Li013. EKSEMPEL 6 35 Anti-Sp35-antistoff-binding spesifikt til Sp35 bestemt med FACS 40 [0403] For å karakterisere bindingsegenskapene til hybridomavledede anti-sp35-mab-ene 1 A7 og 2F3 som produseres som beskrevet i eksempel 3, ble binding til både fiksert og levende COS-7- eller 293- celler som uttrykker muse- eller humant Sp35 sammenlignet. Sp35-transfekterte og ikke-tranfekterte celler ble fiksert og gjennomgikk en FACS-analyse (FACS: Celler transfektert med humant og mus Sp35
142 eller vektorkontroll be dissosiert fra kulturplater, vasket med 2 %FBS/PBS og inkubert med primært antistoff ved 1 μg/ml på is i 1 time. Cellene ble vasket 3 ganger med 2 % FBS/PBS og deretter inkubert med PE-merket sekundært antistoff (1: 100, JacksonImmunoResearch) på is i 30 minutter. Etter 2 vasker med 2 % FBS/PBS ble cellene fiksert i 2 % PFA og gjennomgikk en FACS-analyse med PE.) FACSresultatet viser at MAbs 1A7 og 2F2 bindes til COS-7 eller 293-cellene som uttrykker Sp35, men ikke bindes til kontrollcellene med ingen Sp35-uttrykk (fig. 2). [0404] I lignende forsøk ble CHO-cellene som ble stabilt transfektert med Sp35, brukt i en FACSanalyse til videre karakterisering av bindingsegenskapene til 3B5.2 og 7P1D5.1 G9. Som vist i FIG. 11, 3B5.2 og 7P1D5.1G9 bandt til Sp35 på transfekterte CHO-celler. Mer spesifikt, 3B5.2- og 7P1D5.1G9- antistoffene ble testet når det gjaldt deres evne til å binde CHO-celler som transfekteres med og uttrykker human eller murint Sp35. Som vist i FIG. 11 ble antall celler som bindes til 3B5.2- og 7P1D5.1 G9-antistoffene og målt med MCF (gjennomsnittlig fluorescens), økt med antistoffkonsentrasjonen som ble brukt. I tillegg bandt 3B5.2-antistoffet flere celler som målt med MCF enn 7P1D5.1G9. EKSEMPEL 7 Nevrittutvekst-analyse [0405] For å teste evnen til de hybridomavledede eller Fab-fageavledede monoklonale antistoffene som produseres ovenfor for å reversere hemmingseffekten til CNS-myelinhemmerne, f.eks. OMgp, på nevroner, ble Lab-Tek -kultur-objektglass (4 brønner) belagt med 0,1 mg/ml poly-d lysin (Sigma ). Ap-OMgp (1 μg per «spot») eller PBS med «spottet» med 3 μl-dråper. Lab-Tek -objektglass ble deretter skylt og belagt med 10 μg/ml laminin (Gibco ). Dorsale rotganglioner (DRG-er) fra P6-7 Sprague Dawley-rotteunger ble dissosierte med 1 mg/ml kollagenase-type 1 (Worthington), triturert med brannpusset Pasteur-pipetter som er forhåndsovertrukket for å berikes i nevronale celler og til slutt overtrukket med celler/brønn på de forhåndsbelagte Lab-Tek -kultur-objektglassene. Ti μg/ml mab 1A7 eller 2F3 ble tilsatt straks etter overtrekking av DRG-ene. Kulturmediet var F12 (tilgjengelig hos Gibco/Invitrogen) som inneholdt 5 % varminaktivert donor-hesteserum, 5 % varmeinaktivert fetalt bovinserum og 50 ng/ml musenerve-vektsfaktor (mngf) og inkubert ved 37 C og 5 % CO 2 i 6 timer. Etter inkubering ble objektglassene fiksert i 4 % paraformaldehyd / 20 % sukrose og farget med anti-βiii-tubulin TUJ1-antistoff (Covance) etter 16 timer. [0406] Som sekundært antistoff, ble anti-mus Alexa-Fluor 594 (Molecular Probes) som ble fortynnet 1:300, tilsatt til objektglassene og inkubert i 2 timer ved romtemperatur. Objektglassene ble dekket med GelMount (BiØemeda ). 5 x digitale bilder ble skaffet med OpenLab -programvare (Improvision, Inc., Lexington, MA, USA), og bildene ble analysert for kvantifisering av nevritt-utvekst ved bruk av OPENLAB TM programvaren, alt iht. produsentens spesifikke parametere. [0407] Både MAb 1A7 og 2F3 beskyttet DRG-nevronene fra OMgp-mediert hemming av nevritt utvekst. (Fig 3). 3B5.2 beskyttet også DRG-nevronene fra OMgp-mediert hemming av nevritt-utvekst (dataene vises ikke).
143 142 EKSEMPEL Monoklonalt antistoff 1A7 fremmer funksjonell gjenvinning i ryggradsskademodellen med rotter [0408] Ryggradsskade (SCI) ble indusert med dorsal over-hemi-seksjon på følgende måte, modifisert fra metoder som er beskrevet tidligere (Li, S. et al. J. Neurosci. 24, (2004)). Bedøvede Long Evans-hunnrotter (7 uker gamle, Charles River) ble gitt preoperativ analgesi (Buprenorphine/Buprenex, 0,05 mg/kg s.c.) og beroligende middel (Midazolam, 2,5 mg/kg i.p.), og en dorsal hemiseksjon ble utført på brystvirvel 6/7 som fullstendig forstyrrer hoved-dorsomedial og - dorsolateral pyramidebanen (CST). De dorsale og dorso-laterale komponentene til pyramidebanen ble fullstendig forstyrret og den ventrale delen av CST ble værende intakt. Den ventrale vevsbruen ble værende igjen etter at hemiseksjonen utgjorde omtrent 20 % av barken i begge behandlingsgruppene (dataene vises ikke). [0409] Bakre lemfunksjon ble kvantifisert ved bruk av Basso-Beattie-Bresnahan (BBB) åpentfeltskåringmetode (Eby, M.T. et al., J. Bioi. Chem. 275, (2000), og alle dyrene fikk vedvarende og merkbare funksjonelle underskudd etter SCI med nesten fullstendig lammelse i det bakre lemmet dagen etter inngrepet. Straks etter CST-transeksjon, ble et intratekalt kateter satt inn i det subaraknoide rommet ved T7 og så koblet til en primet mini-osmotisk pumpe (Alzet-modell 2004, Alza Corp) som ble satt inn i det subkutane rommet. Mini-osmotiske pumper tilførte humant IgG isotype kontrollprotein (5 mg/ml) eller monoklonalt antistoff 1A7 (4,8 mg/ml) kontinuerlig med en hastighet på 0,25 μl/time i 5 uker. Kontroll (human IgG-behandlede)-dyrene gjenvant vesentlig funksjon i løpet av de 5 ukene som forsøket pågikk, men flatet ut etter 3-4 uker. Til slutt vare gjennomsnittlig BBB-skåre 9 ± 0,45 (fig. 7). I motsetning til dette, førte kontinuerlig intratekal infusjon av 1A7 i 5 uker til signifikant bedring av BBB-skårene hos kontrolldyr i løpet av 5 uker med fortsatt forbedret funksjon i 2-5 ukes tidsrammen og med oppnådd gjennomsnittlig BBB-skåre på 11,1 ± 0,7 (fig. 4). Disse resultatene viste at behandling med anti-sp35 monoklonalt antistoff 1A7 fremmet gjenoppretting av funksjonen et en ryggradsskade som demonstrert av en økning i BBB-skåren, aksonregenerering og mindre aksonretraksjon som ble observert med immunohistokjemisk farging av aksonene. Antistoff 3B5.2 fremmet også gjenoppretting etter en ryggradsskade (dataene vises ikke). EKSEMPEL 9 35 Anti-Sp3S-antistoffene 1A7, 2F3, 3P1D10.2C3, 3P1E11.3B7, 6P4F4.1D3, 6P4F4.1F9, 7P1DS.1G9, Li05, Li06, Li08, Li13, Li28, Li33, D05, D08 og 3B5.2 fremmer in vitro [0410] Rollen til anti-sp35-antistoffene 1A7 og 2F3 i myelinering ble undersøkt in vitro ved å behandle ko-kulturene DRG (dorsalrotganglion)-nevronene og oligodendrocyttene med anti-sp35- antistoffene 1A7 og 2F3 og teste for myelinering med immunohistokjemi og Western blot. Ved disse
144 studiene er det nødvendig å først generere primære kulturer av DRG-nevronene og oligodendrocyttene. [0411] Long Evans-hunnrottene E14-E17 embryonisk disalrotganglion ble dyrket som beskrevet i Plant et al., J. Neurosci. 22: (2002). Dissekerte DRG-er ble overtrukket poly-l-lysin-belagte dekkglass (l00 μg/ml) i to uker. Cellene ble inkubert i tilstedeværelse av fluordeoksyuridin fra dag 2-6 og i NLA-medium som inneholdt 1 x B27, 100 ng/ml NGF (Gibco) fra dag [0412] Long Evans-hunnrotter postnatal dag 2 (P2)-oligodendrocytter ble dyrket som beskrevet i Conn, Meth. Neurosci. 2: 1-4 (Academic Press; 1990) med følgende modifikasjoner. I korthet, forhjernen ble fjernet fra P2-rotter og plassert i kaldt HBSS-medium (Gibco). Vevsfragmentene ble kuttet inn i 1 mm stykker og inkubert ved 37 C i 15 minutter i 0,01 % trypsin og 10 μg/ml DNase. Dissosierte celler ble lagt på poly-l-lysin belagt T75-vevskulturflasker og dyrket i DMEM med 20 % fetalt bovinserum ved 37 C i 10 dager. A2B5-positive oligodendrocytter ble samlet ved å riste flaskene over natten ved 200 omdreininger per minutt ved 37 C. A2B5-oligodendrocyttene ble dyrket i 7 dager i DMEM (Gibco) som inneholder 25 mm D-glukose, 4 mm L-glutamin, 1 mm natriumpyruvat, 50 μg/ml humant apo-transferrin, 5 μg/ml bovint pancreasinsulin, 30 nm natriumselenat, 10 nm hydrokortison, 10 nm D-biotin, 1 mg/ml BSA, 10 ng/ml FGF og PDGF (Peprotech). Cellene ble deretter høstet med trypsinisering. Deretter ble cellene dyrket sammen med DRG-nevroner der det fantes eller ikke fantes 1,3, 10 eller 30 μg/ml anti-sp35 monoklonale antistoffer 1A7 eller 2F3 eller et negativt kontrollantistoff i et NLA-medium som inneholdt 2 % fetalt bovinserum, 50 μg/ml askorbinsyre, 100 ng/ml NGF (Gibco). Det ble bestemt at en effektiv antistoff-dose å administrere i en slik analyse, er mellom 0,1 μg/ml og 10 μg/ml, avhengig av antistoffet. En person med ferdigheter i faget vil kunne bestemme en effektiv dose ved bruk av analysene som beskrives i dette dokumentet. [0413] Kulturmediet ble endret og de forskjellige monoklonale antistoffene ble etterfylt hver 3. dag. Etter 30 dager ved 37 C ble cellene som ble dyrket sammen, farget med immunohistokjemisk farging (IHC) for nevrofilamentene med anti-βiii-tubulin antistoff for å identifisere aksoner eller anti-mbpantistoff for å identifisere oligodendrocytter (fig. 4A-E). Cellene som ble dyrket sammen, ble også lysert og gjennomgikk en Western blot-analyse for å kvantifisere MBP (fig. 4G). Basert på IHC- og Western blot-analysene utviste cellene som ble dyrket sammen og som ble behandlet med anti-sp35- antistoffene 1A7 and 2F3, økt overlevelse av oligodendrocytter og nevroner, økt antall sammenbuntede aksoner og økt antall MBP-positive celler (fig. 4F, 10-fold flere MBP positive celler i forhold til ko-kulturene som ble behandlet med kontroll-antistoff. [0414] I et lignende forsøk ble oligodendrocyttene og DRG-ko-kulturene inkubert der det fantes eller ikke fantes anti-sp35-antistoffene Li05 og Li06 eller et negativt kontroll-antistoff. Cellene som ble dyrket sammen, ble lysert og gjennomgikk en Western blot-analyse for å kvantifisere MBP (fig. 8). Basert på Western blot-analysene utviste cellene som ble dyrket sammen og som ble behandlet med anti-sp35-antistoffene Li05 og Li06, økt antall MBP-positive celler som lignet på cellene som ble dyrket sammen og som ble behandlet med 3, 10 og 30 μg Sp35-Fe (LINGO-1-Fc). [0414] I et lignende forsøk ble oligodendrocyttene og DRG-ko-kulturene inkubert der det fantes eller ikke fantes anti-sp35-antistoffene 3B5.2, 3P1D10.2C3, 3P1E11.3B7, 6P4F4.1D3, 6P4F4.1F9, 7P1D5.1G9, Li08, Li13, Li28 og Li33 og som også fremmet myelinering. På lignende måte fremmer
145 antistoffene DO5 og D08 med full lengde, myelinering. Den laveste effektive dosen med 3B5.2- og 7P1D5.1G9-antistoff som trengs for å fremme myelinering i DRG-ko-kulturforsøk var 0,1 μg/ml. [0416] I tillegg ble 7PID5.1G9 Fab-fragmentet i en lignende in vitro myelineringsanalyse. 7P1D5.1 G9 Fab-fragmentet fremmet myelinering ved en konsentrasjon på 1,0 μg/ml. [0417] Disse resultatene indikerer at behandling av DRG-oligodendrocytt-ko-kulturer med anti Sp35- antistoffene 1A7, 2F3, 3P1D10.2C3, 3P1E11.3B7, 6P4F4.1D3, 6P4F4.1F9, 7P1D5.1G9, Li05, Li06, Li08, Li13, Li28, Li33, D05, D08 og 3B5.2 fremme modne oligodendrocytt-akson-interaksjoner og myelinering i forhold til kontroll-antistoffbehandlede ko-kulturer. EKSEMPEL 10 Anti-Sp35-antistoffer og Fab-fragmenter fremmer oligodendrocytt-overlevelse og myelinering in vivo [0418] Voksen vill type C57B1/6-hannmus ble matet med cuprizon (0,2 % valset med malt musemat etter vekt) i 6 uker for å indusere demyelinering inni corpus callosum iht. metoden som beskrives i Morell P et al., Mol Cell Neurosci. 12:220-7 (1998). I korthet, anti-sp35 monoklonalt antistoff-1a7 ble stereotaktisk injisert inn i demyelinerings-corpus callosum i uke 2, 2,5 og 3 med cuprizonmating med metoden som beskrives nedenfor. Kontrollmus ble stereotaktisk injisert med samme intervallene med steriliserte medier som inneholder kontroll-antistoff. Etter 6 uker med cuprizonmating var ferdig, ble musene returnert til normal diett i 2, 4 og 6 uker (kun malt musemat) for å muliggjøre remyelinering. [0419] 1A7 og de monoklonale kontroll-antistoffene ble tilført på følgende måte. De cuprizonbehandlede musene ble bedøvet med ketamin (80 mg/kg kroppsvekt) and xylazin (10 mg/kg kroppsvekt) og plassert i et immobiliseringsapparat som var utformet til stereotaktisk kirurgi (David Kopf Instruments). Skallen ble åpnet og sterile sammensetninger ble injisert (1 μm i 1 ml HBSS) ensidig inn i den svært demyelinerte corpus callosum i vill type mottakermus med en 10 μl Hamilton-sprøyte ved bruk av de stereotaktiske koordinatene 0,7 mm posteriørt og 0,3 mm lateralt til bregma på en dybde på 1,7 mm (Messier et al., Pharmacol. Biochem. Behav. 63: (1999)). Flere kontroll-mottakermus ble stereotaktisk injisert med HBSS som ikke inneholdt sammensetninger. Åpningen i skallen ble fylt med Gelfoam og området ble penslet med penicillin og streptomycin (Gibco) og såret ble lukket med sutur. Mus ble avlivet hver uke som forsøket pågikk etter injisering og deres hjerner ble fjernet og behandlet for molekylær, biokjemisk og histologisk analyse. [0420] Dyrene som fikk anti-sp35-antistoff-1a7-behandling, utviste økt moden oligodendrocyttoverlevelse (basert på CC1-antistoff-farging, fig. 5A) og aksonmyelinering med IHC ved bruk av anti- MBP-proteinantistoff eller Luxol Fast Blue (fig. 5B). CC1-antistoff-positive oligodendrocytter ble kvantitert ved fire uker og 6 uker (fig. 5C). Disse resultatene indikerte at anti-sp35-antistoff-1a7- behandling fremmet moden oligodendrocytt-overlevelse og aksonmyelinering i forhold til mus behandlet med kontroll-antistoff. På lignende vis, mottok dyr 1A7-antistoffet eller 1 μg/ml 7P1D5.1G9
146 145 Fab-fragment i en lysolecitin-modell med demyelinering som også fremmet aksonmyelinering i forhold til kontrolldyrene. 5 EKSEMPEL 11 Anti-Sp35-antistoff-1A7 fremmer retinal ganglioncelle (RGC)-overlevelse i synsnervetranseksjonsmodell [0421] Anti-Sp35-antistoff-1A7 ble testet i en synsnerve-transeksjonsmodell som undersøkte faktorer som påvirker den nevronale funksjonen. Unge voksne Sprague Dawley (SD)-hunnrotter ble brukt i denne studien. Den høyre synsnerven til hvert dyr ble transektert intraorbitalt 1,5 mm fra synsnervepapillen. Et stykke gelskum som ble bløtlagt med 6 % Fluoro-Gold (FG), ble påført stedet som nylig var transektert rett bak synsnervepapillen for å merke de overlevende retinale ganglioncellene (RGC-ene). Dyrene ble delt inn i tre grupper (n=6 i hver gruppe) som fikk enten anti- Sp35-antistoff-1A7, kontroll-antistoff eller kun PBS, med intravitreal injeksjon. Volumet til hver intravitreal injeksjon var 4 μl mens dosen i hver injeksjon var 2 μg. De intravitreale injeksjonene ble utført rett etter synsnerve-transeksjonen. [0422] Alle dyrene fikk overleve i 1 uke. To dager før avlivning av dyrene, ble venstre synsnerven hos hvert dyr transektert og 6 % FG ble administrert som beskrevet ovenfor for å merke de overlevende RGC-ene, for å tjene som intern kontroll. Dyrene ble avlivet med en overdose av Nembutal og retinaene ble dissekert i 4 % paraformaldehyd. Fire radiale kutt ble tatt for å dele retinaene inn i fire kvadranter (superior, inferior, nasal og temporal). Deretter ble retinaene postfiksert i samme fikservæske i 1 time før de ble flatmontert med monteringsmedium (Dako). Objektglassene ble undersøkt med et fluorescensmikroskop ved bruk av et ultrafiolett filter (eksitasjons-bølgelengde = nm). Merkede RGC-er ble telt langs medianlinjen for hver kvadrant der man startet med synsnervepapillen til den perifere grensen til retina med 500 μm intervaller, med et okulært nett 200 X 200 μm 2. Prosentandelen av overlevende RGC-er som var et resultat av hver behandling, ble uttrykt ved å sammenligne antall overlevende RGC-er i de skadede øynene i forhold til de kontralaterale øynene. Alle data ble uttrykt som gjennomsnittlig ± SEM. Statistisk signifikans ble evaluert med one way ANOVA med påfølgende Tukey-Kramer post hoc test. Differansene ble regnet som signifikante for p < 0,05. Anti-Sp35-antistoff-1A7-behandlede dyr utviste større nevronal overlevelse (80 %) i forhold til kontroll-antistoff eller PBS-behandlede dyr, der hver av disse utviste omtrent 50 % nevronal overlevelse (fig 6). 35 EKSEMPEL 12 Testing av anti-sp35-antistoffer for remyelinering i synsnerve-knusingsmodellen
147 [0423] Høyre synsnerve knuses helt med en nr. 5 tang i 10 sekunder omtrent 1,5 mm bak øyeeplet intraorbitalt rett før administrasjon av 2 μl monoklonalt antistoff 1A7, 2F3, Li05 og Li06 i 2ml med intravitreal injeksjon. [0424] Dyrene fik en andre intravitreal injeksjon med samme behandling én uke etter inngrepet. To uker etter inngrepet ble dyrene perfusert med EM-fikseringsvæsker, post-fiksert og behandlet for semitynne og ultratynne seksjoner. De langsgående synsnerveseksjonene ble farget og preparert for myelinobservasjon. Myelinering av de proksimale og distale delene av den knuste synsnerven ble sammenlignet blant forskjellige behandlingsgrupper. Sp35-Fc- og 1A7-, 2F3-, Li05- og Li06-behandlede dyr samt aktuelle kontroller, blir analysert for remyelinering i den distale delen av synsnerven i forhold til kontrollene. EKSEMPEL Testing av anti-sp35-antistoffer for regenerering i synsnerve-knusingsmodellen [0425] Høyre synsnerve ble knust med en nr. 5 tang i 10 sekunder omtrent 1,5-2 mm bak øyeeplet intraorbitalt rett før administrasjon av 2 μg monoklonalt antistoff 1A7 i PBS med intravitreal injeksjon. Fire rotter ble testet med 1A7-antistoff, og 8 røtter ble brukt som kontrolldyr. Dyrene fik en andre intravitreal injeksjon med samme behandling én uke etter inngrepet. Tre dager før avlivning av testdyrene (dag 11 av forsøket), ble 2 ml CTB/FITC injisert intravitrealt for å merke anterograd, regenerative synsnerveaksoner. På den 14. dagen etter inngrepet, ble dyrene perfusert og postfiksert. Den knuste synsnerven ble behandlet for frosne langsgående seksjoner. CTB FITC-merkede aksoner som krysset lesjonsstedet, ble telt som regenerative fibre ved forskjellige avstander bak det knuste stedet. Da 1A7 ble injisert i øyet, ble regenereringsaksoner observert inntil 250 μl bak det knuste stedet. Se fig. 10. EKSEMPEL Anti-Sp35-antistoff fremmer remyelinering og reparasjon av synsnerven ved bruk av MOG-indusert EAE-rottemodell [0426] Ved disse forsøkene ble induserte MOG (myelin-oligodendrocytt-glykoprotein) EAE (Experimental Autoimmune Encephalomyelitis)-rottemodell brukt. Dette er dyremodellen for human multippel sklerose. 50 μl 200 ng Complete Freund's adjuvant (Chondrex Inc.) samt 50 μl av 50 μg MOG i saltvannsløsning ble emulsifisert (1:1) og holdt på is før den ble injisert intradermalt ved halebunnen på hvert dyr. Brune Norway-hunnrotter, 8-10 uker gamle ble brukt ved alle forsøkene. Generelle observasjon innen faget indikerer at EAE-modellen induseres rundt 15 dager etter MOG-injeksjon. Rottene skåres for kliniske tegn på EAE. Tegnene skåres på følgende måte: gradering 0,5, distal parese av halen, gradering 1, fullstendig parese av halen, gradering 1,5, parese av halen og mild parese av bakre bein, gradering 2,0, unilateral, alvorlig parese av bakre bein, gradering 2,5, bilateral,
148 alvorlig parese av bakre lem, gradering 3,0, fullstendig bilateral parese av bakre lem, gradering 3,5, fullstendig bilateral parese av bakre lem og parese av et fremre lem, gradering fullstendig parese (tetraplegia), dødende eller død. Dyrene mottar behandling én gang etter at EAE-modellen induseres. [0427] 2μg/μl av et anti-sp35-antistoff (1A7) ble injisert intravitrealt på dag 15 ved MOG-EAEinduksjon. 2μg/μl av anti-sp35-antistoffet 1A7 ble injisert to ganger til på dag 22 og dag 28. På slutten av forsøket ble dyrene perfusert med 4 % PFA. Synsnervene ble postfiksert i 1 % OsO 4, dehydrert og lagt inn i Epon. Semitynne seksjoner (1μM) ble kuttet og farget med Toluidine Blue til evaluering av myelinering. Synsnervene til behandlede dyr ble sammenlignet med ubehandlede dyr for aksonregenerering og remyelinering i synsnerven. Alle prosedyrene ble utført iht. en protokoll som er godkjent av IACUC (Institutional Animal Care and Use Committee). [0428] Dyrene som fikk behandling med anti-sp35-antistoff-1a7, utviste remyelinering av reparasjon av synsnerven i forhold til normale synsnerver eller dyr som gjennomgikk MOG-indusert EAE, men som ikke fikk behandling (fig. 9). I fig. 9C peker pilene på myelinerte aksoner. Dyrene som fikk et antistoff som gjenkjente domene III av Protein G fra Streptococcus (MOPC21), ikke spesifikt fra Sp35, viste ingen tegn til remylinering eller reparasjon av synsnerven i forhold til normale synsnerver til ubehandlede dyr (dataene vises ikke). Sp35 antagonist antistoff 1A7 fremmet remyelinering og reparasjon av synsnerver en en rotte-mog-indusert EAE-synsnevritt-modell (fig. 9). 20 EKSEMPEL 15 Testing av anti-sp35-antistoffer for fremming av CNS-remyelinering ved bruk av MOG-indusert EAEmusemodell [0429] EAE ble indusert i 129B6 blandet musestamme med intradermal immunisering (dag 0) med 100 μg MOG1-125 protein som ble emulsifisert med Complete Freund's adjuvant (CFA). Det injiserte volumet var 100 μl per mus og var fordelt over 3 steder (pinna, rygg og hud). Emulsjonen ble preparert basert på et volumforhold 1:1 og innholdet 1 mg/ml MOG1-125 og 2 mg/ml M. tuberculosis (stamme H37Ra, Chondrex) Pertussis toxin (200 ng/mus) administreres intraperitonealt på immuniseringstidspunktet og 2 dager etterpå. Kroppsvekt og kliniske EAE-skårer (0 = ingen kliniske tegn, 1 = slapp hale, 2 =svakhet i bakre lem, svekket refleks for å få dyret til å komme tilbake i oppreist stilling eller vraltende gange, 3 = fullstendig parese av bakre lem eller manglende refleks for å få dyret til å komme tilbake i oppreist stilling, 4 = fullstendig parese av bakre lem der forlemmet er involvert til en viss grad, 5 = dyret er fullstendig paralysert, 6 = dødende eller død) ble registrert hver dag. Alle prosedyrene ble utført iht. en protokoll som er godkjent av IACUC (Institutional Animal Care and Use Committee). Dyrene fikk behandling med 1A7, 2F3, Li05 og Li06 monoklonale antistoffer eller kontroll antistoff på dag 0 av studien. Blodprøver ble tatt på forskjellige tidspunkter i løpet av forsøkene med retroorbital-blødemetode. Plasma ble skilt ut fra PBMC med sentrifugering og cellefenotyping som ble utført med FACS-farging. Profilering av den humorale anti-mog-antistoffresponsen ble utført med ELISA ved bruk av underklasse-/isotypespesifikke mab-er (Pharmingen). På slutten av hvert forsøk ble hjernen, ryggraden, synsnerve og isjiasnerven høstet etter perfusjon.
149 148 5 [0430] Denne samme protokollen ble brukt til å indusere EAE-en i Sp35-«knockout»-mus og mus av samme kull. Sp35-«knockout»-mus viser vanligvis lavere EAE-skår (1,5), og ingen relaps i forhold til kontrollen (i en 45-dagers periode)og deretter vill type mus fra samme kull (EAE-skår 3,5). [0431] Sp35-Fc- og 1A7-, 2F3-behandlede dyr blir analysert for remyelinering i forhold til kontrollen. [0432] His-tagget MOG protein ble uttrykt i Pichia pastoris ved bruk av doksysyklin-induserbar TetO-AOX1-promoter (M. Levesque, D. Krushinskie and K. Strauch, manuscript in preparation). Den ekstracellulære kodesekvensen (Gly1 til Gly125 til det modne proteinet etter at signalsekvensen ble fjernet) til rotte-mog ble PCR-forsterket ved bruk av følgende primere: 10 EKSEMPEL 16 Konstruksjon av en 3B5.2-variant [0433] Følgende er aminosyresekvensen til det variable lette kjedet (VL) til 3B5.2-antistoffet, CDR-er er understreket og N-koplede glykosyleringsted er uthevet og med dobbel understreking: QIVLTQSPAl MSASPGEKVT MTCSASSRVS YVHWYQQKSG TSPKRWLYDT SNLASGVPAR FGGNGSGTSY SLTISSMEAE DAATYYCQQW STNPPTFGGG TKLEIK (SEQ ID NR:417). For å bestemme om uttrykk av 3B5.2-antistoffet og/eller bindingen av antistoffet til Sp35 ble påvirket av fjerningen av glykosyleringsstedet, ble en 3B5.2-variant konstruert. Mer spesifikt ble Kabat-posisjonene i FR3 mutert til konsensus-humane og murine kappa-lettkjedesekvensen SGSGSG (SEQ ID NR:418). Resulterende mutant 3B5.2 variabel lettkjede er følgende. De muterte aminosyrene er uthevet og med dobbel understreking: QIVLTQSPAI MSASPGEKVT MTCSASSRVS YVHWYQQKSG TSPKRWLYDT SNLASGVPAR FSGSGSGTSY SLTISSMEAE DAATYYCQQW STNPPTFGGG TKLEIK (SEQ ID NR:419). [0434] Evnen til både 3B5.2-antistoffet og variant 3B5.2-antistoffet til å binde Sp35-proteinet var lik da de ble testet. I tillegg, basert på elektroforetisk mobilitetsdata ser det ut som 3B5.2 variable lettkjedet glykosyleres, mens det variante lettkjedet ikke gjør det. Til slutt var uttrykksnivåene til begge antistoffene i transfekterte celler like. 30 EKSEMPEL 17 Konstruksjon av humanisert 1A7-antistoff 35 [0435] SEKVENSER TIL 1A7 LETTE OG TUNGE KJEDER Lettekjede: 1 QIVLTQSPAI MSASPGEKVT MTCSAS SSV SYMHWYQQKS GTSPKWIYD TSKLASGVPA RFSGSGSGTS YSLTISSMEA EDAATYYQQ WSSNPFTFGS 100
150 GTKLEIK (SEQ 10 NR:283) Tungkjede: 1 QVQLVQSGPE LKKPGETVKI SCKASGYTFT NYGMNWKQA PGKGLKWMGW INTDTGEPTYT EDFQGRFAFS LETSASTVYL QFNNLKNEDTATY FCAREGVHF DYWGQGTTVT VSS (SEQ 1D NR 170) 10 Uthevet og understreket: Kabat CDR-rester Kursiv og understreket: Cbotia CDR-rester Kanonikale rester Nummereringen er iht. Kabat-planen ANALYSE AV DE MURINE VARIABLE OMRÅDENE [0436] De komplementære bestemmende områdene (CDR-ene) inneholder resten som mest sannsynlig bindes til antigen og må beholdes i det omformede antistoffet. CDR-er defineres etter sekvens iht. Kabat et al (1991) Sequences of Proteins of Immunological Interest. 5th Edition, U.S. Dept. Health and Human Services. U.S. Govt. Printing Office. CDRs fall into canonical classes (Chothia, C., Lesk, A.M., Tramontano, A., Levitt, M., Smith-Gill, S.J., Air, G., Sheriff, S., Padlan, E.A., Davies, D., Tulip, W.R., Colman, P.M., Spinelli, S., Alzari, P.M. and Poljak, R.J. (1989) Nature 342: , der nøkkelrester til en stor grad bestemmer den strukturelle konformasjonen til CDR-sløyfen. Disse restene beholdes nesten alltid i det omformede antistoffet. CDR-ene til lett-+ og tungkjedet ble klassifisert i følgende kanonikale klasser: Lettkjede: Tungkjede L1: 10 rester Klass 1 H1: 5 rester Klasse 1 L2: 7 rester Klasse 1 H2: 17 rester Klasse 2 L3: 9 rester Klasse 1 H3: 7 rester Ingen kanonikal klasse De kanonikale restene som er viktige ved disse CDR-klassene vises i tabell 10. TABELL L1 klasse 1 2(I) 25(A) 30(V) 33(M) 71(Y) L2 klasse 1 48(I) 51(T) 52(S) 64(G) L3 klasse 1 90(Q) 95 (P) HI klasse 1 24(A), 26 (G), 27(F), 29(F), 34(M), 94(R) H2 klasse 2 52a(T) 55(G) 71(L) H3 Ingen kanonikal klasse [0437] De variable lett- og tungkjedene sammenlignes med konsensus (Kabat et al, 1991) og kimlinjesekvenser (Brensing-Kuppers J, Zocher I, Thiebe R, Zachau HG. (1997). Gene. 191(2):173-81
151 150 5 and Matsuda F, Ishii K, Bourvagnet P, Kuma K, Hayashida H, Miyata T, Honjo T.(1998) J Exp Med. 188(11): ved murine and humane undergrupper ved bruk av BLAST-programmet og internt samlet konsensus og databaser med kimlinje-blastproteinsekvenser. [0438] Det variable lette kjedet er medlem av den murine undergruppen Kappa 6 med 94 % identitet i 109 aminosyreoverlappet og med opphav i den murine kk4-kimlinjen (100 % ID) (se nedenfor) > mukk4 10 [0439] Det variable tunge kjedet er medlem av den murine undergruppen HVMS med 55 % identitet i 132 aminosyreoverlappet og med opphav i den murine VGK6-kimlinjen (92 % ID) (se nedenfor) 15 > muvgk6 20 [0440] Det variable lette kjedet korresponderer med den humane undergruppen Kappa 3 med 67 % identitet i 109 aminosyreoverlappet og er nærmest den humane L6-kimlinjen (64 % ID) (se nedenfor) > hul6
152 151 5 [0441] Det variable tungekjedet korresponderer med den humane undergruppen MHV1 med 59 % identitet i 129 aa aminosyreoverlappet og nærmest den humane huvh7-81-kimlinjen (70% ID)(se nedenfor) > huvh MODELLERING AV STRUKTUREN TIL DE VARIABLE OMRÅDENE [0442] Ved denne humaniseringen bygget vi en modell av dep1a7-variable områdene basert på krystallstrukturene til OKT3 (PDB ID 1 SY6 som ble brukt ved modellering av de lette kjedene)- og TE33 (PDB ID 1 TET som ble brukt ved modellering av de tunge kjedene)-antistoffer. ANALYSE AV OMFORMEDE VARIABLE OMRÅDENE [0443] Vi forsøkte å finne de humanuttrykte sekvensene som lignet mest og som ikke trengte å tilbakemuteres i posisjonene (L4, 38, 43, 44, 58, 62, 65-69, 73, 85, 98 og H2, 4, 36, 39, 43, 45, 69, 70, 74, 92) (se f.eks. det amerikanske patentnr. 6,407,213, og bruke dem som antistoff-rammeverk. Vi brukte en database med interne antistoff-sekvenser som er bearbeidet av fagfolk og forespørselsverktøy for å identifisere egnede mal som er mest like de murine P1A7-sekvensene i
153 152 5 kanonikale rester, grensesnittrester og rester i beleggsområdet for å minimere antall tilbakemuteringer. Kimlinjesekvenser som ble fylt med konsensusrester i FR4 området, ble vurdert separat. Etter at flere rammeverk ble vurdert, ble kimlinjesekvensene hul6 og VH7-81 valgt som akseptorrammeverk til henholdsvis tung- og lettkjede. [0444] Tre versjoner av det lette variable kjedet som er omformet og tre versjoner av det tunge variable kjedet som er omformet, er blitt utformet. Den første versjonen inneholdt færrest tilbakemuteringer og den tredje versjonen inneholdt flest (dvs. den som er minst «humanisert»). 10 TILBAKEMUTERINGER I OMFORMET VL hul [0445] E1Q Q1 peker mot antigen og ladeendring kan endre bindingen. Finnes i versjon 2 og 3 L46R R46 er et uvanlig rest på VH/VL-grensesnittet som støtter CDR-L1 og CDR-H3. Finnes i alle versjonene L47W -W47 er plassert i en klynge under CDR-L2. Finnes kun i versjon 3 I58V -V58 er plassert under en klynge under CDR-L2. Finnes kun i versjon 3 F71 Y -Y71 er en kanonikal rest som er viktig ved støtte av CDR-Ll og CDR-L3. Finnes i alle versjonene TILBAKEMUTERINGER I OMFORMET VH huvh [0446] P38K K38 støtter CDR-H2. Finnes i versjon 2 og 3 E46K K46 støtter CDR-H2. Finnes i versjon 2 og 3 M71L L71 er en kanonikal rest som støtter CDR-H1. Finnes i alle versjonene A78V V78 er hypermutert fra kimlinje A og støtter CDR-H1 I82F F82 er en kjernepakkingsrest. Finnes kun i versjon 3 Y91F F91 er en rest på VH/NL-grensesnittet. Finnes kun i versjon 3 HUMANISERINGSUTFORMINGER FOR P1A7 35 [0447] Rammeverk tatt fra sekvensene: Lettkjede: hul6 Tungkjede: huvh7-81 Tilbakemutering er med små bokstaver og uthevet CDR-er er understreket
154 153 5 POLYPEPTID MED FULL LENGDE OG POLYNUKLEOTID TUNGKJEDE SEKVENS FOR tung variant 2 DNA-sekvens til hup1a7-igg1 H2 tungkjede (pxw465)
155 154 NO/EP
156 155 (SEQ ID NR:461) 5 Anslått proteinsekvens til hup1a7 H2 tungkjede (signalsekvens er understreket)
157 HNHYTQKSLS LSPG* (SEQ ID NR:462 POLYPEPTID MED FULL LENGDE OG POLYNUKLEOTID TUNGKJEDE SEKVENS FOR lett variant 1 10 DNA-sekvens til hup1a7 L1 kappa lettkjede (PXW480)
158 AGTGTTAG (SEQ ID NR:463) 5 Anslått proteinsekvens til hup1a7 L1 lettkjede (signalsekvens er understreket) 201 TLSKADYEKH KVYACEVTHQ GLSSPVTKSF NRGEC* (SEQ ID NR:464)
159 158 POLYPEPTID MED FULL LENGDE OG POLYNUKLEOTID TUNGKJEDE SEKVENS FOR lett variant 2 5 DNA-sekvens til hup1a7 L2 kappa lettkjede (PXW476) AGTGTTAG (SEQ ID NR:465) Anslått proteinsekvens til hup1a7 L2 lettkjede (signalsekvens er understreket)
160 TLSKADYEKH KVYACEVTHQ GLSSPVTKSF NRGEC* (SEQ ID NR:466) [0215] Tung- og lettkjede-immunoglobulin polypeptid- og polynukleotidsekvens til et murint og humant kimærisk 1A7-antistoff følger: DNA-sekvens til chp1a7 kappa lettkjede (peag2110)
161 AGTGTTAG (SEQ ID NR:467) Anslått proteinsekvens til chp1a7 lettkjede (signalsekvens er understreket) TLSKADYEKHKVYACEVTHQ GLSSPVTKSF NRGEC* (SEQ ID NR:468) DNA-sekvens til chp1a7 huigg1 tungkjede (peag2112)
162 161 NO/EP
163 CACAACCACT ACACGCAGAA GAGCCTCTCC CTGTCTCCCG GTTGA (SEQ ID NR:469) 5 Anslått proteinsekvens til chp1a7 tungkjede (signalsekvens er understreket) 1 MDWTWRVFCL LAVAPGAHSQ VQLVQSGPEL KKPGETVKlS CKASGYTFTN
164 HNHYTQKSLS LSPG* (SEQ ID NR:470) 5 EKSEMPEL 18 Omstrukturering av Li33Ig2 for å redusere effektorfunksjonen, glysering og aggregering 10 [0449] Forskjellige muteringer ble laget i Li33 for å muligens redusere effektorfunksjon, glysering og aggregering. Effekten av hver av disse muteringene på proteinuttrykk, oppløselighet, antistoffaktivitet i oligodendrocytt-drg-ko-kulturanalyse og glysering eller CD32-binding ble bestemt. Resultatene er summert nedenfor i tabell 11. TABELL 11: LI33IG2-OMSTRUKTURERING 15 Effektor-funksjon Konstruksjon uttrykt Oppløselighet (mg/ml) IC50 CD32 Binding (μg/ml) Aktivitets- Analyse (ko-kultur) Li33Ig2wt J > 20 4,3 + Li33Ig2 agly J 0, Li33Ig2 Rinat J > 5 9,5 + Li33Ig2 PDL J 5,8 > Li33Ig2 Alexion Underveis Ikke bestemt Ikke bestemt Ikke bestemt Glysering Konstruksjon uttrykt Oppløselighet (mg/ml) % glysering Aktivitets- Analyse (ko-kultur) Li33wt J > Li33Ig2 PDL J 5,8 15(5) +
 % glysering Aktivitets- Analyse (ko-kultur) Li33Ig1a94V 157P J Ikke bestemt Ikke bestemt + Li33Ig1a94V 157S J Ikke bestemt Ikke bestemt +")
165 164 Li33Ig2 PDLW94G J Ikke bestemt Ikke bestemt - Li33Ig2 PDLW94V J Ikke bestemt > 2 + Li33Ig2 PDLW94Q J Ikke bestemt < 2 - Li33Ig2 PDLW93N J Ikke bestemt < 2 - Li33Ig2 PDLK93R J 0,4 < 2 + Aggregering Konstruksjon uttrykt Oppløselighet (mg/ml) % glysering Aktivitets- Analyse (ko-kultur) Li33Ig1a94V 157P J Ikke bestemt Ikke bestemt + Li33Ig1a94V 157S J Ikke bestemt Ikke bestemt + Li33Ig1a94V 157T J Ikke bestemt Ikke bestemt - Li33Ig1a94V 157V J Ikke bestemt Ikke bestemt + Li33Ig1a94V 157S J Ikke bestemt Ikke bestemt + Li33Ig1a94V 157A J Ikke bestemt Ikke bestemt + Li33Ig2PDL94V W103Q Li33Ig2PDL94V W103A Li33Ig2PDL94V 103Q57S Li33Ig2PDL94V 103Q57A J Ikke bestemt Ikke bestemt - J Ikke bestemt Ikke bestemt - J Ikke bestemt < 2 + J Ikke bestemt < 2 + EKSEMPEL 19 5 Konstruksjon av en Li81-variant [0450] Antistoff Li81 er en affinitetsmodnet versjon av antistoff Li13. En aglykosylatert versjon av Li81-antistoffet ble laget ved å endre en enkel aminosyre i Li81-tungkjedesekvensen. Følgende er aminosyresekvensen til det variable tunge kjedet (VH) til den aglykosylerte varianten [0451] Ledesekvensen (de første 19 aminosyrene), som ikke vil være tilstede i det modne proteinet, vises uthevet og CDRS-ene er understreket. Endringen av det enkle aminosyren i forhold til Li81 variabel sekvens med tungkjede (SEQ ID NR: 433) vises uthevet og med to understrekinger. Følgende er nukleotidsekvensen til det variable tunge kjedet (VH) til den aglykosylerte varianten.

166 165 NO/EP
167 166 PATENTKRAV Et isolert antistoff eller et antigenbindende fragment av det som kan spesifikt bindes til et Sp35-polypeptid der antistoffet eller fragmentet av det omfatter et VH-område som omfatter VH CDR1-, CDR2- og CDR3-sekvenser av henholdsvis SEQ ID NR:436, SEQ ID NR:437 og SEQ ID NR:438 og et VL-område som omfatter VL CDR1-, CDR2- og CDR3-sekvenser av henholdsvis SEQ ID NR:442, SEQ ID NR:443 og SEQ ID NR:444 og der antistoffet eller fragmentet derav binder seg til et Sp35-polypeptid med affinitet som karakteriseres av en dissosiasjonskonstant som ikke er større enn 5 x M som målt med FACS på CHO-celler som er stabilt transfektert med human Sp Antistoffet eller fragmentet derav ved krav 1, der antistoffet omfatter aminosyresekvensen som fremlegges i SEQ ID NR:434 og aminosyresekvensen som fremlegges i SEQ ID NR: Antistoffet eller fragmentet derav ved krav 1, der antistoffet omfatter den modne formen av aminosyresekvensen til SEQ ID NR: 474 og som alternativ, aminosyresekvensen som fremlegges i SEQ ID NR: Antistoffet eller fragmentet derav i hvilket som helst krav 1-3, som er en antagonist av Sp35- mediert nevronal celledød. 5. Antistoffet eller fragmentet derav i hvilket som helst krav 1-3, som er en antagonist av Sp35- mediert myelineringshemming Antistoffet eller fragmentet derav i hvilket som helst krav 1-3, som er en antagonist av Sp35- mediert oligodendrocytt-differensieringshemming Bruken av isolert Sp 35-antistoff eller fragment derav i hvilket som helst krav 1-3, ved preparering av et medikament til behandling av en CNS-skade i et dyr. 8. Bruken av isolert Sp35-antistoff eller fragment derav i hvilket som helst krav 1-3, ved preparering av et medikament til behandling et en sykdom som velges fra gruppen som består av: multippel sklerose (MS), progressiv multifokal leukoencefalopati (PML), enkefalomyelitt (EPL), sentral pontin myelolyse (CPM), adrenoleukodystrofi, Alexanders sykdom, Pelizaeus Merzbacher-sykdom (PMZ), globoid celleleukodystrofi (Krabbes sykdom) og Wallerisk degenerasjon, optikusnevritt, transversell myelitt, amyotrofisk-lateralsklerose (ALS), Huntingtons sykdom, Alzheimers sykdom, Parkinsons sykdom, ryggmargsskade, traumatisk hjerneskade, skader etter stråling, nevrologiske komplikasjoner ved kjemoterapi, slag, akutt iskemisk optisk nevropati, vitamin E-mangel, isolert vitamin E-mangel syndrom, AR, Bassen-
168 167 Kornzweig-syndrom, Marchiafava-Bignami-syndrom, metakromatisk leukodystrofi, trigeminusnevralgi og ansiktslammelse i et dyr Bruken i krav 8, der sykdommen eller lidelsen er multippel sklerose. 10. Det isolert Sp35-antistoffet eller fragment derav i hvilket som helst krav 1-3, til bruk ved behandling av en CNS-skade i et dyr Det isolerte Sp35-antistoffet eller fragment derav i hvilket som helst krav 1-3, til bruk ved behandling av en sykdom som velges fra gruppen som består av: multippel sklerose (MS), progressiv multifokal leukoencefalopati (PML), enkefalomyelitt (EPL), sentral pontin myelolyse (CPM), adrenoleukodystrofi, Alexanders sykdom, Pelizaeus Merzbacher-sykdom (PMZ), globoid celleleukodystrofi (Krabbes sykdom) og Wallerisk degenerasjon, optikusnevritt, transversell myelitt, amyotrofisk-lateralsklerose (ALS), Huntingtons sykdom, Alzheimers sykdom, Parkinsons sykdom, ryggmargsskade, traumatisk hjerneskade, skader etter stråling, nevrologiske komplikasjoner ved kjemoterapi, slag, akutt iskemisk optisk nevropati, vitamin E- mangel, isolert vitamin E-mangel syndrom, AR, Bassen-Kornzweig-syndrom, Marchiafava- Bignami-syndrom, metakromatisk leukodystrofi, trigeminusnevralgi og ansiktslammelse i et dyr. 12. Det isolerte Sp35-antistoffet eller fragmentet derav i krav 11, der sykdommen eller lidelsen er multippel sklerose En in vitro metode for å hemme signaltransduksjon av NgR1, som omfatter NgR1 med en effektiv mengde isolert Sp35-antistoff eller fragment derav i hvilket som helst krav En vertscelle som omfatter: (a) to uttrykksvektorer, der den første uttrykksvektoren koder VH-området til antistoffet eller fragmentet derav i hvilket som helst krav 1-3, og den andre uttrykksvektoren koder VL-området til antistoffet eller fragmentet av hvilket som helst krav 1-3; eller (b) en enkel uttrykksvektor som koder både VH-området til antistoffet eller fragmentet derav i hvilket som helst krav 1-3, og VL-området til antistoffet eller fragmentet av hvilket som helst krav 1-3
169 1 SEKVENSLISTE <110> Biogen Idec MA Inc. <120> SP35 Antistoffer og bruken av dem <130> PC01 <150> US 60/879,324 <151> <160> 474 <170> Patent i versjon 3.3 <210> 1 <211> 1845 <212> DNA <213> Homo sapiens <400> 1
170 2 <210> 2 <211> 614 <213> Homo sapiens <400> 2
171 3 NO/EP
172 4 NO/EP
173 5 <210> 3 <211> 6 <213> Homo sapiens <400> 3 Met Gln Val Ser Lys Arg 1 5 <210> 4 <211> 29 <212> DNA <223> Syntetisk primer brukt for å bestemme det lette kjedet til P1E11.3B7 <400> 4 gcgtctagaa ctggatggtg ggagatgga 29 <210> 5 <211> 15 <212> DNA <223> Syntetisk sekvens Li10-antistoff som koder for VH-CDR1 <400> 5 acttacccta tggtt 15 <210> 6 <211> 5 <223> Syntetisk Syntetisk sekvens Li10-antistoff som koder for VH-CDR1 <400> 6 Thr Tyr Pro Met Val 1 5 <210> 7 <211> 51 <212> DNA <223> Syntetisk sekvens Li10-antistoff som koder for VH-CDR2 <400> 7
174 6 tggatcggtc cttctggtgg cgttactgct tatgctgact ccgttaaagg t 51 <210> 8 <211> 17 <223> Syntetisk sekvens Li10-antistoff som koder for VH-CDR2 <400> 8 Trp Ile Gly Pro Ser Gly Gly Val Thr Ala Tyr Ala Asp Ser Val Lys Gly <210> 9 <211> 33 <212> DNA <223> Syntetisk sekvens Li10-antistoff som koder for VH-CDR3 <400> 9 ccctatagca gtggctggtg ggacttcgat ctc 33 <210> 10 <211> 11 <223> Syntetisk sekvens Li10-antistoff som koder for VH-CDR3 <400> 10 Pro Tyr Ser Ser Gly Trp Trp Asp Phe Asp Leu <210> 11 <211> 15 <212> DNA <223> Syntetisk sekvens Li07-antistoff som koder for VH-CDR1 <400> 11 atgtacttta tgggt 15 <210> 12 <211> 5
175 7 <223> Syntetisk sekvens Li07-antistoff som koder for VH-CDR1 <400> 12 Met Tyr Phe Met Gly 1 5 <210> 13 <211> 51 <212> DNA <223> Syntetisk sekvens Li07-antistoff som koder for VH-CDR2 <400> 13 tctatctctc cttctggtgg ctttacttct tatgctgact ccgttaaagg t 51 <210> 14 <211> 17 <223> Syntetisk sekvens Li07-antistoff som koder for VH-CDR2 <400> 14 Ser Ile Ser Pro Ser Gly Gly Phe Thr Ser Tyr Ala Asp Ser Val Lys Gly <210> 15 <211> 21 <212> DNA <223> Syntetisk sekvens Li07-antistoff som koder for VH-CDR3 <400> 15 gatcggcatg cttttgatat c 21 <210> 16 <211> 7 <223> Syntetisk sekvens Li07-antistoff som koder for VH-CDR3
176 8 <400> 16 Asp Arg His Ala Phe Asp Ile 1 5 <210> 17 <211> 14 <212> DNA <223> Syntetisk sekvens Li05-antistoff som koder for VH-CDR1 <400> 17 cttacgctat gggt 14 <210> 18 <211> 5 <223> Syntetisk sekvens fra Li05-antistoff som koder for VH-CDR1 <400> 18 Ala Tyr Ala Met Gly 1 5 <210> 19 <211> 51 <212> DNA <223> Syntetisk sekvens fra LiD5-antistoff som koder for VH-CDR2 <400> 19 tctatcgttt cttctggtgg ctatactgat tatgctgact ccgttaaagg t 51 <210> 20 <211> 17 <223> Syntetisk sekvens fra Li05-antistoff som koder for VH-CDR2 <400> 20 Ser Ile Val Ser Ser Gly Gly Tyr Thr Asp Tyr Ala Asp Ser Val Lys Gly
177 9 <210> 21 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li05-antistoff som koder for VH-CDR3 <400> 21 gagggtgacc ataatgcttt tgatatc 27 <210> 22 <211> 9 <223> Syntetisk sekvens fra Li05-antistoff som koder for VH-CDR3 <400> 22 Glu Gly Asp His Asn Ala Phe Asp Ile 1 5 <210> 23 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li11-antistoff som koder for VH-CDR1 <400> 23 tcttacgcta tgtat 15 <210> 24 <211> 5 <223> Syntetisk sekvens fra Li11-antistoff som koder for VH-CDR1 <400> 24 Ser Tyr Ala Met Tyr 1 5 <210> 25 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li11-antistoff som koder for VH-CDR2
178 10 <400> 25 tctatctcta cttctggtggctatactggt tatgctgact ccgttaaagg t 51 <210> 26 <211> 17 <223> Syntetisk sekvens fra Li11-antistoff som koder for VH-CDR2 <400> 26 Ser Ile Ser Thr Ser Gly Gly Tyr Thr Gly Tyr Ala Asp Ser Val Lys Gly <210> 27 <211> 36 <212> DNA <223> Syntetisk sekvens fra Li11-antistoff som koder for VH-CDR3 <400> 27 gataccagcg ataatgacta ctactacatg gacgtc 36 <210> 28 <211> 12 <223> Syntetisk sekvens fra Li11-antistoff som koder for VH-CDR3 <400> 28 Asp Thr Ser Asp Asn Asp Tyr Tyr Tyr Met Asp Val <210> 29 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li01-antistoff som koder for VH-CDR1 <400> 29 aagtaccaga tgact 15
179 11 <210> 30 <211> 5 <223> Syntetisk sekvens fra Li01-antistoff som koder for VH-CDR1 <400> 30 Lys Tyr Gln Met Thr 1 5 <210> 31 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li01-antistoff som koder for VH-CDR2 <400> 31 tctatctatc cttctggtgg caatactgtt tatgctgact ccgttaaagg t 51 <210> 32 <211> 17 <223> Syntetisk sekvens fra Li01-antistoff som koder for VH-CDR2 <400> 32 Ser Ile Tyr Pro Ser Gly Gly Asn Thr Val Tyr Ala Asp Ser Val Lys Gly <210> 33 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li01-antistoff som koder for VH-CDR3 <400> 33 gggactacag aggcagtctt tgactac 27 <210> 34 <211> 9
180 12 <223> Syntetisk sekvens fra Li01-antistoff som koder for VH-CDR3 <400> 34 Gly Thr Thr Glu Ala Val Phe Asp Tyr 1 5 <210> 35 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li12-antistoff som koder for VH-CDR1 <400> 35 cagtacaata tgttt 15 <210> 36 <211> 5 <223> Syntetisk sekvens fra Li12-antistoff som koder for VH-CDR1 <400> 36 Gln Tyr Asn Met Phe 1 5 <210> 37 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li12-antistoff som koder for VH-CDR2 <400> 37 cgtatctctt cttctggtgg catgactatg tatgctgact ccgttaaagg t 51 <210> 38 <211> 17 <223> Syntetisk sekvens fra Li12-antistoff som koder for VH-CDR2 <400> 38 Arg Ile Ser Ser Ser Gly Gly Met Thr Met Tyr Ala Asp Ser Val Lys
181 13 Gly <210> 39 <211> 69 <212> DNA <223> Syntetisk sekvens fra Li12-antistoff som koder for VH-CDR3 <400> 39 gaagcgttac ggccttattg tagtggtggt agctgctact ccgactacta ctactacggt 60 atggacgtc 69 <210> 40 <211> 23 <223> Syntetisk sekvens fra Li12-antistoff som koder for VH-CDR3 <400> 40 Glu Ala Leu Arg Pro Tyr Cys Ser Gly Gly Ser Cys Tyr Ser Asp Tyr Tyr Tyr Tyr Gly Met Asp Val 20 <210> 41 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li06-antistoff som koder for VH-CDR1 <400> 41 gagtacccta tggat 15 <210> 42 <211> 5 <223> Syntetisk sekvens fra Li06-antistoff som koder for VH-CDR1 <400> 42 Glu Tyr Pro Met Asp 1 5
182 14 <210> 43 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li06-antistoff som koder for VH-CDR2 <400> 43 tctatctatt cttctggtgg ctctactgtt tatgctgact ccattaaagg t 51 <210> 44 <211> 17 <223> Syntetisk sekvens fra Li06-antistoff som koder for VH-CDR2 <400> 44 Ser Ile Tyr Ser Ser Gly Gly Ser Thr Val Tyr Ala Asp Ser Ile Lys Gly <210> 45 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li06-antistoff som koder for VH-CDR3 <400> 45 gagggtgact ctgatgcttt tgatatc 27 <210> 46 <211> 9 <223> Syntetisk sekvens fra Li06-antistoff som koder for VH-CDR3 <400> 46 Glu Gly Asp Ser Asp Ala Phe Asp Ile 1 5 <210> 47 <211> 15 <212> DNA
183 15 <223> Syntetisk sekvens fra Li08-antistoff som koder for VH-CDR1 <400> 47 cattacgaga tggtt 15 <210> 48 <211> 5 <223> Syntetisk sekvens fra Li08-antistoff som koder for VH-CDR1 <400> 48 His Tyr Glu Met Val 1 5 <210> 49 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li08-antistoff som koder for VH-CDR2 <400> 49 tctatccgtt cttctggtgg cgctactaag tatgctgact ccgttaaagg t 51 <210> 50 <211> 17 <223> Syntetisk sekvens fra Li08-antistoff som koder for VH-CDR2 <400> 50 Ser Ile Arg Ser Ser Gly Gly Ala Thr Lys Tyr Ala Asp Ser Val Lys Gly <210> 51 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li08-antistoff som koder for VH-CDR3 <400> 51
184 16 gagtcgccag acgactactt tgactac 27 <210> 52 <211> 9 <223> Syntetisk sekvens fra Li08-antistoff som koder for VH-CDR3 <400> 52 Glu Ser Pro Asp Asp Tyr Phe Asp Tyr 1 5 <210> 53 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li03-antistoff som koder for VH-CDR1 <400> 53 cagtacccta tggag 15 <210> 54 <211> 5 <223> Syntetisk sekvens fra Li03-antistoff som koder for VH-CDR1 <400> 54 Gln Tyr Pro Met Glu 1 5 <210> 55 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li03-antistoff som koder for VH-CDR2 <400> 55 ggtatctatc cttctggtgg ctctactgtt tatgctgact ccgttaaagg t 51 <210> 56 <211> 17
185 17 <223> Syntetisk sekvens fra Li03-antistoff som koder for VH-CDR2 <400> 56 Gly Ile Tyr Pro Ser Gly Gly Ser Thr Val Tyr Ala Asp Ser Val Lys Gly <210> 57 <211> 30 <212> DNA <223> Syntetisk sekvens fra Li03-antistoff som koder for VH-CDR3 <400> 57 gcggggcagt ggctggggga ctttgactac 30 <210> 58 <211> 10 <223> Syntetisk sekvens fra Li03-antistoff som koder for VH-CDR3 <400> 58 Ala Gly Gln Trp Leu Gly Asp Phe Asp Tyr <210> 59 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li09-antistoff som koder for VH-CDR1 <400> 59 atgtactcta tggtt 15 <210> 60 <211> 5 <223> Syntetisk sekvens fra Li09-antistoff som koder for VH-CDR1 <400> 60 Met Tyr Ser Met Val 1 5
186 18 <210> 61 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li09-antistoff som koder for VH-CDR2 <400> 61 tatatctctc cttctggtgg caagactatg tatgctgact ccgttaaagg t 51 <210> 62 <211> 17 <223> Syntetisk sekvens fra Li09-antistoff som koder for VH-CDR2 <400> 62 Tyr Ile Ser Pro Ser Gly Gly Lys Thr Met Tyr Ala Asp Ser Val Lys Gly <210> 63 <211> 69 <212> DNA <223> Syntetisk sekvens fra Li09-antistoff som koder for VH-CDR3 <400> 63 gattcgagac gccggtatta cgatttttgg agtggttatc acaactacta ctactactac 60 atggacgtc 69 <210> 64 <211> 23 <223> Syntetisk sekvens fra Li09-antistoff som koder for VH-CDR3 <400> 64 Asp Ser Arg Arg Arg Tyr Tyr Asp Phe Trp Ser Gly Tyr His Asn Tyr
187 19 Tyr Tyr Tyr Tyr Met Asp Val 20 <210> 65 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li04-antistoff som koder for VH-CDR1 <400> 65 cgttacaata tgggt 15 <210> 66 <211> 5 <223> Syntetisk sekvens fra Li04-antistoff som koder for VH-CDR1 <400> 66 Arg Tyr Asn Met Gly 1 5 <210> 67 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li04-antistoff som koder for VH-CDR2 <400> 67 gttatctatc cttctggtgg cggtactcat tatgctgact ccgttaaagg t 51 <210> 68 <211> 17 <223> Syntetisk sekvens fra Li04-antistoff som koder for VH-CDR2 <400> 68 Val Ile Tyr Pro Ser Gly Gly Gly Thr His Tyr Ala Asp Ser Val Lys Gly <210> 69
188 20 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li04-antistoff som koder for VH-CDR3 <400> 69 tctatagcag atgatgcttt tgatatc 27 <210> 70 <211> 9 <223> Syntetisk sekvens fra Li04-antistoff som koder for VH-CDR3 <400> 70 Ser Ile Ala Asp Asp Ala Phe Asp Ile 1 5 <210> 71 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li02-antistoff som koder for VH-CDR1 <400> 71 acttacgaga tgatt 15 <210> 72 <211> 5 <223> Syntetisk sekvens fra Li02-antistoff som koder for VH-CDR1 <400> 72 Thr Tyr Glu Met Ile 1 5 <210> 73 <211> 48 <212> DNA <223> Syntetisk sekvens fra Li02-antistoff som koder for VH-CDR2 <400> 73 tctatcggtc cttctggtgg ccttacttgg tatgctgact ccgttaaa 48
189 21 <210> 74 <211> 17 <223> Syntetisk sekvens fra Li02-antistoff som koder for VH-CDR2 <400> 74 Ser Ile Gly Pro Ser Gly Gly Leu Thr Trp Tyr Ala Asp Ser Val Lys Gly <210> 75 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li02-antistoff som koder for VH-CDR3 <400> 75 atgtattact gtgtacggat tgatgatagt agtggttggg cttttgatat c 51 <210> 76 <211> 17 <223> Syntetisk sekvens fra Li02-antistoff som koder for VH-CDR3 <400> 76 Met Tyr Tyr Cys Val Arg Ile Asp Asp Ser Ser Gly Trp Ala Phe Asp Ile <210> 77 <211> 5 <223> Syntetisk sekvens fra 1A7-antistoff som koder for VH-CDR1 <400> 77 Asn Tyr Gly Met Asn 1 5
190 22 <210> 78 <211> 17 <223> Syntetisk sekvens fra 1A7-antistoff som koder for VH-CDR2 <400> 78 Trp Ile Asn Thr Asp Thr Gly Glu Pro Thr Tyr Thr Glu Asp Phe Gln Gly <210> 79 <211> 7 <223> Syntetisk sekvens fra 1A7-antistoff som koder for VH-CDR3 <400> 79 Glu Gly Val His Phe Asp Tyr 1 5 <210> 80 <211> 7 <223> Syntetisk sekvens fra 2F3-antistoff som koder for VH-CDR1 <400> 80 Phe Ser Asp Ala Trp Leu Asp 1 5 <210> 81 <211> 19 <223> Syntetisk sekvens fra 2F3-antistoff som koder for VH-CDR2 <400> 81 Glu Ile Arg Ser Lys Ala Asn Asn His Ala Thr Asn Tyr Ala Glu Ser
191 23 Val Lys Gly <210> 82 <211> 4 <223> Syntetisk sekvens fra 2F3-antistoff som koder for VH-CDR3 <400> 82 Ser Phe Ala Tyr 1 <210> 83 <211> 5 <223> Syntetisk sekvens fra 3P1D10.2C3 eller 3P1E11.3B7-antistoff som koder koder for VH-CDR1 <400> 83 Ser Ser Trp Thr Gln 1 5 <210> 84 <211> 17 <223> Syntetisk sekvens fra 3P1D10.2C3 eller 3P1E11.3B7-antistoff som koder for VH-CDR2 <400> 84 Ala Ile Tyr Pro Gly Asp Gly Asp Thr Arg Tyr Thr Gln Lys Phe Lys Gly <210> 85 <211> 8 <223> Syntetisk sekvens fra 3P1D10.2C3 eller 3P1E11.3B7-antistoff som koder for VH-CDR VH-CDR3
192 24 <400> 85 His Asn Ser Tyr Gly Met Asp Tyr 1 5 <210> 86 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li10-antistoff som koder for VL-CDR1 <400> 86 cgggcgagtc agggtattgg caactggtta gcc 33 <210> 87 <211> 11 <223> Syntetisk sekvens fra Li10-antistoff som koder for VL-CDR1 <400> 87 Arg Ala Ser Gln Gly Ile Gly Asn Trp Leu Ala <210> 88 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li10-antistoff som koder for VL-CDR2 <400> 88 gctgcatcca gtttggaaag t 21 <210> 89 <211> 7 <223> Syntetisk sekvens fra Li10-antistoff som koder for VL-CDR2 <400> 89 Ala Ala Ser Ser Leu Glu Ser 1 5 <210> 90
193 25 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li10-antistoff som koder for VL-CDR3 <400> 90 caacaggctc agactttccc gctcacc 27 <210> 91 <211> 9 <223> Syntetisk sekvens fra Li10-antistoff som koder for VL-CDR3 <400> 91 Gln Gln Ala Gln Thr Phe Pro Leu Thr 1 5 <210> 92 <211> 33 <212> DNA <223> Syntetisk sekvens Li07-antistoff som koder for VL-CDR1 <400> 92 tctggagatc agttgggtga caaacatgtg gct 33 <210> 93 <211> 11 <223> Syntetisk sekvens Li07-antistoff som koder for VL-CDR1 <400> 93 Ser Gly Asp Gln Leu Gly Asp Lys His Val Ala <210> 94 <211> 21 <212> DNA <223> Syntetisk sekvens Li07-antistoff som koder for VL-CDR2 <400> 94 ctagacatta agaggcccgc a 21
194 26 <210> 95 <211> 7 <223> Syntetisk sekvens Li07-antistoff som koder for VL-CDR2 <400> 95 Leu Asp Ile Lys Arg Pro Ala 1 5 <210> 96 <211> 24 <212> DNA <223> Syntetisk sekvens Li07-antistoff som koder for VL-CDR3 <400> 96 caggcgtggg acatcaagac ggtc 24 <210> 97 <211> 8 <223> Syntetisk sekvens Li07-antistoff som koder for VL-CDR3 <400> 97 Gln Ala Trp Asp Ile Lys Thr Val 1 5 <210> 98 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li05-antistoff som koder for VL-CDR1 <400> 98 gggggagaca acattggaag taagagtgtc cac 33 <210> 99 <211> 11 <223> Syntetisk sekvens fra Li05-antistoff som koder for VL-CDR1
195 27 <400> 99 Gly Gly Asp Asn Ile Gly Ser Lys Ser Val His <210> 100 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li05-antistoff som koder for VL-CDR2 <400> 100 gatgattatg accggccctc a 21 <210> 101 <211> 7 <223> Syntetisk sekvens fra Li05-antistoff som koder for VL-CDR2 <400> 101 Asp Asp Tyr Asp Arg Pro Ser 1 5 <210> 102 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li05-antistoff som koder for VL-CDR3 <400> 102 caggtgaggg acagccgtac tgaggaacgg gtg 33 <210> 103 <211> 11 <223> Syntetisk sekvens fra Li05-antistoff som koder for VL-CDR3 <400> 103 Gln Val Arg Asp Ser Arg Thr Glu Glu Arg Val <210> 104 <211> 33
196 28 <212> DNA <223> Syntetisk sekvens fra Li11-antistoff som koder for VL-CDR1 <400> 104 cgggcgagtc aggagattgc caactactta gcc 33 <210> 105 <211> 11 <223> Syntetisk sekvens fra Li11-antistoff som koder for VL-CDR1 <400> 105 Arg Ala Ser Gln Glu Ile Ala Asn Tyr Leu Ala <210> 106 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li11-antistoff som koder for VL-CDR2 <400> 106 gatacataca ctttgcagac t 21 <210> 107 <211> 7 <223> Syntetisk sekvens fra Li11-antistoff som koder for VL-CDR2 <400> 107 Asp Thr Tyr Thr Leu Gln Thr 1 5 <210> 108 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li11-antistoff som koder for VL-CDR3 <400> 108 caacaggctg acattttccc gctctct 27
197 29 <210> 109 <211> 9 <223> Syntetisk sekvens fra Li11-antistoff som koder for VL-CDR3 <400> 109 Gln Gln Ala Asp Ile Phe Pro Leu Ser 1 5 <210> 110 <211> 32 <212> DNA <223> Syntetisk sekvens fra Li01-antistoff som koder for VL-CDR1 <400> 110 caggcgagtc aggacattag caactattta aa 32 <210> 111 <211> 11 <223> Syntetisk sekvens fra Li01-antistoff som koder for VL-CDR1 <400> 111 Gln Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn <210> 112 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li01-antistoff som koder for VL-CDR2 <400> 112 gatgcatcca atttggaaac a 21 <210> 113 <211> 7 <223> Syntetisk sekvens fra Li01-antistoff som koder for VL-CDR2
198 30 <400> 113 Asp Ala Ser Asn Leu Glu Thr 1 5 <210> 114 <211> 30 <212> DNA <223> Syntetisk sekvens fra Li01-antistoff som koder for VL-CDR3 <400> 114 caacaggctg acaggttccc tgcggtcact 30 <210> 115 <211> 10 <223> Syntetisk sekvens fra Li01-antistoff som koder for VL-CDR3 <400> 115 Gln Gln Ala Asp Arg Phe Pro Ala Val Thr <210> 116 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li06-antistoff som koder for VL-CDR1 <400> 116 cgggccagtc agagtattag tagctggttg gcc 33 <210> 117 <211> 11 <223> Syntetisk sekvens fra Li06-antistoff som koder for VL-CDR1 <400> 117 Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala <210> 118 <211> 21 <212> DNA
199 31 <223> Syntetisk sekvens fra Li06-antistoff som koder for VL-CDR2 <400> 118 gctgcatcca gtttacgaac t 21 <210> 119 <211> 7 <223> Syntetisk sekvens fra Li06-antistoff som koder for VL-CDR2 <400> 119 Ala Ala Ser Ser Leu Arg Thr 1 5 <210> 120 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li06-antistoff som koder for VL-CDR3 <400> 120 ctacaagatt acagttaccc tctcact 27 <210> 121 <211> 9 <223> Syntetisk sekvens fra Li06-antistoff som koder for VL-CDR3 <400> 121 Leu Gln Asp Tyr Ser Tyr Pro Leu Thr 1 5 <210> 122 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li08-antistoff som koder for VL-CDR1 <400> 122 caggcgagtc aggacattag ttactattta aat 33
200 32 <210> 123 <211> 11 <223> Syntetisk sekvens fra Li08-antistoff som koder for VL-CDR1 <400> 123 Gln Ala Ser Gln Asp Ile Ser Tyr Tyr Leu Asn <210> 124 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li08-antistoff som koder for VL-CDR2 <400> 124 gatgtatcca atttgcaaac a 21 <210> 125 <211> 7 <223> Syntetisk sekvens fra Li08-antistoff som koder for VL-CDR2 <400> 125 Asp Val Ser Asn Leu Gln Thr 1 5 <210> 126 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li05-antistoff som koder for VL-CDR3 <400> 126 caacagtctg ataatctccc tctcact 27 <210> 127 <211> 9 <223> Syntetisk sekvens fra Li08-antistoff som koder for VL-CDR3 <400> 127
201 33 Gln Gln Ser Asp Asn Leu Pro Leu Thr 1 5 <210> 128 <211> 32 <212> DNA <223> Syntetisk sekvens fra Li03-antistoff som koder for VL-CDR1 <400> 128 gggcaagtca gagcattagc agctatttaa at 32 <210> 129 <211> 11 <223> Syntetisk sekvens fra Li03-antistoff som koder for VL-CDR1 <400> 129 Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn <210> 130 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li03-antistoff som koder for VL-CDR2 <400> 130 gctgcatcca gtttgcaaag t 21 <210> 131 <211> 7 <223> Syntetisk sekvens fra Li03-antistoff som koder for VL-CDR2 <400> 131 Ala Ala Ser Ser Leu Gln Ser 1 5 <210> 132 <211> 27 <212> DNA
202 34 <223> Syntetisk sekvens fra Li03-antistoff som koder for VL-CDR3 <400> 132 caacagagtt acagtacccc gtggacg 27 <210> 133 <211> 9 <223> Syntetisk sekvens fra Li03-antistoff som koder for VL-CDR3 <400> 133 Gln Gln Ser Tyr Ser Thr Pro Trp Thr 1 5 <210> 134 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li09-antistoff som koder for VL-CDR1 <400> 134 cgcgcaagtc agagcatcga cacctattta aat 33 <210> 135 <211> 11 <223> Syntetisk sekvens fra Li09-antistoff som koder for VL-CDR1 <400> 135 Arg Ala Ser Gln Ser Ile Asp Thr Tyr Leu Asn <210> 136 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li09-antistoff som koder for VL-CDR2 <400> 136 gctgcatcca agttggaaga c 21 <210> 137
203 35 <211> 7 <21~> Kunstig sekvens <223> Syntetisk sekvens fra Li09-antistoff som koder for VL-CDR2 <400> 137 Ala Ala Ser Lys Leu Glu Asp 1 5 <210> 138 <211> 26 <212> DNA <223> Syntetisk sekvens fra Li09-antistoff som koder for VL-CDR3 <400> 138 caacagagtt acagtccccc tctcac 26 <210> 139 <211> 9 <223> Syntetisk sekvens fra Li09-antistoff som koder for VL-CDR3 <400> 139 Gln Gln Ser Tyr Ser Pro Pro Leu Thr 1 5 <210> 140 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li02-antistoff som koder for VL-CDR1 <400> 140 tctggagata aattggggga taaatttgct tcc 33 <210> 141 <211> 11 <223> Syntetisk sekvens fra Li02-antistoff som koder for VL-CDR1 <400> 141
204 36 Ser Gly Asp Lys Leu Gly Asp Lys Phe Ala Ser <210> 142 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li02-antistoff som koder for VL-CDR2 <400> 142 caagatagga agcgtctctc a 21 <210> 143 <211> 7 <223> Syntetisk sekvens fra Li02-antistoff som koder for VL-CDR2 <400> 143 Gln Asp Arg Lys Arg Leu Ser 1 5 <210> 144 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li02-antistoff som koder for VL-CDR3 <400> 144 caggcgtggg acaccaacac tgtggtc 27 <210> 145 <211> 9 <223> Syntetisk sekvens fra Li02-antistoff som koder for VL-CDR3 <400> 145 Gln Ala Trp Asp Thr Asn Thr Val Val 1 5 <210> 146 <211> 10
205 37 <223> Syntetisk sekvens fra 1A7-antistoff som koder for VL-CDR1 <400> 146 Ser Ala Ser Ser Ser Val Ser Tyr Met His <210> 147 <211> 7 <223> Syntetisk sekvens fra 1A7-antistoff som koder for VL-CDR2 <400> 147 Asp Thr Ser Lys Leu Ala Ser 1 5 <210> 148 <211> 9 <223> Syntetisk sekvens fra 1A7-antistoff som koder for VL-CDR3 <400> 148 Gln Gln Trp Ser Ser Asn Pro Phe Thr 1 5 <210> 149 <211> 11 <223> Syntetisk sekvens fra 2F3-antistoff som koder for VL~CDR1 <400> 149 Arg Ala Ser Gly Asn Ile Tyr Asn Tyr Leu Ala <210> 150 <211> 7 <223> Syntetisk sekvens fra 2F3-antistoff som koder for VL-CDR2 <400> 150
206 38 Asn Ala Lys Thr Leu Pro Asp 1 5 <210> 151 <211> 9 <223> Syntetisk sekvens fra 2F3-antistoff som koder for VL-CDR3 <400> 151 Gln His Phe Trp Ala Ile Pro Tyr Thr 1 5 <210> 152 <211> 17 <223> Syntetisk sekvens fra 3P1D10.2C3-antistoff som koder for VL-CDR1 <400> 152 Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr <210> 153 <211> 7 <223> Syntetisk sekvens fra 3P1D10.2C3-antistoff som koder for VL-CDR2 <400> 153 Trp Ala Ser Thr Arg Glu Ser 1 5 <210> 154 <211> 10 <223> Syntetisk sekvens fra 3P1D10.2C3-antistoff som koder for VL-CDR3
207 39 <400> 154 Gln Asn Asp Tyr Ser Tyr Pro Leu Phe Thr <210> 155 <211> 17 <213> Kunstig sekvens <223> Syntetisk sekvens fra 3P1E11.3B7-antistoff som koder for VL-CDR1 <400> 155 Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Ser Tyr Leu Thr <210> 156 <211> 7 <223> Syntetisk sekvens fra 3P1E11.3B7-antistoff som koder for VL-CDR2 <400> 156 Trp Ala Ser Thr Arg Glu Ser 1 5 <210> 157 <211> 10 <223> Syntetisk sekvens fra 3P1E11.3B7-antistoff som koder for VL-CDR3 <400> 157 Gln Asn Asp Tyr Ser Tyr Pro Leu Phe Thr <210> 158 <211> 133
208 40 <223> Syntetisk Li02 variabel tungkjede-sekvens <400> 158 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr Glu Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Gly Pro Ser Gly Gly Leu Thr Trp Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys Val Arg Ile Asp Asp Ser Ser Gly Trp Ala Phe Asp Ile Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 159 <211> 145 <223> Syntetisk Li09 variabel tungkjede-sekvens <400> 159 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Met Tyr Ser Met Val Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
209 41 Ser Tyr Ile Ser Pro Ser Gly Gly Lys Thr Met Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Phe Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Ser Arg Arg Arg Tyr Tyr Asp Phe Trp Ser Gly Tyr His Asn Tyr Tyr Tyr Tyr Tyr Met Asp Val Trp Gly Lys Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 145 <210> 160 <211> 131 <223> Syntetisk Li06 variabel tungkjede-sekvens <400> 160 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Glu Tyr Pro Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Tyr Ser Ser Gly Gly Ser Thr Val Tyr Ala Asp Ser Ile Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
210 42 Ala Arg Glu Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 161 <211> 131 <223> Syntetisk Li05 variabel tungkjede-sekvens <400> 161 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ala Tyr Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Val Ser Ser Gly Gly Tyr Thr Asp Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Asp His Asn Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 162
211 43 <211> 131 <223> Syntetisk Li04 variabel tungkjede-sekvens <400> 162 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr Asn Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Val Ile Tyr Pro Ser Gly Gly Gly Thr His Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Ser Ser Ile Ala Asp Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 163 <211> 131 <223> Syntetisk Li08 variabel tungkjede-sekvens <400> 163 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
212 44 Glu Met Val Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Arg Ser Ser Gly Gly Ala Thr Lys Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Glu Ser Pro Asp Asp Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 164 <211> 134 <223> Syntetisk Li11 variabel tungkjede-sekvens <400> 164 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Thr Ser Gly Gly Tyr Thr Gly Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
213 45 Ala Arg Asp Thr Ser Asp Asn Asp Tyr Tyr Tyr Met Asp Val Trp Gly Lys Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 165 <211> 133 <223> Syntetisk Li1O variabel tungkjede-sekvens <400> 165 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr Pro Met Val Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Trp Ile Gly Pro S~r Gly Gly Val Thr Ala Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Pro Tyr Ser Ser Gly Trp Trp Asp Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130
214 46 <210> 166 <211> 131 <223> Syntetisk Li01 variabel tungkjede-sekvens <400> 166 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Lys Tyr Gln Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Tyr Pro Ser Gly Gly Asn Thr Val Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Ser Gly Thr Thr Glu Ala Val Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 167 <211> 129 <223>Syntetisk Li07 variabel tungkjede-sekvens <400> 167 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Met Tyr
215 47 Phe Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Pro Ser Gly Gly Phe Thr Ser Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg His Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro <210> 168 <211> 132 <223> Syntetisk Li03 variabel tungkjede-sekvens <400> 168 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gln Tyr Pro Met Glu Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Gly Ile Tyr Pro Ser Gly Gly Ser Thr Val Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
216 48 Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ala Gly Gln Trp Leu Gly Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro 130 <210> 169 <211> 145 <223> Syntetisk Li12 variabel tungkjede-sekvens <400> 169 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gln Tyr Asn Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Arg Ile Ser Ser Ser Gly Gly Met Thr Met Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Ala Leu Arg Pro Tyr Cys Ser Gly Gly Ser Cys Tyr Ser Asp Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
217 49 Pro 145 <210> 170 <211> 116 <223> Syntetisk 1A7 variabel tungkjede-sekvens <400> 170 Gln Val Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Asp Thr Gly Glu Pro Thr Tyr Thr Glu Asp Phe Gln Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Val Tyr Leu Gln Phe Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Glu Gly Val His Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser 115 <210> 171 <211> 115 <223> Syntetisk 2F3 variabel tungkjede-sekvens <400> 171 Glu Val Lys Leu Glu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
218 50 Ser Met Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ala Trp Leu Asp Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val Ala Glu Ile Arg Ser Lys Ala Asn Asn His Ala Thr Aen Tyr Ala Glu Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Ile Tyr Phe Cys Thr Pro Ser Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser 115 <210> 172 <211> 117 <223> Syntetisk 3P1D10.2C3 og 3P1E11.3B7 variabel tungkjede-sekvens <400> 172 Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Leu Ser Cys Arg Ala Ser Gly Tyr Thr Phe Thr Ser Ser Trp Thr Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asp Gly Asp Thr Arg Tyr Thr Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
219 51 Ala Arg His Asn Ser Tyr Gly Met Asp Tyr Trp Gly Gln Gly Val Thr Val Ser Ser 115 <210> 173 <211> 399 <212> DNA <223> Syntetisk Li02 variabel tungkjede-sekvens <210> 174 <211> 435 <212> DNA <223> Syntetisk Li09 variabel tungkjede-sekvens <210> 175 <211> 393
220 52 <212> DNA <223> Syntetisk Li06 variabel tungkjede-sekvens <210> 176 <211> 393 <212> DNA <223> Syntetisk Li05 variabel tungkjede-sekvens <210> 177 <211> 393 <212> DNA <223> Syntetisk Li04 variabel tungkjede-sekvens
221 53 <210> 178 <211> 393 <212> DNA <223> Syntetisk Li08 variabel tungkjede-sekvens <210> 179 <211> 402 <212> DNA <223> Syntetisk Li11 variabel tungkjede-sekvens <210> 180 <211> 399 <212> DNA
222 54 <223> Syntetisk Li10 variabel tungkjede-sekvens < 210> 181 <211> 393 <212> DNA <223> Syntetisk Li01 variabel tungkjede-sekvens <210> 182 <211> 387 <212> DNA <223> Syntetisk Li07 variabel tungkjede-sekvens
223 55 <210> 183 <211> 396 <212> DNA <223> Syntetisk Li03 variabel tungkjede-sekvens <210> 184 <211> 435 <212> DNA <223> Syntetisk Li12 variabel tungkjede-sekvens <210> 185 <211> 357 <212> DNA
224 56 <223> Syntetisk Li02 variabel lettkjede-sekvens <210> 186 <211> 360 <212> DNA <223> Syntetisk Li09 variabel lettkjede-sekvens <210> 187 <211> 360 <212> DNA <223> Syntetisk Li06 variabel lettkjede-sekvens
225 57 <210> 188 <211> 363 <212> DNA <223> Syntetisk Li05 variabel lettkjede-sekvens <210> 189 <211> 360 <212> DNA <223> Syntetisk Li08 variabel lettkjede-sekvens <210> 190 <211> 360 <212> DNA <223> Syntetisk Li11 variabel lettkjede-sekvens
226 58 <210> 191 <211> 366 <212> DNA <223> Syntetisk Li10 variabel lettkjede-sekvens <210> 192 <211> 363 <212> DNA <223> Syntetisk Li01 variabel lettkjede-sekvens <210> 193 <211> 354 <212> DNA
227 59 <223> Syntetisk Li07 variabel lettkjede-sekvens <210> 194 <211> 360 <212> DNA <223> Syntetisk Li03 variabel lettkjede-sekvens <210> 195 <211> 10 <223> Syntetisk sekvens fra L1a.01-antistoff som koder for VH-CDR1 <400> 195 Gly Tyr Ser Phe Thr Asn Tyr Trp Ile Gly <210> 196 <211> 17 <223> Syntetisk sekvens fra L1a.01-antistoff som koder for VH-CDR2
228 60 <400> 196 Ile Ile Asp Pro Asp Asp Ser Tyr Thr Thr Tyr Ser Pro Ser Phe Gln Gly <210> 197 <211> 10 <223> Syntetisk sekvens fra L1a.01-antistoff som koder for VH-CDR3 <400> 197 Ala Glu Phe Tyr Trp Gly Ala Tyr Asp Gly <210> 198 <211> 10 <223> Syntetisk sekvens fra L1a.02-antistoff som koder for VH-CDR1 <400> 198 Gly Gly Ser Ile Arg Gly Asn Tyr Trp Ser <210> 199 <211> 14 <223> Syntetisk sekvens fra L1a.02-antistoff som koder for VH-CDR2 <400> 199 Ser Ile Asn Tyr Ser Gly Phe Thr Asn Pro Ser Leu Lys Gly <210> 200 <211> 8 <223> Syntetisk sekvens fra L1a.02-antistoff som koder for VH-CDR3 <400> 200
229 61 Val Arg His Trp Tyr Phe Asp Val 1 5 <210> 201 <211> 10 <223> Syntetisk sekvens fra L1a.03-antistoff som koder for VH-CDR1 <400> 201 Gly Tyr Thr Phe Asn Gly Phe Asp Met His <210> 202 <211> 17 <223> Syntetisk sekvens fra L1a.03-antistoff som koder for VH-CDR2 <400> 202 Trp Ile Asp Pro Tyr Asn Gly Ser Thr Thr Tyr Ala Gln Lys Phe Gln Gly <210> 203 <211> 13 <223> Syntetisk sekvens fra L1a.03-antistoff som koder for VH-CDR3 <400> 203 Asp Phe Tyr Met Asp Gly His Tyr Tyr Ile Phe Asp Val <210> 204 <211> 10 <223> Syntetisk sekvens fra L1a.04-antistoff som koder for VH-CDR1 <400> 204
230 62 Gly Tyr Ser Phe Ser Asn Tyr Tyr Ile His <210> 205 <211> 17 <223> Syntetisk sekvens fra L1a.04-antistoff som koder for VH-CDR2 <400> 205 Ile Ile Asp Pro Gly Asp Ser Phe Thr Ser Tyr Ser Pro Ser Phe Gln Gly <210> 206 <211> 11 <223> Syntetisk sekvens fra L1a.04-antistoff som koder for VH-CDR3 <400> 206 Asp Leu Ala Trp Ile Asp Tyr Gly Phe Asp Tyr <210> 207 <211> 10 <223> Syntetisk sekvens fra L1a.05-antistoff som koder for VH-CDR1 <400> 207 Gly Phe Thr Phe Thr Ser His Thr Val Ser <210> 208 <211> 17 <223> Syntetisk sekvens fra L1a.05-antistoff som koder for VH-CDR2 <400> 208 Ser Ile Thr Gly Asn Gly Ser Thr Thr Tyr Tyr Ala Asp Ser Val Lys
231 63 Gly <210> 209 <211> 7 <223> Syntetisk sekvens fra L1a.05-antistoff som koder for VH-CDR3 <400> 209 Phe Tyr Gly Asp Phe Asp Ser 1 5 <210> 210 <211> 10 <223> Syntetisk sekvens fra L1a.06-antistoff som koder for VH-CDR1 <400> 210 Gly Phe Thr Phe Ser Ser Asn Trp Met Ser <210> 211 <211> 17 <223> Syntetisk sekvens fra L1a.06-antistoff som koder for VH-CDR2 <400> 211 Thr Ile Phe Tyr Ser Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly <210> 212 <211> 17 <223> Syntetisk sekvens fra L1a.06-antistoff som koder for VH-CDR3
232 64 <400> 212 Asp Leu Pro Met Lys Gly Phe Ile Gln Gln Arg Tyr Gly Phe Asp Asp Val <210> 213 <211> 10 <223> Syntetisk sekvens fra L1a.07-antistoff som koder for VH-CDR1 <400> 213 Gly Phe Thr Phe Ser Gly Tyr Ala Ile Ser <210> 214 <211> 17 <223> Syntetisk sekvens fra L1a.07-antistoff som koder for VH-CDR2 <400> 214 Thr Ile Trp Gly Ser Gly Ser Thr Thr Tyr Tyr Ala Asp Ser Val Lys Gly <210> 215 <211> 11 <223> Syntetisk sekvens fra L1a.07-antistoff som koder for VH-CDR3 <400> ; 215 Glu Tyr Trp Tyr Tyr Asp Gln Phe Thr Ala Val <210> 216 <211> 12
233 65 <223> Syntetisk sekvens fra L1a.08-antistoff som koder for VH-CDR1 <400> 216 Gly Asp Ser Val Ser Ser Asn Ser Ala Ala Trp Ser <210> 217 <211> 18 <223> Syntetisk sekvens fra L1a.08-antistoff som koder for VH-CDR2 <400> 217 Arg Ile Tyr Tyr Arg Ser Lys Trp Tyr Asn Asp Tyr Ala Val Ser Val Lys Ser <210> 218 <211> 10 <223> Syntetisk sekvens fra L1a.08-antistoff som koder for VH-CDR3 <400> 218 Glu Val Tyr Ser Ala Gly Ile Met Asp Tyr <210> 219 <211> 10 <223> Syntetisk sekvens fra L1a.09-antistoff som koder for VH-CDR1 <400> 219 Gly Tyr Ser Phe Thr Asn His Trp Ile Gly <210> 220 <211> 17
234 66 <223> Syntetisk sekvens fra L1a.09-antistoff som koder for VH-CDR2 <400> 220 Ile Ile Asp Pro Ser Asp Ser Asp Thr Asn Tyr Ser Pro Ser Phe Gln Gly <210> 221 <211> 11 <223> Syntetisk sekvens fra L1a.09-antistoff som koder for VH-CDR3 <400> 221 Gly Phe Tyr Gly Ile Ala Asp Thr Phe Asp Val <210> 222 <211> 10 <223> Syntetisk sekvens fra L1a.10-antistoff som koder for VH-CDR1 <400> 222 Gly Tyr Ser Phe Thr Asn Tyr Trp Ile Ala <210> 223 <211> 17 <223> Syntetisk sekvens fra L1a.10-antistoff som koder for VH-CDR2 <400> 223 Met Ile Tyr Pro Asp Asp Ser Asn Thr Asn Tyr Ser Pro Ser Phe Gln Gly <210> 224 <211> 9
235 67 <223> Syntetisk sekvens fra L1a.10-antistoff som koder for VH-CDR3 <400> 224 Thr Asn Tyr Leu Gly Phe Tyr Asp Ser 1 5 <210> 225 <211> 10 <223> Syntetisk sekvens fra L1a.11-antistoff som koder for VH-CDR1 <400> 225 Gly Phe Thr Phe Ser Asp Tyr Gly Ile Ser <210> 226 <211> 17 <223> Syntetisk sekvens fra L1a.11-antistoff som koder for VH-CDR2 <400> 226 Asn Ile Leu Tyr Asp Gly Ser Glu Thr Tyr Tyr Ala Asp Ser Val Lys Gly <210> 227 <211> 11 <223> Syntetisk sekvens fra L1a.11-antistoff som koder for VH-CDR3 <400> 227 Gly Tyr Pro Thr Asp Asp Tyr Ser Phe Asp Ile <210> 228 <211> 12
236 68 <223> Syntetisk sekvens fra L1a.12-antistoff som koder for VH-CDR1 <400> 228 Gly Asp Ser Val Ser Asp Asn Ser Ala Ala Trp Gly <210> 229 <211> 18 <223> Syntetisk sekvens fra L1a.12-antistoff som koder for VH-CDR2 <400> 229 Arg Ile Tyr Tyr Arg Ser Lys Trp Tyr Asn Asp Tyr Ala Val Ser Val Lys Ser <210> 230 <211> 16 <223> Syntetisk sekvens fra L1a.12-antistoff som koder for VH-CDR3 <400> 230 Gly Arg His Glu Tyr Gly Gly Leu Gly Tyr Ala Glu Ala Met Asp His <210> 231 <211> 10 <223> Syntetisk sekvens fra L1a.13-antistoff som koder for VH-CDR1 <400> 231 Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser <210> 232 <211> 17
237 69 <223> Syntetisk sekvens fra L1a.13-antistoff som koder for VH-CDR2 <400> 232 Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly <210> 233 <211> 10 <223> Syntetisk sekvens fra L1a.13-antistoff som koder for VH-CDR3 <400> 233 His Tyr Thr Tyr Met His Phe Glu Asp Tyr <210> 234 <211> 11 <223> Syntetisk sekvens fra L1a.01-antistoff som koder for VL-CDR1 <400> 234 Ser Gly Asp Ser Leu Pro Ser Lys Phe Val His <210> 235 <211> 7 <223> Syntetisk sekvens fra L1a.01-antistoff som koder for VL-CDR2 <400> 235 Arg Asp Asn Asn Arg Pro Ser 1 5 <210> 236 <211> 8
238 70 <223> Syntetisk sekvens fra L1a.01-antistoff som koder for VL-CDR3 <400> 236 Ser Ser Tyr Asp Ala Leu Thr Asp 1 5 <210> 237 <211> 12 <223> Syntetisk sekvens fra L1a.02-antistoff som koder for VL-CDR1 <400> 237 Arg Ala Ser Gln Ser Ile Thr Asn Ser Tyr Leu Gly <210> 238 <211> 7 <223> Syntetisk sekvens fra L1a.02-antistoff som koder for VL-CDR2 <400> 238 Asp Ala Ser Ser Arg Ala Thr 1 5 <210> 239 <211> 8 <223> Syntetisk sekvens fra L1a.02-antistoff som koder for VL-CDR3 <400> 239 Gln Gln Ala Ser Asp Ala Pro Glu 1 5 <210> 240 <211> 11 <223> Syntetisk sekvens fra L1a.03-antistoff som koder for VL-CDR1 <400> 240
239 71 Arg Ala Ser Gln Gly Ile Asn Phe Trp Leu Asn <210> 241 <211> 7 <223> Syntetisk sekvens fra L1a.03-antistoff som koder for VL-CDR2 <400> 241 Ala Gly Ser Asn Leu Gln Ser 1 5 <210> 242 <211> 8 <223> Syntetisk sekvens fra L1a.03-antistoff som koder for VL-CDR3 <400> 242 Met Gln Asp Ser Asp Phe Pro Phe 1 5 <210> 243 <211> 14 <223> Syntetisk sekvens fra L1a.04-antistoff som koder for VL-CDR1 <400> 243 Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val Ser <210> 244 <211> 7 <223> Syntetisk sekvens fra L1a.04-antistoff som koder for VL-CDR2 <400> 244 Arg Asn Asn Asn Arg Pro Ser 1 5 <210> 245 <211> 8
240 72 <223> Syntetisk sekvens fra L1a.04-antistoff som koder for VL-CDR3 <400> 245 Gln Thr Tyr Asp Asn Ser Thr Asp 1 5 <210> 246 <211> 11 <223> Syntetisk sekvens fra L1a.05-antistoff som koder for VL-CDR1 <400> 246 Ser Gly Asp Asn Ile Arg Ser Tyr Tyr Val His <210> 247 <211> 7 <223> Syntetisk sekvens fra L1a.05-antistoff som koder for VL-CDR2 <400> 247 Glu Asp Ser Asn Arg Pro Ser 1 5 <210> 248 <211> 10 <223> Syntetisk sekvens fra L1a.05-antistoff som koder for VL-CDR3 <400> 248 Gln Ser Tyr Asp Ser Ala Ile Leu Leu His <210> 249 <211> 16 <223> Syntetisk sekvens fra L1a.06-antistoff som koder for VL-CDR1
241 73 <400> 249 Arg Ser Ser Gln Ser Leu Val Leu Arg Thr Gly Tyr Thr Tyr Leu Asn <210> 250 <211> 7 <223> Syntetisk sekvens fra L1a.06-antistoff som koder for VL-CDR2 <400> 250 Leu Val Ser Asn Arg Ala Ser 1 5 <210> 251 <211> 8 <223> Syntetisk sekvens fra L1a.06-antistoff som koder for VL-CDR3 <400> 251 Gln Gln Tyr Tyr Gly Met Pro Leu 1 5 <210> 252 <211> 12 <223> Syntetisk sekvens fra L1a.07-antistoff som koder for VL-CDR1 <400> 252 Arg Ala Ser Gln Ser Val Ser Tyr Gln Tyr Leu Ala <210> 253 <211> 7 <223> Syntetisk sekvens fra L1a.07-antistoff som koder for VL-CDR2 <400> 253 Gly Ala Ser Ser Arg Ala Thr 1 5
242 74 <210> 254 <211> 8 <223> Syntetisk sekvens fra L1a.07-antistoff som koder for VL-CDR3 <400> 254 Gln Gln Tyr Gly Ser Val Pro Arg 1 5 <210> 255 <211> 11 <223> Syntetisk sekvens fra L1a.08-antistoff som koder for VL-CDR1 <400> 255 Ser Gly Asp Ser Leu Gly Ser Tyr Tyr Val His <210> 256 <211> 7 <223> Syntetisk sekvens fra L1a.08-antistoff som koder for VL-CDR2 <400> 256 Asp Asp Asn Asp Arg Pro Ser 1 5 <210> 257 <211> 9 <223> Syntetisk sekvens fra L1a.08-antistoff som koder for VL-CDR3 <400> 257 Ser Ala Tyr Asp Tyr Ser Ala Arg Thr 1 5 <210> 258 <211> 11
243 75 <223> Syntetisk sekvens fra L1a.09-antistoff som koder for VL-CDR1 <400> 258 Ser Gly Asp Asn Leu Gly Ser Lys Tyr Val Ser <210> 259 <211> 7 <223> Syntetisk sekvens fra L1a.09-antistoff som koder for VL-CDR2 <400> 259 Asp Asp Asp Asp Arg Pro Ser 1 5 <210> 260 <211> 10 <223> Syntetisk sekvens fra L1a.09-antistoff som koder for VL-CDR3 <400> 260 Ser Ser Tyr Asp Phe Leu Asn Ile Gly Leu <210> 261 <211> 11 <223> Syntetisk sekvens fra L1a.10-antistoff som koder for VL-CDR1 <400> 261 Ser Gly Asp Ser Leu Gly Lys Lys Ser Val His <210> 262 <211> 7 <223> Syntetisk sekvens fra L1a.10-antistoff som koder for VL-CDR2 <400> 262 Glu Asp Ser Glu Arg Pro Ser
244 <210> 263 <211> 8 <223> Syntetisk sekvens fra L1a.10-antistoff som koder for VL-CDR3 <400> 263 Ser Ser Tyr Thr Asn Ser Val Asp 1 5 <210> 264 <211> 11 <223> Syntetisk sekvens fra L1a.11-antistoff som koder for VL-CDR1 <400> 264 Ser Gly Asp Asn Leu Gly Lys Lys Tyr Val Gly <210> 265 <211> 7 <223> Syntetisk sekvens fra L1a.11-antistoff som koder for VL-CDR2 <400> 265 Asp Asp Asp Asn Arg Pro Ser 1 5 <210> 266 <211> 8 <223> Syntetisk sekvens fra L1a.11-antistoff som koder for VL-CDR3 <400> 266 Gln Ser Tyr Asp Asp Thr Ser Ile 1 5
245 77 <210> 267 <211> 11 <223> Syntetisk sekvens fra L1a.12-antistoff som koder for VL-CDR1 <400> 267 Ser Gly Asp Ser Leu Gly Asn Lys Tyr Val His <210> 268 <211> 7 <223> Syntetisk sekvens fra L1a.12-antistoff som koder for VL-CDR2 <400> 268 Asp Asp Ser Asp Arg Pro Ser 1 5 <210> 269 <211> 8 <223> Syntetisk sekvens fra L1a.12-antistoff som koder for VL-CDR3 <400> 269 Gln Thr Trp Asp Tyr Val Gly Tyr 1 5 <210> 270 <211> 14 <223> Syntetisk sekvens fra L1a.13-antistoff som koder for VL-CDR1 <400> 270 Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr Asn Tyr Val Ser <210> 271 <211> 7
246 78 <223> Syntetisk sekvens fra L1a.13-antistoff som koder for VL-CDR2 <400> 271 Asp Val Ser Asn Arg Pro Ser 1 5 <210> 272 <211> 10 <223> Syntetisk sekvens fra L1a.13-antistoff som koder for VL-CDR3 <400> 272 Gln Ser Tyr Asp Arg Tyr Arg Leu Lys Asn <210> 273 <211> 119 <223> Syntetisk Li02 variabel lettkjede-sekvens <400> 273 Phe Tyr Ser His Ser Ala Gln Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Leu Gly Asp Lys Phe Ala Ser Trp Tyr Gln Gln Lys Ala Gly Gln Ser Pro Val Leu Val Ile Phe Gln Asp Arg Lys Arg Leu Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Thr Asn Thr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
247 79 Gly Gln Pro Lys Ala Ala Pro 115 <210> 274 <211> 120 <223> Syntetisk Li09 variabel lettkjede-sekvens <400> 274 Phe Tyr Ser His Ser Ala Gln Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Phe Val Gly Asp Arg Val Ala Ile Thr Cys Arg Ala Ser Gln Ser Ile Asp Thr Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Lys Leu Glu Asp Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Thr Gly Thr Asp Phe Thr Leu Thr Ile Arg Ser Leu Gln Pro Glu Asp Phe Gly Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Pro Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro <210> 275 <211> 120 <223> Syntetisk Li06 variabel lettkjede-sekvens <400> 275 Phe Tyr Ser His Ser Ala Gln Asp Ile Gln Met Thr Gln Ser Pro Ser
248 80 Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile Tyr Ala Ala Ser Ser Leu Arg Thr Gly Val Pro Ser Arg Phe Arg Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro <210> 276 <211> 121 <223> Syntetisk Li05 variabel lettkjede-sekvens <400> 276 Phe Tyr Ser His Ser Ala Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Ser Cys Gly Gly Asp Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Tyr Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asp Thr Ala Ile Leu Thr Ile Thr Arg Val Glu Val Gly Asp Glu Ala Asp Phe Tyr Cys Gln Val Arg
249 81 Asp Ser Arg Thr Glu Glu Arg Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu Gly Gln Pro Lys Ala Ala Pro <210> 277 <211> 120 <223> Syntetisk Li08 variabel lettkjede-sekvens <400> 277 Phe Tyr Ser His Ser Ala Gln Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Tyr Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile Tyr Asp Val Ser Asn Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Ala Ser Ala Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Ser Asp Asn Leu Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro <210> 278 <211> 120 <223> Syntetisk Li11 variabel lettkjede-sekvens <400> 278
250 82 Phe Tyr Ser His Ser Ala Gln Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Pro Ile Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Glu Ile Ala Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Thr Tyr Thr Leu Gln Thr Asp Val Pro Pro Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Thr Ala Thr Tyr Phe Cys Gln Gln Ala Asp Ile Phe Pro Leu Ser Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro <210> 279 <211> 122 <223> Syntetisk Li10 variabel lettkjede-sekvens <400> 279 Phe Tyr Ser His Ser Ala Gln Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Met Ser Ala Ser Val Gly Asp Thr Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Gly Asn Trp Leu Ala TrpTyr Gln Gln Lys Pro Gly Lys Ala Pro Thr Leu Leu Ile Tyr Ala Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Thr Gly Ser Gly Ser Ser Ser Gly Ile Asp Phe
251 83 Thr Leu Thr Ile Ser Asp Leu His Pro Glu Asp Leu Ala Thr Tyr Tyr Cys Gln Gln Ala Gln Thr Phe Pro Leu Thr Phe Gly Gly Gly Thr Arg Val Asp Leu Lys Arg Thr Val Ala Ala Pro <210> 280 <211> 121 <223> Syntetisk Li01 variabel lettkjede-sekvens <400> 280 Phe Tyr Ser His Ser Ala Gln Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asp Arg Phe Pro Ala Val Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro <210> 281 <211> 118
252 84 <223> Syntetisk Li07 variabel lettkjede-sekvens <400> 281 Phe Tyr Ser His Ser Ala Gln Ser Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln Thr Ala Ile Ile Thr Cys Ser Gly Asp Gln Leu Gly Asp Lys His Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Leu Asp Ile Lys Arg Pro Ala Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Arg Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ile Lys Thr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ser Gln Pro Lys Ala Ala Pro 115 <210> 282 <211> 120 <223> Syntetisk Li03 variabel lettkjede-sekvens <400> 282 Phe Tyr Ser His Ser Ala Gln Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser Leu Gln Ser Gly
253 85 Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro <210> 283 <211> 106 <223> Syntetisk 1A7 variabel lettkjede-sekvens <400> 283 Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys <210> 284 <211> 108
254 86 <223> Syntetisk 2F3 variabel lettkjede-sekvens <400> 284 Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Gly Asn Ile Tyr Asn Tyr Leu Ala Trp Phe Gln Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Val Tyr Asn Ala Lys Thr Leu Pro Asp Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Gln Tyr Phe Leu Lys Ile Asn Ser Leu Gln Pro Glu Asp Phe Gly Ser Tyr Tyr Cys Gln His Phe Trp Ala Ile Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg <210> 285 <211> 114 <223> Syntetisk 3P1D10.2C3 variabel lettkjede-sekvens <400> 285 Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
255 87 Ile Asn Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Arg <210> 286 <211> 114 <223> Syntetisk 3P1E11.3B7 variabel lettkjede-sekvens <400> 286 Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Ser Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Arg <210> 287 <211> 5
256 88 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (3).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <400> 287 Ile Thr Xaa Xaa Xaa 1 5 <210> 288 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (3).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin, eller asparagin <400> 288 Ala Cys Xaa Xaa Xaa 1 5 <210> 289 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (3).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <400> 289 Val Cys Xaa Xaa Xaa 1 5 <210> 290 <211> 5 <223> Syntetisk Sp35-peptidfragment
257 89 <221> DIVERSE_FUNKSJON <222> (3).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <400> 290 Ser Pro Xaa Xaa Xaa 1 5 <210> 291 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 291 Ser Pro Arg Lys His 1 5 <210> 292 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 292 Ser Pro Arg Lys Lys 1 5 <210> 293 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 293 Ser Pro Arg Lys Arg 1 5 <210> 294 <211> 5
258 90 <223> Syntetisk Sp35-peptidfragment <400> 294 Ser Pro Lys Lys His 1 5 <210> 295 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 295 Ser Pro His Lys His 1 5 <210> 296 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 296 Ser Pro Arg Arg His 1 5 <210> 297 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 297 Ser Pro Arg His His 1 5 <210> 298 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 298
259 91 Ser Pro Arg Arg Arg 1 5 <210> 299 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 299 Ser Pro His His His 1 5 <210> 300 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 300 Ser Pro Lys Lys Lys 1 5 <210> 301 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 301 Leu Ser Pro Arg Lys His 1 5 <210> 302 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 302 Leu Ser Pro Arg Lys Lys 1 5 <210> 303 <211> 6
260 92 <223> Syntetisk Sp35-peptidfragment <400> 303 Leu Ser Pro Arg Lys Arg 1 5 <210> 304 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 304 Leu Ser Pro Lys Lys His 1 5 <210> 305 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 305 Leu Ser Pro His Lys His 1 5 <210> 306 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 306 Leu Ser Pro Arg Arg His 1 5 <210> 307 <211> 6 <223> Syntetisk Sp35-peptidfragment
261 93 <400> 307 Leu Ser Pro Arg His His 1 5 <210> 308 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 308 Leu Ser Pro Arg Arg Arg 1 5 <210> 309 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 309 Leu Ser Pro His His His 1 5 <210> 310 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 310 Leu Ser Pro Lys Lys Lys 1 5 <210> 311 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 311 Trp Leu Ser Pro Arg Lys His 1 5
262 94 <210> 312 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 312 Trp Leu Ser Pro Arg Lys Lys 1 5 <210> 313 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 313 Trp Leu Ser Pro Arg Lys Arg 1 5 <210> 314 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 314 Trp Leu Ser Pro Lys Lys His 1 5 <210> 315 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 315 Trp Leu Ser Pro His Lys His 1 5 <210> 316 <211> 7
263 95 <223> Syntetisk Sp35-peptidfragment <400> 316 Trp Leu Ser Pro Arg Arg His 1 5 <210> 317 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 317 Trp Leu Ser Pro Arg His His 1 5 <210> 318 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 318 Trp Leu Ser Pro Arg Arg Arg 1 5 <210> 319 <211> 7 <223> Syntetisk Sp35-peptidfragment <400> 319 Trp Leu Ser Pro His His His 1 5 <210> 320 <211> 7 <223> Syntetisk Sp35-peptidfragment
264 96 <400> 320 Trp Leu Ser Pro Lys Lys Lys 1 5 <210> 321 <211> 6 <223> Syntetisk Sp35-peptidfragment <400> 321 Ile Thr Pro Lys Arg Arg 1 5 <210> 322 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 322 Ala Cys His His Lys 1 5 <210> 323 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 323 Val Cys His His Lys 1 5 <210> 324 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre
265 97 <400> 324 Xaa Xaa Arg Lys His 1 5 <210> 325 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 325 Xaa Xaa Arg Arg Arg 1 5 <210> 326 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 326 Xaa Xaa Lys Lys Lys 1 5 <210> 327 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre
266 98 <400> 327 Xaa Xaa His His His 1 5 <210> 328 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 328 Xaa Xaa Arg Lys Lys 1 5 <210> 329 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 329 Xaa Xaa Arg Lys Arg 1 5 <210> 330 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 330
267 99 Xaa Xaa Lys Lys His 1 5 <210> 331 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 331 Xaa Xaa His Lys His 1 5 <210> 332 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 332 Xaa Xaa Arg Arg His 1 5 <210> 333 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (1).. (2) <223> Xaa kan være hvilken som helst aminosyre <400> 333
268 100 Xaa Xaa Arg His His 1 5 <210> 334 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (3).. (3) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <221> DIVERSE_FUNKSJON <222> (4).. (4) <223> Xaa kan være hvilken som helst aminosyre <221> DIVERSE_FUNKSJON <222> (5).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <400> 334 Ile Thr Xaa Xaa Xaa 1 5 <210> 335 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (3).. (3) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <221> DIVERSE_FUNKSJON <222> (4).. (4) <223> Xaa kan være hvilken som helst aminosyre <221> DIVERSE_FUNKSJON <222> (5).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <400> 335
269 101 Ala Cys Xaa Xaa Xaa 1 5 <210> 336 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (3).. (3) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <221> DIVERSE_FUNKSJON <222> (4).. (4) <223> Xaa kan være hvilken som helst aminosyre <221> DIVERSE_FUNKSJON <222> (5).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <400> 336 Val Cys Xaa Xaa Xaa 1 5 <210> 337 <211> 5 <223> Syntetisk Sp35-peptidfragment <221> DIVERSE_FUNKSJON <222> (3).. (3) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <221> DIVERSE_FUNKSJON <222> (4).. (4) <223> Xaa kan være hvilken som helst aminosyre <221> DIVERSE_FUNKSJON <222> (5).. (5) <223> Xaa kan være lysin, argnin, histidin, glutamin eller asparagin <400> 337 Ser Pro Xaa Xaa Xaa 1 5
270 102 <210> 338 <211> 5 <223> Syntetisk Sp35-peptidfragment <400> 338 Ser Pro Arg Leu His 1 5 <210> 339 <211> 9 <223> Syntetisk Sp35-peptidfragment <400> 339 Arg Arg Ala Arg Ile Arg Asp Arg Lys 1 5 <210> 340 <211> 9 <223> Syntetisk Sp35-peptidfragment <400> 340 Lys Lys Val Lys Val Lys Glu Lys Arg 1 5 <210> 341 <211> 9 <223> Syntetisk Sp35-peptidfragment <400> 341 Arg Arg Leu Arg Leu Arg Asp Arg Lys 1 5 <210> 342 <211> 9
271 103 <223> Syntetisk Sp35-peptidfragment <400> 342 Arg Arg Gly Arg Gly Arg Asp Arg Lys 1 5 <210> 343 <211> 9 <223> Syntetisk Sp35-peptidfragment <400> 343 Arg Arg Ile Arg Ala Arg Asp Arg Lys 1 5 <210> 344 <211> 9 <223> Syntetisk fremover-primer brukt til å vise Sp35-uttrykk <400> 344 agagacatgc gattggtga 19 <210> 345 <211> 9 <223> Syntetisk revers-primer brukt til å vises Sp35-uttrykk <400> 345 agagatgtag acgaggtcat t 21 <210> 346 <211> 9 <223> Syntetisk Sp35 RNAi-nukleotid <400> 346 tgatcgtcat cctgctagac ttcaagagag tctagcagga tgacgatctt tttttc 55
272 104 <210> 347 <211> 9 <223> Syntetisk Sp35 RNAi-nukleotid <400> 347 tcgagaaaaa agatcgtcat cctgctagac tctcttgaag tctagcagga tgacgatca 59 <210> 348 <223> Syntetisk Sp35 RNAi-nukleotid <400> 348 tgatcctcat ccttctatac ttcaagagag tgtagcagga tgacgatctt ttttctcga 59 <210> 349 <211> 9 <223> Syntetisk Sp35 RNAi-nukleotid <400> 349 tcgagaaaaa agatcgtcat cctgctagac tctcttgaag tatagaagga tgacgatca 59 <210> 350 <211> 9 <223> Syntetisk primer brukt til å forsterke mus-sp35 med full lengde <400> 350 gaggatctcg acgcggccgc atggagacag acacactcct 9 41 <210> 351 <211> 9 <223> Syntetisk primer brukt til å forsterke mus-sp35 med full lengde <400> 351 ggggcggaat tggatcctca cagatcctct tctgagatga 9 41 <210> 352
273 105 <211> 41 <212> DNA <223> Syntetisk primer brukt til å forsterke dominant negativ Sp35 <400> 352 gaggatctcg acgcggccgc atggagacag acacactcct 9 41 <210> 353 <211> 42 <212> DNA <223> Syntetisk primer brukt til å forsterke dominant negativ Sp35 <400> 353 gatacggatc ctcagccttt gccccggctc catagaaaca gc 42 <210> 354 <211> 37 <212> DNA <223> Syntetisk primer brukt til å skaffe delvis kodesekvens for humant Sp35 Sp35 <400> 354 cagcaggtcg acgcggccgc atgctggcgg ggggcgt 37 <210> 355 <211> 59 <212> DNA <223> Syntetisk primer brukt til å skaffe delvis kodesekvens for humant Sp35 <400> 355 cagcaggtcg acctcgcccg gctggttggc caaccagccg ggcgaggtcg acctcgagg 59 <210> 356 <211> 39 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til lettkjedet til P1E11.3B7 <400> 356 ggggatatcc accatggatt ttcaggtgca gattttcag 39
274 106 <210> 357 <211> 40 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til lettkjedet til P1E11.3B7 <400> 357 ggggatatcc accatgragt cacakacyca ggtcttyrta 40 <210> 358 <211> 37 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til lettkjedet til P1E11.3B7 <400> 358 ggggatatcc accatgaagt tgcctgttag gctgttg 37 <210> 359 <211> 40 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til lettkjedet til P1E11.3B7 <400> 359 ggggatatcc accatgaggk ccccwgctca gytyctkgga 40 <210> 360 <211> 39 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til tungkjedet til P1E11.3B7 <400> 360 ggggatatcc accatggrat gsagctgkgt matsctctt 39 <210> 361 <211> 39 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til tungkjedet til P1E11.3B7
275 107 <400> 361 ggggatatcc accatgract tcgggytgag ctkggtttt 39 <210> 362 <211> 38 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til tungkjedet til P1E11.3B7 <400> 362 ggggatatcc accatggctg tcttggggct gctcttct 38 <210> 363 <211> 39 <212> DNA <223> Syntetisk primer brukt til å bestemme sekvensen til tungkjedet til P1E11.3B7 <400> 363 aggtctagaa yctccacaca caggrrccag tggatagac 39 <210> 364 <211> 22 <212> DNA <223> Syntetisk tungkjede universal primer <400> 364 aggtsmarct gcagsagtcw gg 22 <210> 365 <211> 34 <212> DNA <223> Syntetisk tungkjede universal primer <400> 365 tgaggagacg gtgaccgtgg tcccttggcc ccag 34 <210> 366 <211> 28 <212> DNA
276 108 <223> Syntetisk degenerert signalsekvensprimer <221> diverse_funksjon <222> (21).. (21) <223> n kan være et hvilket som helst nykleotid <221> diverse_funksjon <222> (24).. (24) <223> n kan være et hvilket som helst nykleotid <400> 366 atggartgya aytggathct nccnttya 28 <210> 367 <211> 39 <212> DNA <223> Syntetisk konstant domain primer <400> 367 aggtctagaa yctccacaca caggrrccag tggatagac 39 <210> 368 <211> 66 <212> DNA <223> Syntetisk primer som brukes til å forsterke rotte-mog <400> 368 ggggtatctc tcgagaaaag agagcatcat catcatcatc atatgggaca gttcagagtg 60 ataggg 66 <210> 369 <211> 40 <212> DNA <223> Syntetisk primer som brukes til å forsterke rotte-mog <400> 369 ttcgcggccg ctattagcca gggttgatcc agtagaaggg 40 <210> 370 <211> 354 <212> DNA
277 109 <223> Syntetisk Li13 variabel tungkjede-sekvens <210> 371 <211> 324 <212> DNA <223> Syntetisk Li13 variabel lettkjede-sekvens <210> 372 <211> 118 <223> Syntetisk Li13 variabel tungkjede-sekvens <400> 372 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr Glu Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Arg Ile Val Ser Ser Gly Gly Phe Thr Lys Tyr Ala Asp Ser Val
278 110 Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Thr Glu Gly Asp Asn Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser 115 <210> 373 <211> 108 <223> Syntetisk Li13 variabel lettkjede-sekvens <400> 373 Asp Ile Gln Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Met Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile LYs <210> 374 <211> 354 <212> DNA
279 111 <223> Syntetisk Li32 variabel tungkjede-sekvens <210> 375 <211> 321 <212> DNA <223> Syntetisk Li32 variabel lettkjede-sekvens <210> 376 <211> 118 <223> Syntetisk Li32 variabel tungkjede-sekvens <400> 376 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ala Tyr Met Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
280 112 Ser Ser Ile Ser Pro Ser Gly Gly Asn Thr Lys Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Asp Tyr Gly Tyr Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser 115 <210> 377 <211> 107 <223> Syntetisk Li32 variabel lettkjede-sekvens <400> 377 Asp Ile Gln Met Thr Gln Ser Pro Asp Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Tyr Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Met Ala Pro Lys Leu Leu Ile Tyr Asp Ala Phe Ile Leu Glu Gly Gly Ala Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Phe Thr Ile Ser Asn Leu Gln Pro Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Ser Asp Gln Leu Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Arg <210> 378 <211> 357
281 113 <212> DNA <223> Syntetisk Li33 variabel tungkjede-sekvens <210> 379 <211> 321 <212> DNA <223> Syntetisk Li33 variabel lettkjede-sekvens <210> 380 <211> 119 <223> Syntetisk Li33 variabel tungkjede-sekvens <400> 380 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ile Tyr Pro Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
282 114 Ser Trp Ile Gly Pro Ser Gly Gly Ile Thr Lys Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys Ala Arg Glu Gly His Asn Asp Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser 115 <210> 381 <211> 107 <223> Syntetisk Li33 variabel lettkjede-sekvens <400> 381 Asp Ile Gln Met Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asp Lys Trp Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
283 115 <210> 382 <211> 357 <212> DNA <223> Syntetisk Li34 variabel tungkjede-sekvens <210> 383 <211> 321 <212> DNA <223> Syntetisk Li34 variabel lettkjede-sekvens <210> 384 <211> 119 <223> Syntetisk Li34 variabel tungkjede-sekvens <400> 384 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
284 116 Glu Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Gly Ile Tyr Ser Ser Gly Gly Ile Thr Val Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr O Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ala Ala Ile Leu Asp Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser 115 <210> 385 <211> 107 <223> Syntetisk Li34 variabel lettkjede-sekvens <400> 385 Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys His Ala Ser Gln Asp Ile Ser Asn Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Phe Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro O Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Asp Asn Leu Pro Phe Thr Phe Gly Pro Gly Thr Arg Val Ala Ile Arg
285 117 <210> 386 <211> 11 <223> Syntetisk sekvens fra Li13-antistoff som koder for VL-CDR1 <400> 386 Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala <210> 387 <211> 7 <223> Syntetisk sekvens fra Li13-antistoff som koder for VL-CDR2 <400> 387 Asp Ala Ser Asn Arg Ala Thr 1 5 <210> 388 <211> 10 <223> Syntetisk sekvens fra Li13-antistoff som koder for VL-CDR3 <400> 388 Gln Gln Arg Ser Asn Trp Pro Met Tyr Thr <210> 389 <211> 5 <223> Syntetisk sekvens fra Li13-antistoff som koder for VH-CDR1 <400> 389 His Tyr Glu Met Tyr 1 5 <210> 390 <211> 17
286 118 <223> Syntetisk sekvens fra Li13-antistoff som koder for VH-CDR2 <400> 390 Arg Ile Val Ser Ser Gly Gly Phe Thr Lys Tyr Ala Asp Ser Val Lys Gly <210> 391 <211> 9 <223> Syntetisk sekvens fra Li13-antistoff som koder for VH-CDR3 <400> 391 Glu Gly Asp Asn Asp Ala Phe Asp Ile 1 5 <210> 392 <211> 11 <223> Syntetisk sekvens fra Li32-antistoff som koder for VL-CDR1 <400> 392 Gln Ala Ser Gln Asp Ile Ser Tyr Tyr Leu Asn <210> 393 <211> 7 <223> Syntetisk sekvens fra Li32-antistoff som koder for VL-CDR2 <400> 393 Asp Ala Phe Ile Leu Glu Gly 1 5 <210> 394 <211> 9
287 119 <223> Syntetisk sekvens fra Li32-antistoff som koder for VL-CDR3 <400> 394 Gln Gln Ser Asp Gln Leu Pro Val Thr 1 5 <210> 395 <211> 5 <223> Syntetisk sekvens fra Li32-antistoff som koder for VH-CDR1 <400> 395 Ala Tyr Met Met Gln 1 5 <210> 396 <211> 17 <223> Syntetisk sekvens fra Li32-antistoff som koder for VH-CDR2 <400> 396 Ser Ile Ser Pro Ser Gly Gly Asn Thr Lys Tyr Ala Asp Ser Val Lys Gly <210> 397 <211> 9 <223> Syntetisk sekvens fra Li32-antistoff som koder for VH-CDR3 <400> 397 Gly Asp Tyr Gly Tyr Trp Phe Asp Pro 1 5 <210> 398 <211> 11
288 120 <223> Syntetisk sekvens fra Li33-antistoff som koder for VL-CDR1 <400> 398 Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala <210> 399 <211> 7 <223> Syntetisk sekvens fra Li33-antistoff som koder for VL-CDR2 <400> 399 Asp Ala Ser Asn Arg Ala Thr 1 5 <210> 400 <211> 9 <223> Syntetisk sekvens fra Li33-antistoff som koder for VL-CDR3 <400> 400 Gln Gln Tyr Asp Lys Trp Pro Leu Thr 1 5 <210> 401 <211> 5 <223> Syntetisk sekvens fra Li33-antistoff som koder for VH-CDR1 <400> 401 Ile Tyr Pro Met Phe 1 5 <210> 402 <211> 17 <223> Syntetisk sekvens fra Li33-antistoff som koder for VH-CDR2 <400> 402
289 121 Trp Ile Gly Pro Ser Gly Gly Ile Thr Lys Tyr Ala Asp Ser Val Lys Gly <210> 403 <211> 10 <223> Syntetisk sekvens fra Li33-antistoff som koder for VH-CDR3 <400> 403 Glu Gly His Asn Asp Trp Tyr Phe Asp Leu <210> 404 <211> 11 <223> Syntetisk sekvens fra Li34-antistoff som koder for VL-CDR1 <400> 404 His Ala Ser Gln Asp Ile Ser Asn Tyr Leu Ser <210> 405 <211> 7 <223> Syntetisk sekvens fra Li34-antistoff som koder for VL-CDR2 <400> 405 Asp Ala Phe Asn Leu Glu Thr 1 5 <210> 406 <211> 9 <223> Syntetisk sekvens fra Li34-antistoff som koder for VL-CDR3 <400> 406 Gln His Tyr Asp Asn Leu Pro Phe Thr 1 5
290 122 <210> 407 <211> 5 <223> Syntetisk sekvens fra Li34-antistoff som koder for VH-CDR1 <400> 407 Asn Tyr Glu Met Tyr 1 5 <210> 408 <211> 17 <223> Syntetisk sekvens fra Li34-antistoff som koder for VH-CDR2 <400> 408 Gly Ile Tyr Ser Ser Gly Gly Ile Thr Val Tyr Ala Asp Ser Val Lys Gly <210> 409 <211> 10 <223> Syntetisk sekvens fra Li34-antistoff som koder for VH-CDR3 <400> 409 Ala Ala Ile Leu Asp Trp Tyr Phe Asp Leu <210> 410 <211> 5 <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VH-CDR1 <400> 410 Ser Tyr Trp Met His 1 5
291 123 <210> 411 <211> 17 <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VH-CDR2 <400> 411 Val Ile Asp Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Arg Gly <210> 412 <211> 11 <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VH-CDR3 <400> 412 Pro Tyr Tyr Gly Ser His Trp Phe Phe Asp Val <210> 413 <211> 10 <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VL-CDR1 <400> 413 Ser Ala Ser Ser Arg Val Ser Tyr Val His <210> 414 <211> 7 <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VL-CDR2 <400> 414 Asp Thr Ser Asn Leu Ala Ser 1 5
292 124 <210> 415 <211> 9 <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VL-CDR3 <400> 415 Gln Gln Trp Ser Thr Asn Pro Pro Thr 1 5 <210> 416 <211> 120 <223> Syntetisk 3B5.2 variabel tungkjede-sekvens <400> 416 Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Thr Ser Val Lys Leu Ser Cys Arg Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Val Ile Asp Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Arg Gly Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Pro Tyr Tyr Gly Ser His Trp Phe Phe Asp Val Trp Gly Thr Gly Thr Thr Val Thr Val Ser Ser <210> 417 <211> 106
293 125 <223> Syntetisk 3B5.2 variabel lettkjede-sekvens <400> 417 Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Arg Val Ser Tyr Val His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Leu Tyr Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Gly Gly Asn Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Thr Asn Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys <210> 418 <211> 6 <223> Syntetisk consensus human and murine kappa lettkjede-sekvens <400> 418 Ser Gly Ser Gly Ser Gly 1 5 <210> 419 <211> 106 <223> Syntetisk mutant 3B5.2 variabel lettkjede-sekvens <400> 419 Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
294 126 Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Arg Val Ser Tyr Val His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Leu Tyr Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Thr Asn Pro Pro Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys <210> 420 <211> 1404 <212> DNA <223> Syntetisk murin og human kimærisk antistoff-3b5.2-sekvens
295 127 <210> 421 <211> 708 <212> DNA <223> Syntetisk murin og human kimærisk antistoff-3b5.2-sekvens <210> 422 <211> 360 <212> DNA
296 128 <223> Syntetisk 3B5.2 variabel tungkjede-sekvens <210> 423 <211> 318 <212> DNA <223> Syntetisk 3B5.2 variabel lettkjede-sekvens <210> 424 <211> 15 <212> DNA <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VH-CDR1 <400> 424 agctactgga tgcac 15 <210> 425 <211> 51 <212> DNA <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VH-CDR2 <400> 425 gtgattgatc cttctgatag ttatactaac tacaatcaaa agttcagggg c 51
297 129 <210> 426 <211> 33 <212> DNA <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VH-CDR3 <400> 426 ccttactacg gtagtcactg gttcttcgat gtc 33 <210> 427 <211> 30 <212> DNA <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VL-CDR1 <400> 427 agtgccagct cacgtgtaag ttacgtgcac 30 <210> 428 <211> 21 <212> DNA <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VL-CDR2 <400> 428 gacacatcca acctggcttc t 21 <210> 429 <211> 27 <212> DNA <223> Syntetisk sekvens fra 3B5.2-antistoff som koder for VL-CDR3 <400> 429 cagcagtgga gtactaaccc acccacg 27 <210> 430 <211> 106 <223> Syntetisk lettkjede variant 1 <400> 430 Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
298 130 Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Leu Ile Tyr Asp Thr Ser Lys L~u Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys <210> 431 <211> 106 <223> Syntetisk lettkjede variant 2 <400> 431 Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr /J68
299 131 Phe Gly Gln Gly Thr Lys Val Glu Ile Lys <210> 432 <211> 116 <223> Syntetisk tungkjede variant 2 <400> 432 Gln Val Gln Leu Val Gln Ser Gly His Glu Val Lys Gln Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Asp Thr Gly Glu Pro Thr Tyr Thr Glu Asp Phe Gln Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Ala Ser Thr Val Tyr Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Met Ala Met Tyr Tyr Cys Ala Arg Glu Gly Val His Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser 115 <210> 433 <211> 447 <223> Syntetisk Li81 variabel tungkjede-sekvens <400> 433 Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ala Tyr
300 132 NO/EP
301 133 <210> 434 <211> 215 <223> Syntetisk Li81 variabel lettkjede-sekvens <400> 434 Asp Ile Gln Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
302 134 <210> 435 <211> 447 <223> Syntetisk Li81 aglycosylated variabel tungkjede-sekvens <400> 435
303 135 NO/EP
304 136 <210> 436 <211> 5 <223> Syntetisk sekvens fra Li81-antistoff som koder for VH-CDR1 <400> 436
305 137 Ala Tyr Glu Met Lys 1 5 <210> 437 <211> 17 <223> Syntetisk sekvens fra Li81-antistoff som koder for VH-CDR2 <400> 437 Val Ile Gly Pro Ser Gly Gly Phe Thr Phe Tyr Ala Asp Ser Val Lys Gly <210> 438 <211> 9 <223> Syntetisk sekvens fra Li81-antistoff som koder for VH-CDR3 <400> 438 Glu Gly Asp Asn Asp Ala Phe Asp Ile 1 5 <210> 439 <211> 15 <212> DNA <223> Syntetisk sekvens fra Li81-antistoff som koder for VH-CDR1 <400> 439 gcttacgaga tgaag 15 <210> 440 <211> 51 <212> DNA <223> Syntetisk sekvens fra Li81-antistoff som koder for VH-CDR2 <400> 440 gttatcggtc cttctggtgg ctttactttt tatgctgact ccgttaaagg t 51 <210> 441
306 138 <211> 27 <212> DNA <223> Syntetisk sekvens fra Li81-antistoff som koder for VH-CDR3 <400> 441 gagggtgata atgatgcttt tgatatc 27 <210> 442 <211> 11 <223> Syntetisk sekvens fra Li81-antistoff som koder for VL-CDR1 <400> 442 Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala <210> 443 <211> 7 <223> Syntetisk sekvens fra Li81-antistoff som koder for VL-CDR2 <400> 443 Asp Ala Ser Asn Arg Ala Thr 1 5 <210> 444 <211> 10 <223> Syntetisk sekvens fra Li81-antistoff som koder for VL-CDR3 <400> 444 Gln Gln Arg Ser Asn Trp Pro Met Tyr Thr <210> 445 <211> 33 <212> DNA <223> Syntetisk sekvens fra Li81-antistoff som koder for VL-CDR1
307 139 <400> 445 agggccagtc agagtgttag cagctactta gcc 33 <210> 446 <211> 21 <212> DNA <223> Syntetisk sekvens fra Li81-antistoff som koder for VL-CDR2 <400> 446 gatgcatcca acagggccac t 21 <210> 447 <211> 30 <212> DNA <223> Syntetisk sekvens fra Li81-antistoff som koder for VL-CDR3 <400> 447 cagcagcgta gcaactggcc gatgtacact 30 <210> 448 <211> 1341 <212> DNA <223> Syntetisk Li81 variabel tungkjede-sekvens
308 140 <210> 449 <211> 648 <212> DNA <223> Syntetisk Li81 variabel lettkjede-sekvens <210> 450 <211> 1341 <212> DNA
309 141 <223> Syntetisk Li81 aglycosylated variabel tungkjede-sekvens <210> 451 <211> 94 <223> syntetisk murin undergruppe kappa 6 variabel lettkjede-sekvens <400> 451 Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
310 142 <210> 452 <211> 96 <223> Syntetisk murin undergruppe HVMS variabel tungkjede-sekvens <210> 453 <211> 96
311 143 <223> Syntetisk murin konsensussekvens <221> DIVERSE_FUNKSJON <222> (2).. (2) <223> Xaa er enten isoleucin eller valin <221> DIVERSE_FUNKSJON <222> (54).. (54) <223> Xaa er enten aspartat eller glutamat <221> DIVERSE_FUNKSJON <222> (61).. (61) <223> Xaa er enten treonin eller alanin <221> DIVERSE_FUNKSJON <222> (62).. (62) <223> Xaa er enten glutamat eller aspartat <221> DIVERSE_FUNKSJON <222> (65).. (65) <223> Xaa er enten glutamin eller lysin <221> DIVERSE_FUNKSJON <222> (79).. (79) <223> Xaa er enten valin eller alanin <221> DIVERSE_FUNKSJON <222> (83).. (83) <223> Xaa er enten fenylalanin eller isoleucin
312 144 Leu Gln Xaa Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys <210> 454 <211> 96 <223> Murin VGK6-kimlinje <210> 455 <211> 94 <223> Syntetisk human subgruppe kappa 3 variabel lettkjede-sekvens
313 145 <210> 456 <211> 95 <223> Syntetisk konsensussekvens <221> DIVERSE_FUNKSJON <222> (1).. (1) <223> Xaa kan være enten glutamin eller glutamat <221> DIVERSE_FUNKSJON <222> (10).. (10) <223> Xaa kan være enten isoleucin eller treonin <221> DIVERSE_FUNKSJON <222> (11).. (11) <223> Xaa kan være enten metionin eller leucin <221> DIVERSE_FUNKSJON <222> (13).. (13) <223> Xaa kan være enten alanin eller leucin <221> DIVERSE_FUNKSJON <222> (18).. (18) <223> Xaa kan være enten lysin eller argnin <221> DIVERSE_FUNKSJON <222> (19).. (19) <223> Xaa kan være enten valin eller alanin <221> DIVERSE_FUNKSJON <222> (21).. (21) <223> Xaa kan være enten metionin eller leucin <221> DIVERSE_FUNKSJON <222> (22).. (22) <223> Xaa kan være enten treonin eller serin
314 146 <221> DIVERSE_FUNKSJON <222> (24).. (24) <223> Xaa kan være enten serin eller arginin <221> DIVERSE_FUNKSJON <222> (27).. (27) <223> Xaa kan være enten serin eller glutamin <221> DIVERSE_FUNKSJON <222> (31).. (31) <223> Xaa kan være enten serin eller ikke finnes <221> DIVERSE_FUNKSJON <222> (33).. (33) <223> Xaa kan være enten metionin eller leucin <221> DIVERSE_FUNKSJON <222> (34).. (34) <223> Xaa kan være enten histidin eller alanin <221> DIVERSE_FUNKSJON <222> (40).. (40) <223> Xaa kan være enten serin eller prolin <221> DIVERSE_FUNKSJON <222> (42).. (42) <223> Xaa kan være enten treonin eller glutamin <221> DIVERSE_FUNKSJON <222> (43).. (43) <223> Xaa kan være enten serin eller alanin <221> DIVERSE_FUNKSJON <222> (45).. (45) <223> Xaa kan være enten lysin eller arginin <221> DIVERSE_FUNKSJON <222> (46).. (46) <223> Xaa kan være enten arginin eller leucin <221> DIVERSE_FUNKSJON <222> (47).. (47) <223> Xaa kan være enten tryptofan eller leucin
315 147 <221> DIVERSE_FUNKSJON <222> (51).. (51) <223> Xaa kan være enten treonin eller alanin <221> DIVERSE_FUNKSJON <222> (53).. (53) <223> Xaa kan være enten lysin eller asparagin <221> DIVERSE_FUNKSJON <222> (54).. (54) <223> Xaa kan være enten leucin eller arginin <221> DIVERSE_FUNKSJON <222> (56).. (56) <223> Xaa kan være enten serin eller treonin <221> DIVERSE_FUNKSJON <222> (58).. (58) <223> Xaa kan være enten valin eller isoleucin <221> DIVERSE_FUNKSJON <222> (70).. (70) <223> Xaa kan være enten serin eller aspartat <221> DIVERSE_FUNKSJON <222> (71).. (71) <223> Xaa kan være enten tyrosin eller fenylalanin <221> DIVERSE_FUNKSJON <222> (72).. (72) <223> Xaa kan være enten serin eller treonin <221> DIVERSE_FUNKSJON <222> (78).. (78) <223> Xaa kan være enten metionin eller leucin <221> DIVERSE_FUNKSJON <222> (80).. (80) <223> Xaa kan være enten alanin eller prolin <221> DIVERSE_FUNKSJON <222> (85).. (85) <223> Xaa kan være enten treonin eller valin
316 148 <221> DIVERSE_FUNKSJON <222> (91).. (91) <223> Xaa kan være enten tryptofan eller arginin <221> DIVERSE_FUNKSJON <222> (93).. (93) <223> Xaa kan være enten serin eller asparagin <221> DIVERSE_FUNKSJON <222> (94).. (94) <223> Xaa kan være enten asparagin eller tryptofan <210> 457 <211> 95 <223> Human L6-kimlinje
317 149 <210> 458 <211> 98 <223> Syntetisk human undergruppe MHV1 variabel tungkjede-sekvens Ala Arg
318 150 <210> 459 <211> 98 <223> Syntetisk konsensussekvens <221> DIVERSE_FUNKSJON <222> (11).. (11) <223> Xaa kan være enten leucin eller valin <221> DIVERSE_FUNKSJON <222> (13).. (13) <223> Xaa kan være enten lysin eller glutamin <221> DIVERSE_FUNKSJON <222> (16).. (16) <223> Xaa kan være enten glutamat eller alanin <221> DIVERSE_FUNKSJON <222> (17).. (17) <223> Xaa kan være enten treonin eller serin <221> DIVERSE_FUNKSJON <222> (20).. (20) <223> Xaa kan være enten isoleucin eller valin <221> DIVERSE_FUNKSJON <222> (28).. (28) <223> Xaa kan være enten treonin eller serin <221> DIVERSE_FUNKSJON <222> (31).. (31) <223> Xaa kan være enten argnin eller treonin <221> DIVERSE_FUNKSJON <222> (38).. (38) <223> Xaa kan være lysin eller prolin <221> DIVERSE_FUNKSJON <222> (43).. (43) <223> Xaa kan være enten lysin eller glutamin <221> DIVERSE_FUNKSJON <222> (46).. (46) <223> Xaa kan være enten lysin eller glutamat
319 151 <221> DIVERSE_FUNKSJON <222> (51).. (51) <223> Xaa kan være enten isoleucin eller fenylalanin <221> DIVERSE_FUNKSJON <222> (54).. (54) <223> Xaa kan være enten aspartat eller tyrosine <221> DIVERSE_FUNKSJON <222> (57).. (57) <223> Xaa kan være enten glutamat eller arginin <221> DIVERSE_FUNKSJON <222> (61).. (61) <223> Xaa kan være enten treonin eller alanin <221> DIVERSE_FUNKSJON <222> (62).. (62) <223> Xaa kan være enten glutamat eller glutamin <221> DIVERSE_FUNKSJON <222> (63).. (63) <223> Xaa kan være enten aspartat eller glycin <221> DIVERSE_FUNKSJON <222> (65).. (65) <223> Xaa kan være enten glutamin eller treonin <221> DIVERSE_FUNKSJON <222> (69).. (69) <223> Xaa kan være enten alanin eller valin <221> DIVERSE_FUNKSJON <222> (72).. (72) <223> Xaa kan være enten leucin eller metionin <221> DIVERSE_FUNKSJON <222> (73).. (73) <223> Xaa kan være enten glutamat eller aspartat <221> DIVERSE_FUNKSJON <222> (79).. (79) <223> Xaa kan være enten valin eller alanin
320 152 <221> DIVERSE_FUNKSJON <222> (83).. (83) <223> Xaa kan være enten fenylalanin eller isoleucin <221> DIVERSE_FUNKSJON <222> (84).. (84) <223> Xaa kan være enten argnin eller serin <221> DIVERSE_FUNKSJON <222> (85).. (85) <223> Xaa kan være enten argnin eller serin <221> DIVERSE_FUNKSJON <222> (88).. (88) <223> Xaa kan være enten argnin eller alanin <221> DIVERSE_FUNKSJON <222> (91).. (91) <223> Xaa kan være enten treonin eller metionin <221> DIVERSE_FUNKSJON <222> (93).. (93) <223> Xaa kan være enten treonin eller metionin <221> DIVERSE_FUNKSJON <222> (95).. (95) <223> Xaa kan være enten fenylalanin eller tyrosin
321 153 Ala Arg <210> 460 <211> 98 <223> Human huvh7-81-kimlinje
322 154 Ala Arg <210> 461 <211> 1395 <212> DNA <223> Human hup1a7-igg1 H2 tungkjede-sekvens
323 155 <210> 462 <211> 464 <223> Human hup1a7 H2 tungkjede-sekvens
324 156 NO/EP
325 157 <210> 463 <211> 708 <212> DNA <223> Human hup1a7 L1 kappa lettkjede-sekvens <210> 464 <211> 235
326 158 <223> Human hup1a7 L1 lettkjede-sekvens
327 159 Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys <210> 465 <211> 708 <212> DNA <223> Human hup1a7 L2 kappa lettkjede-sekvens <210> 466 <211> 235 <223> Human hup1a7 L2-lettkjede-sekvens
328 160 NO/EP
329 161 <210> 467 <211> 708 <212> DNA <223> Murin og human kimærisk chp1a7 kappa lettkjede-sekvens
330 162 <210> 468 <211> 235 <223> Murin og human kimærisk chp1a7-lettkjede-sekvens
331 163 NO/EP
332 164 <210> 469 <211> 1395 <212> DNA <223> Murin og human kimærisk chp1a7 huigg1-tungkjede-sekvens
333 165 <210> 470 <211> 464 <223> Murin og human kimærisk chp1a7-tungkjede-sekvens
334 166 NO/EP
335 167 <210> 471 <211> 106 <223> Syntetisk lettkjede variant 3
336 168 <210> 472 <211> 116 <223> Syntetisk tungkjede variant 1 <210> 473 <211> 116 <223> Syntetisk tungkjede variant 3 <400> 473
337 169 <210> 474 <211> 466 <223> Syntetisk Li81 aglykosylert variabel tungkjede-sekvens
338 170 NO/EP
339 171 NO/EP
340 1 ERSTATNINGSARK (REGEL 26)
341 2 ERSTATNINGSARK (REGEL 26)
342 3 ERSTATNINGSARK (REGEL 26)
343 4 ERSTATNINGSARK (REGEL 26)
344 5 ERSTATNINGSARK (REGEL 26)
345 6 ERSTATNINGSARK (REGEL 26)
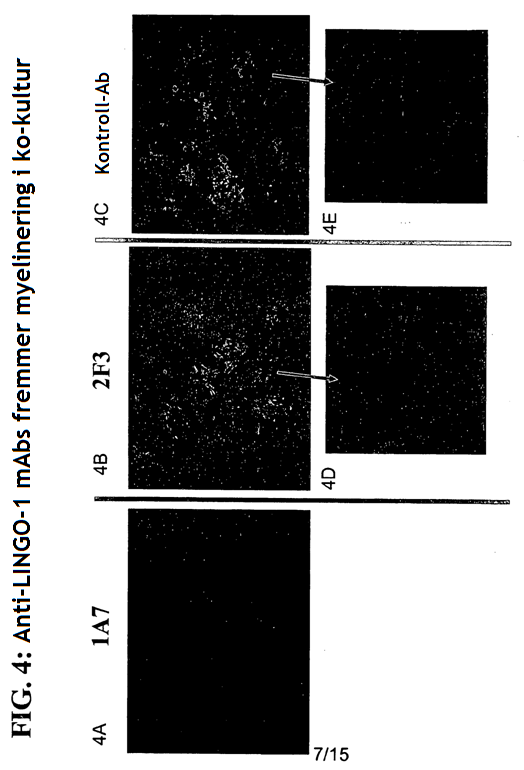
346 7 ERSTATNINGSARK (REGEL 26)
347 fort. NO/EP ERSTATNINGSARK (REGEL 26)
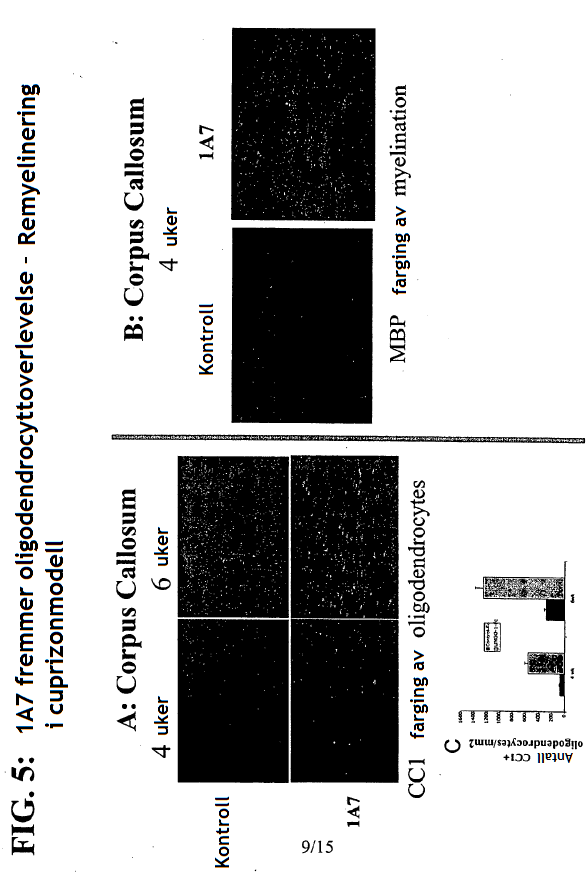
348 9 ERSTATNINGSARK (REGEL 26)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2310382 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/4412 (2006.01) A61P 35/00 (2006.01) C07D 401/14 (2006.01) C07D 403/12 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2310382 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/4412 (2006.01) A61P 35/00 (2006.01) C07D 401/14 (2006.01) C07D 403/12 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2148670 B1 (19) NO NORGE (51) Int Cl. A61K 31/137 (2006.01) A61P 25/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.02 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2148670 B1 (19) NO NORGE (51) Int Cl. A61K 31/137 (2006.01) A61P 25/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.02 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2011486 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.09.17 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2011486 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.09.17 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2129377 B1 (19) NO NORGE (51) Int Cl. A61K 31/451 (2006.01) A61K 9/08 (2006.01) A61P 25/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.23
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2129377 B1 (19) NO NORGE (51) Int Cl. A61K 31/451 (2006.01) A61K 9/08 (2006.01) A61P 25/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.23
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2246321 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/135 (2006.01) C07C 211/42 (2006.01) Patentstyret (21) Oversettelse publisert 2011.12.12
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2246321 B1 (19) NO NORGE (51) Int Cl. A61K 9/20 (2006.01) A61K 31/135 (2006.01) C07C 211/42 (2006.01) Patentstyret (21) Oversettelse publisert 2011.12.12
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2114970 B1 (19) NO NORGE (51) Int Cl. C07F 9/58 (2006.01) A61K 31/44 (2006.01) A61P 1/00 (2006.01) A61P 11/06 (2006.01) A61P 19/02 (2006.01) A61P
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2114970 B1 (19) NO NORGE (51) Int Cl. C07F 9/58 (2006.01) A61K 31/44 (2006.01) A61P 1/00 (2006.01) A61P 11/06 (2006.01) A61P 19/02 (2006.01) A61P
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2274977 B1 (19) NO NORGE (1) Int Cl. A01K 83/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2274977 B1 (19) NO NORGE (1) Int Cl. A01K 83/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2178851 B1 (19) NO NORGE (51) Int Cl. C07D 261/08 (2006.01) A61K 31/42 (2006.01) A61P 3/06 (2006.01) C07D 413/12 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2178851 B1 (19) NO NORGE (51) Int Cl. C07D 261/08 (2006.01) A61K 31/42 (2006.01) A61P 3/06 (2006.01) C07D 413/12 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2445326 B1 (19) NO NORGE (51) Int Cl. H05K 5/02 (2006.01) B43K 23/12 (2006.01) B43K 24/06 (2006.01) H01R 13/60 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2445326 B1 (19) NO NORGE (51) Int Cl. H05K 5/02 (2006.01) B43K 23/12 (2006.01) B43K 24/06 (2006.01) H01R 13/60 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2272978 B1 (19) NO NORGE (51) Int Cl. C12Q 1/68 (2006.01) Patentstyret (21) Oversettelse publisert 2012.08.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2272978 B1 (19) NO NORGE (51) Int Cl. C12Q 1/68 (2006.01) Patentstyret (21) Oversettelse publisert 2012.08.13 (80) Dato for Den Europeiske Patentmyndighets
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2125711 B1 (19) NO NORGE (51) Int Cl. C07C 321/20 (2006.01) A61K 31/216 (2006.01) A61K 31/421 (2006.01) A61K 31/4402 (2006.01) A61K 31/495 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2125711 B1 (19) NO NORGE (51) Int Cl. C07C 321/20 (2006.01) A61K 31/216 (2006.01) A61K 31/421 (2006.01) A61K 31/4402 (2006.01) A61K 31/495 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 240726 B1 (19) NO NORGE (1) Int Cl. H0K 3/36 (2006.01) H0K 3/42 (2006.01) H0K 3/46 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.17 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 240726 B1 (19) NO NORGE (1) Int Cl. H0K 3/36 (2006.01) H0K 3/42 (2006.01) H0K 3/46 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.17 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") 1 3 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2207775 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/5377 (2006.01) A61P 3/06 (2006.01) C07D 401/14 (2006.01) C07D 413/14 (2006.01)
1 3 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2207775 B1 (19) NO NORGE (51) Int Cl. C07D 401/12 (2006.01) A61K 31/5377 (2006.01) A61P 3/06 (2006.01) C07D 401/14 (2006.01) C07D 413/14 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP22342 (12) Oversettelse av europeisk patentskrift (11) NO/EP 22342 B1 (19) NO NORGE (1) Int Cl. F2D 23/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske Patentmyndighets
NO/EP22342 (12) Oversettelse av europeisk patentskrift (11) NO/EP 22342 B1 (19) NO NORGE (1) Int Cl. F2D 23/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.27 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 213696 B1 (19) NO NORGE (1) Int Cl. B23K 9/32 (2006.01) B23K 9/28 (2006.01) Patentstyret (21) Oversettelse publisert 2014.04.07 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 213696 B1 (19) NO NORGE (1) Int Cl. B23K 9/32 (2006.01) B23K 9/28 (2006.01) Patentstyret (21) Oversettelse publisert 2014.04.07 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 11438 B1 (19) NO NORGE (1) Int Cl. E04B 1/343 (06.01) B63B 29/02 (06.01) Patentstyret (21) Oversettelse publisert.02.23 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 11438 B1 (19) NO NORGE (1) Int Cl. E04B 1/343 (06.01) B63B 29/02 (06.01) Patentstyret (21) Oversettelse publisert.02.23 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2128505 B1 (19) NO NORGE (51) Int Cl. F16L 9/12 (2006.01) F16L 3/14 (2006.01) F16L 11/127 (2006.01) F24F 13/02 (2006.01) H05F 3/02 (2006.01) Patentstyret
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2128505 B1 (19) NO NORGE (51) Int Cl. F16L 9/12 (2006.01) F16L 3/14 (2006.01) F16L 11/127 (2006.01) F24F 13/02 (2006.01) H05F 3/02 (2006.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2285808 B1 (19) NO NORGE (51) Int Cl. C07D 471/20 (2006.01) A61K 31/407 (2006.01) A61K 31/424 (2006.01) A61K 31/437 (2006.01) A61K 31/438 (2006.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2285808 B1 (19) NO NORGE (51) Int Cl. C07D 471/20 (2006.01) A61K 31/407 (2006.01) A61K 31/424 (2006.01) A61K 31/437 (2006.01) A61K 31/438 (2006.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2170890 B1 (19) NO NORGE (51) Int Cl. C07D 487/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.03.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2170890 B1 (19) NO NORGE (51) Int Cl. C07D 487/04 (2006.01) Patentstyret (21) Oversettelse publisert 2012.03.12 (80) Dato for Den Europeiske Patentmyndighets
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 17118 B1 (19) NO NORGE (1) Int Cl. B60M 1/06 (06.01) B60M 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.09.29 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 17118 B1 (19) NO NORGE (1) Int Cl. B60M 1/06 (06.01) B60M 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.09.29 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2173868 B1 (19) NO NORGE (51) Int Cl. C12N 9/50 (2006.01) C07K 14/415 (2006.01) C12N 15/29 (2006.01) C12N 15/57 (2006.01) C12N 15/81 (2006.01) A23J
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2173868 B1 (19) NO NORGE (51) Int Cl. C12N 9/50 (2006.01) C07K 14/415 (2006.01) C12N 15/29 (2006.01) C12N 15/57 (2006.01) C12N 15/81 (2006.01) A23J
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2613860 B1 (19) NO NORGE (51) Int Cl. B01D 15/18 (2006.01) C11B 3/10 (2006.01) C11C 1/00 (2006.01) C11C 1/08 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2613860 B1 (19) NO NORGE (51) Int Cl. B01D 15/18 (2006.01) C11B 3/10 (2006.01) C11C 1/00 (2006.01) C11C 1/08 (2006.01) Patentstyret (21) Oversettelse
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 224294 B1 (19) NO NORGE (1) Int Cl. F16K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.10 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 224294 B1 (19) NO NORGE (1) Int Cl. F16K 31/44 (2006.01) Patentstyret (21) Oversettelse publisert 2012.04.10 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2252286 B1 (19) NO NORGE (51) Int Cl. A61K 31/357 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.16 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2252286 B1 (19) NO NORGE (51) Int Cl. A61K 31/357 (2006.01) Patentstyret (21) Oversettelse publisert 2012.01.16 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 237066 B1 (19) NO NORGE (1) Int Cl. E06C 1/12 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 237066 B1 (19) NO NORGE (1) Int Cl. E06C 1/12 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 270722 B1 (19) NO NORGE (1) Int Cl. F21V 23/02 (06.01) F21S 8/02 (06.01) F21V 23/00 (06.01) Patentstyret (21) Oversettelse publisert 14.03. (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 270722 B1 (19) NO NORGE (1) Int Cl. F21V 23/02 (06.01) F21S 8/02 (06.01) F21V 23/00 (06.01) Patentstyret (21) Oversettelse publisert 14.03. (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 242166 B1 (19) NO NORGE (1) Int Cl. G06K 19/077 (06.01) G06K 19/06 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 242166 B1 (19) NO NORGE (1) Int Cl. G06K 19/077 (06.01) G06K 19/06 (06.01) Patentstyret (21) Oversettelse publisert 14.02.24 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2216387 B1 (19) NO NORGE (51) Int Cl. C10L 5/44 (2006.01) C10L 5/14 (2006.01) C10L 5/36 (2006.01) Patentstyret (21) Oversettelse publisert 2013.05.06
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2216387 B1 (19) NO NORGE (51) Int Cl. C10L 5/44 (2006.01) C10L 5/14 (2006.01) C10L 5/36 (2006.01) Patentstyret (21) Oversettelse publisert 2013.05.06
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2672278 B1 (19) NO NORGE (1) Int Cl. G01R 1/067 (2006.01) G01R 1/04 (2006.01) G01R 19/1 (2006.01) Patentstyret (21) Oversettelse publisert 201.04.20
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2672278 B1 (19) NO NORGE (1) Int Cl. G01R 1/067 (2006.01) G01R 1/04 (2006.01) G01R 19/1 (2006.01) Patentstyret (21) Oversettelse publisert 201.04.20
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2097141 B1 (19) NO NORGE (51) Int Cl. A62B 35/00 (2006.01) Patentstyret (21) Oversettelse publisert 2013.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2097141 B1 (19) NO NORGE (51) Int Cl. A62B 35/00 (2006.01) Patentstyret (21) Oversettelse publisert 2013.08.19 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2317621 B1 (19) NO NORGE (1) Int Cl. H02G 3/12 (06.01) Patentstyret (21) Oversettelse publisert 1.02.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2317621 B1 (19) NO NORGE (1) Int Cl. H02G 3/12 (06.01) Patentstyret (21) Oversettelse publisert 1.02.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 223094 B1 (19) NO NORGE (1) Int Cl. A43B 7/32 (06.01) A43B 7/12 (06.01) A43B 7/34 (06.01) A43B 13/12 (06.01) A43B 13/41 (06.01) B29D 3/14 (.01) Patentstyret
(12) Oversettelse av europeisk patentskrift (11) NO/EP 223094 B1 (19) NO NORGE (1) Int Cl. A43B 7/32 (06.01) A43B 7/12 (06.01) A43B 7/34 (06.01) A43B 13/12 (06.01) A43B 13/41 (06.01) B29D 3/14 (.01) Patentstyret
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2384729 B1 (19) NO NORGE (1) Int Cl. A61G /12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2384729 B1 (19) NO NORGE (1) Int Cl. A61G /12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.04.08 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2708433 B1 (19) NO NORGE (1) Int Cl. B61B 1/02 (2006.01) B61B 12/02 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.12 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2708433 B1 (19) NO NORGE (1) Int Cl. B61B 1/02 (2006.01) B61B 12/02 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.12 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse
 Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2175588 B2 (19) NO NORGE (51) Int Cl. H04L 12/14 (2006.01) H04L 29/08 (2006.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2175588 B2 (19) NO NORGE (51) Int Cl. H04L 12/14 (2006.01) H04L 29/08 (2006.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse
(12) Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse
 Oversettelse av europeisk patentskrift. Avviker fra Patent B1 etter innsigelse") (12) Oversettelse av europeisk patentskrift (11) NO/EP 217368 B2 (19) NO NORGE (1) Int Cl. B42D / (06.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse publisert.04. (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 217368 B2 (19) NO NORGE (1) Int Cl. B42D / (06.01) Patentstyret Avviker fra Patent B1 etter innsigelse (21) Oversettelse publisert.04. (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22799 B1 (19) NO NORGE (1) Int Cl. A61K 31/23 (06.01) A61K 31/047 (06.01) A61K 31/231 (06.01) A61K 31/232 (06.01) A61K 31/3 (06.01) A61K 31/93 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22799 B1 (19) NO NORGE (1) Int Cl. A61K 31/23 (06.01) A61K 31/047 (06.01) A61K 31/231 (06.01) A61K 31/232 (06.01) A61K 31/3 (06.01) A61K 31/93 (06.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 218466 B1 (19) NO NORGE (1) Int Cl. B67C 3/26 (06.01) B6D 47/ (06.01) B67C 7/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02. (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 218466 B1 (19) NO NORGE (1) Int Cl. B67C 3/26 (06.01) B6D 47/ (06.01) B67C 7/00 (06.01) Patentstyret (21) Oversettelse publisert 12.02. (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 24012 B1 (19) NO NORGE (1) Int Cl. B2C 1/00 (2006.01) B2C 1/06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.12.22 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 24012 B1 (19) NO NORGE (1) Int Cl. B2C 1/00 (2006.01) B2C 1/06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.12.22 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2491293 B1 (19) NO NORGE (1) Int Cl. F17C 3/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.2 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2491293 B1 (19) NO NORGE (1) Int Cl. F17C 3/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.2 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 88493 B1 (19) NO NORGE (1) Int Cl. G06F 1/00 (06.01) H01L 23/34 (06.01) G06F 1/ (06.01) Patentstyret (21) Oversettelse publisert 13.04.22 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 88493 B1 (19) NO NORGE (1) Int Cl. G06F 1/00 (06.01) H01L 23/34 (06.01) G06F 1/ (06.01) Patentstyret (21) Oversettelse publisert 13.04.22 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2082973 B1 (19) NO NORGE (1) Int Cl. B6D 81/34 (2006.01) Patentstyret (21) Oversettelse publisert 2014.06.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2082973 B1 (19) NO NORGE (1) Int Cl. B6D 81/34 (2006.01) Patentstyret (21) Oversettelse publisert 2014.06.02 (80) Dato for Den Europeiske Patentmyndighets
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2238877 B1 (19) NO NORGE (1) Int Cl. A47J 31/08 (06.01) Patentstyret (21) Oversettelse publisert 13.03.11 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2238877 B1 (19) NO NORGE (1) Int Cl. A47J 31/08 (06.01) Patentstyret (21) Oversettelse publisert 13.03.11 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2636033 B1 (19) NO NORGE (51) Int Cl. Patentstyret G09B 23/28 (2006.01) G09B 23/30 (2006.01) (21) Oversettelse publisert 2015.11.09 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2636033 B1 (19) NO NORGE (51) Int Cl. Patentstyret G09B 23/28 (2006.01) G09B 23/30 (2006.01) (21) Oversettelse publisert 2015.11.09 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2261144 B1 (19) NO NORGE (1) Int Cl. B6G 21/00 (06.01) B6G 21/08 (06.01) Patentstyret (21) Oversettelse publisert 13.07.08 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2261144 B1 (19) NO NORGE (1) Int Cl. B6G 21/00 (06.01) B6G 21/08 (06.01) Patentstyret (21) Oversettelse publisert 13.07.08 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 222 B1 (19) NO NORGE (1) Int Cl. F16F 1/376 (06.01) F16F 1/373 (06.01) F16F 1/08 (06.01) Patentstyret (21) Oversettelse publisert 13.02.18 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 222 B1 (19) NO NORGE (1) Int Cl. F16F 1/376 (06.01) F16F 1/373 (06.01) F16F 1/08 (06.01) Patentstyret (21) Oversettelse publisert 13.02.18 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2231428 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) Patentstyret (21) Oversettelse publisert 12.11.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2231428 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) Patentstyret (21) Oversettelse publisert 12.11.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2243894 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2243894 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 201.01.26 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP2770 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2770 B1 (19) NO NORGE (1) Int Cl. B23K 3/00 (06.01) C21D 6/00 (06.01) C21D 9/04 (06.01) C22C 38/00 (06.01) C22C 38/44 (06.01) Patentstyret
NO/EP2770 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2770 B1 (19) NO NORGE (1) Int Cl. B23K 3/00 (06.01) C21D 6/00 (06.01) C21D 9/04 (06.01) C22C 38/00 (06.01) C22C 38/44 (06.01) Patentstyret
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2311023 B1 (19) NO NORGE (51) Int Cl. G09F 17/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2311023 B1 (19) NO NORGE (51) Int Cl. G09F 17/00 (2006.01) Patentstyret (21) Oversettelse publisert 2014.02.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22619 B1 (19) NO NORGE (1) Int Cl. B21D 1/4 (2006.01) B21K 21/04 (2006.01) F42B /02 (2006.01) F42B /188 (2006.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22619 B1 (19) NO NORGE (1) Int Cl. B21D 1/4 (2006.01) B21K 21/04 (2006.01) F42B /02 (2006.01) F42B /188 (2006.01) Patentstyret (21) Oversettelse publisert
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 26098 B1 (19) NO NORWAY (1) Int Cl. C07K 14/08 (06.01) C12Q 1/70 (06.01) Norwegian Industrial Property Office (21) Translation Published 16..24
(12) Translation of european patent specification (11) NO/EP 26098 B1 (19) NO NORWAY (1) Int Cl. C07K 14/08 (06.01) C12Q 1/70 (06.01) Norwegian Industrial Property Office (21) Translation Published 16..24
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 21181 B1 (19) NO NORGE (1) Int Cl. F16L 2/00 (2006.01) F16L 33/26 (2006.01) H01P 1/04 (2006.01) Patentstyret (21) Oversettelse publisert 2013.10.28
(12) Oversettelse av europeisk patentskrift (11) NO/EP 21181 B1 (19) NO NORGE (1) Int Cl. F16L 2/00 (2006.01) F16L 33/26 (2006.01) H01P 1/04 (2006.01) Patentstyret (21) Oversettelse publisert 2013.10.28
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2264391 B1 (19) NO NORGE (1) Int Cl. F27D 3/1 (2006.01) C21B 7/12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.11.18 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2264391 B1 (19) NO NORGE (1) Int Cl. F27D 3/1 (2006.01) C21B 7/12 (2006.01) Patentstyret (21) Oversettelse publisert 2013.11.18 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2217383 B1 (19) NO NORGE (1) Int Cl. B0B 12/00 (06.01) B0B 11/00 (06.01) G01F 11/02 (06.01) G01F 1/07 (06.01) G07C 3/04 (06.01) Patentstyret (21)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2217383 B1 (19) NO NORGE (1) Int Cl. B0B 12/00 (06.01) B0B 11/00 (06.01) G01F 11/02 (06.01) G01F 1/07 (06.01) G07C 3/04 (06.01) Patentstyret (21)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2146022 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.11.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2146022 B1 (19) NO NORGE (1) Int Cl. E04F /06 (2006.01) Patentstyret (21) Oversettelse publisert 2014.11.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22442 B1 (19) NO NORGE (1) Int Cl. G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13..28 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22442 B1 (19) NO NORGE (1) Int Cl. G07B 1/00 (11.01) Patentstyret (21) Oversettelse publisert 13..28 (80) Dato for Den Europeiske Patentmyndighets
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 21847 B1 (19) NO NORGE (1) Int Cl. F24F 7/08 (06.01) F24F 11/04 (06.01) F24F 12/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 21847 B1 (19) NO NORGE (1) Int Cl. F24F 7/08 (06.01) F24F 11/04 (06.01) F24F 12/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 261673 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) B60H 1/00 (06.01) Patentstyret (21) Oversettelse publisert 1.01.12 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 261673 B1 (19) NO NORGE (1) Int Cl. B60H 1/32 (06.01) B60H 1/00 (06.01) Patentstyret (21) Oversettelse publisert 1.01.12 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 99826 B1 (19) NO NORGE (1) Int Cl. C07K 16/18 (06.01) A61K 39/39 (06.01) A61P 2/28 (06.01) Patentstyret (21) Oversettelse publisert 14.02.17 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 99826 B1 (19) NO NORGE (1) Int Cl. C07K 16/18 (06.01) A61K 39/39 (06.01) A61P 2/28 (06.01) Patentstyret (21) Oversettelse publisert 14.02.17 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 20789 B1 (19) NO NORGE (1) Int Cl. B61D 1/00 (06.01) B61D 17/ (06.01) B61D 23/00 (06.01) Patentstyret (21) Oversettelse publisert 12.06.04 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 20789 B1 (19) NO NORGE (1) Int Cl. B61D 1/00 (06.01) B61D 17/ (06.01) B61D 23/00 (06.01) Patentstyret (21) Oversettelse publisert 12.06.04 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2096736 B1 (19) NO NORGE (1) Int Cl. H02K 1/32 (2006.01) H02K 3/24 (2006.01) H02K 9/00 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2096736 B1 (19) NO NORGE (1) Int Cl. H02K 1/32 (2006.01) H02K 3/24 (2006.01) H02K 9/00 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2184425 B1 (19) NO NORGE (51) Int Cl. E05B 17/20 (2006.01) E05B 63/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.02.06 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2184425 B1 (19) NO NORGE (51) Int Cl. E05B 17/20 (2006.01) E05B 63/00 (2006.01) Patentstyret (21) Oversettelse publisert 2012.02.06 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 19724 B1 (19) NO NORGE (1) Int Cl. B63H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 12.12. (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 19724 B1 (19) NO NORGE (1) Int Cl. B63H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 12.12. (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2146836 B1 (19) NO NORGE (1) Int Cl. A47G 9/ (06.01) B26D 3/00 (06.01) B26D 3/28 (06.01) B29C 44/6 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2146836 B1 (19) NO NORGE (1) Int Cl. A47G 9/ (06.01) B26D 3/00 (06.01) B26D 3/28 (06.01) B29C 44/6 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2148223 B1 (19) NO NORGE (1) Int Cl. G01V 3/ (06.01) G01V 3/24 (06.01) Patentstyret (21) Oversettelse publisert 13.03.04 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2148223 B1 (19) NO NORGE (1) Int Cl. G01V 3/ (06.01) G01V 3/24 (06.01) Patentstyret (21) Oversettelse publisert 13.03.04 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 7044 B1 (19) NO NORGE (1) Int Cl. A61K 36/18 (06.01) A61K 33/04 (06.01) A61K 33/18 (06.01) A61K 33/ (06.01) A61K 36/22 (06.01) A61K 36/28 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 7044 B1 (19) NO NORGE (1) Int Cl. A61K 36/18 (06.01) A61K 33/04 (06.01) A61K 33/18 (06.01) A61K 33/ (06.01) A61K 36/22 (06.01) A61K 36/28 (06.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2497702 B1 (19) NO NORGE (1) Int Cl. B62H 3/02 (06.01) B62H /00 (06.01) B62M 6/80 (.01) Patentstyret (21) Oversettelse publisert 1.03.16 (80) Dato
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2497702 B1 (19) NO NORGE (1) Int Cl. B62H 3/02 (06.01) B62H /00 (06.01) B62M 6/80 (.01) Patentstyret (21) Oversettelse publisert 1.03.16 (80) Dato
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 08940 B1 (19) NO NORGE (1) Int Cl. B6D 2/2 (06.01) A47G 19/34 (06.01) B6D 83/06 (06.01) G01F 11/26 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 08940 B1 (19) NO NORGE (1) Int Cl. B6D 2/2 (06.01) A47G 19/34 (06.01) B6D 83/06 (06.01) G01F 11/26 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2012637 B1 NORGE (19) NO (1) Int Cl. A47K 13/00 (2006.01) Patentstyret (4) Oversettelse publisert: 20.08.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2012637 B1 NORGE (19) NO (1) Int Cl. A47K 13/00 (2006.01) Patentstyret (4) Oversettelse publisert: 20.08.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") NO/EP20 (12) Translation of european patent specification (11) NO/EP 20 B1 (19) NO NORWAY (1) Int Cl. C07K 16/28 (2006.01) A61P 3/00 (2006.01) A61P 37/00 (2006.01) Norwegian Industrial Property Office
NO/EP20 (12) Translation of european patent specification (11) NO/EP 20 B1 (19) NO NORWAY (1) Int Cl. C07K 16/28 (2006.01) A61P 3/00 (2006.01) A61P 37/00 (2006.01) Norwegian Industrial Property Office
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2216871 B1 (19) NO NORGE (51) Int Cl. H02J 7/00 (2006.01) H01R 13/22 (2006.01) H01R 13/62 (2006.01) Patentstyret (21) Oversettelse publisert 2014.09.08
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2216871 B1 (19) NO NORGE (51) Int Cl. H02J 7/00 (2006.01) H01R 13/22 (2006.01) H01R 13/62 (2006.01) Patentstyret (21) Oversettelse publisert 2014.09.08
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2244923 B1 (19) NO NORGE (1) Int Cl. B61K 9/ (2006.01) Patentstyret (21) Oversettelse publisert 2013.09.30 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2244923 B1 (19) NO NORGE (1) Int Cl. B61K 9/ (2006.01) Patentstyret (21) Oversettelse publisert 2013.09.30 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2236434 B1 (19) NO NORGE (1) Int Cl. B6D 77/04 (06.01) B6D 77/06 (06.01) Patentstyret (21) Oversettelse publisert 11.12.19 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2236434 B1 (19) NO NORGE (1) Int Cl. B6D 77/04 (06.01) B6D 77/06 (06.01) Patentstyret (21) Oversettelse publisert 11.12.19 (80) Dato for Den Europeiske
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 211778 B1 (19) NO NORWAY (1) Int Cl. A61K 38/16 (06.01) C07K 7/08 (06.01) Norwegian Industrial Property Office (21) Translation Published 1.09.07
(12) Translation of european patent specification (11) NO/EP 211778 B1 (19) NO NORWAY (1) Int Cl. A61K 38/16 (06.01) C07K 7/08 (06.01) Norwegian Industrial Property Office (21) Translation Published 1.09.07
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 216340 B1 (19) NO NORGE (1) Int Cl. B60C 11/11 (06.01) B60C 11/03 (06.01) B60C 11/12 (06.01) Patentstyret (21) Oversettelse publisert 12.12.03 (80)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 216340 B1 (19) NO NORGE (1) Int Cl. B60C 11/11 (06.01) B60C 11/03 (06.01) B60C 11/12 (06.01) Patentstyret (21) Oversettelse publisert 12.12.03 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2477830 B1 (19) NO NORGE (1) Int Cl. B60K 1/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2477830 B1 (19) NO NORGE (1) Int Cl. B60K 1/00 (06.01) Patentstyret (21) Oversettelse publisert 13.12.02 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 22473 B1 (19) NO NORGE (1) Int Cl. H01H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 1.0.04 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 22473 B1 (19) NO NORGE (1) Int Cl. H01H 23/02 (06.01) Patentstyret (21) Oversettelse publisert 1.0.04 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 240126 B1 (19) NO NORGE (1) Int Cl. C07D 211/62 (06.01) A61K 31/16 (06.01) A61K 31/44 (06.01) A61K 31/0 (06.01) A61K 31/06 (06.01) C07D 7/277 (06.01)
(12) Oversettelse av europeisk patentskrift (11) NO/EP 240126 B1 (19) NO NORGE (1) Int Cl. C07D 211/62 (06.01) A61K 31/16 (06.01) A61K 31/44 (06.01) A61K 31/0 (06.01) A61K 31/06 (06.01) C07D 7/277 (06.01)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 9863 B1 (19) NO NORGE (1) Int Cl. E04B 2/96 (06.01) Patentstyret (21) Oversettelse publisert 13.09.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 9863 B1 (19) NO NORGE (1) Int Cl. E04B 2/96 (06.01) Patentstyret (21) Oversettelse publisert 13.09.09 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP2563678 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2563678 B1 (19) NO NORGE (51) Int Cl. B65D 6/00 (2006.01) Patentstyret (21) Oversettelse publisert 2015.01.19 (80) Dato for Den Europeiske
NO/EP2563678 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2563678 B1 (19) NO NORGE (51) Int Cl. B65D 6/00 (2006.01) Patentstyret (21) Oversettelse publisert 2015.01.19 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2003466 B1 (19) NO NORGE (1) Int Cl. G01S /02 (2010.01) Patentstyret (21) Oversettelse publisert 2014.07.14 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2003466 B1 (19) NO NORGE (1) Int Cl. G01S /02 (2010.01) Patentstyret (21) Oversettelse publisert 2014.07.14 (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2231998 B1 (19) NO NORWAY (51) Int Cl. E21B 17/10 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2015.10.05 (80)
(12) Translation of european patent specification (11) NO/EP 2231998 B1 (19) NO NORWAY (51) Int Cl. E21B 17/10 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2015.10.05 (80)
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2231500 B1 (19) NO NORGE (51) Int Cl. B66F 9/00 (2006.01) B60P 1/02 (2006.01) B60P 3/022 (2006.01) B62B 3/065 (2006.01) B66D 1/00 (2006.01) B66F 9/06
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2231500 B1 (19) NO NORGE (51) Int Cl. B66F 9/00 (2006.01) B60P 1/02 (2006.01) B60P 3/022 (2006.01) B62B 3/065 (2006.01) B66D 1/00 (2006.01) B66F 9/06
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2328616 B1 (19) NO NORWAY (1) Int Cl. A61K 39/39 (2006.01) C07K 16/18 (2006.01) Norwegian Industrial Property Office (21) Translation Published
(12) Translation of european patent specification (11) NO/EP 2328616 B1 (19) NO NORWAY (1) Int Cl. A61K 39/39 (2006.01) C07K 16/18 (2006.01) Norwegian Industrial Property Office (21) Translation Published
PATENTKRAV. og en første lett kjede og en andre lett kjede, hvor første og andre lette kjeder er forskjellige.
 1 PATENTKRAV 1. Isolert monoklonalt antistoff som bærer en forskjellig spesifisitet i hvert kombinasjonssete og som består av to kopier av et enkelt tung kjede polypeptid og en første lett kjede og en
1 PATENTKRAV 1. Isolert monoklonalt antistoff som bærer en forskjellig spesifisitet i hvert kombinasjonssete og som består av to kopier av et enkelt tung kjede polypeptid og en første lett kjede og en
(86) Europeisk innleveringsdag
 Europeisk innleveringsdag") (12) Oversettelse av europeisk patentskrift (11) NO/EP 297978 B1 (19) NO NORGE (1) Int Cl. A41B 9/02 (06.01) Patentstyret (21) Oversettelse publisert 14.03.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 297978 B1 (19) NO NORGE (1) Int Cl. A41B 9/02 (06.01) Patentstyret (21) Oversettelse publisert 14.03.17 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 273 B1 (19) NO NORGE (51) Int Cl. B41J 2/175 (06.01) Patentstyret (21) Oversettelse publisert 14.05.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 273 B1 (19) NO NORGE (51) Int Cl. B41J 2/175 (06.01) Patentstyret (21) Oversettelse publisert 14.05.12 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2117944 B1 (19) NO NORGE (1) Int Cl. B6D 21/02 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2117944 B1 (19) NO NORGE (1) Int Cl. B6D 21/02 (2006.01) Patentstyret (21) Oversettelse publisert 2011.09.0 (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2229212 B1 (19) NO NORWAY (51) Int Cl. A61N 1/36 (2006.01) H01L 27/144 (2006.01) Norwegian Industrial Property Office (21) Translation Published
(12) Translation of european patent specification (11) NO/EP 2229212 B1 (19) NO NORWAY (51) Int Cl. A61N 1/36 (2006.01) H01L 27/144 (2006.01) Norwegian Industrial Property Office (21) Translation Published
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 246764 B1 (19) NO NORGE (1) Int Cl. F2C 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.13 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 246764 B1 (19) NO NORGE (1) Int Cl. F2C 3/04 (06.01) Patentstyret (21) Oversettelse publisert 14.01.13 (80) Dato for Den Europeiske Patentmyndighets
europeisk patentskrift
 (12) Oversettelse av europeisk patentskrift (11) NO/EP 2404809 B1 (19) NO NORGE (1) Int Cl. B62D 21/02 (2006.01) Patentstyret (21) Oversettelse publisert 2013.07.22 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2404809 B1 (19) NO NORGE (1) Int Cl. B62D 21/02 (2006.01) Patentstyret (21) Oversettelse publisert 2013.07.22 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 264739 B1 (19) NO NORGE (1) Int Cl. A61F 13/00 (06.01) A61B 17/132 (06.01) A61F 13/64 (06.01) A61F 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift (11) NO/EP 264739 B1 (19) NO NORGE (1) Int Cl. A61F 13/00 (06.01) A61B 17/132 (06.01) A61F 13/64 (06.01) A61F 1/00 (06.01) Patentstyret (21) Oversettelse publisert
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP28769 (12) Oversettelse av europeisk patentskrift (11) NO/EP 28769 B1 (19) NO NORGE (1) Int Cl. F17D 1/18 (06.01) F16L 3/00 (06.01) Patentstyret (21) Oversettelse publisert 1.04. (80) Dato for Den
NO/EP28769 (12) Oversettelse av europeisk patentskrift (11) NO/EP 28769 B1 (19) NO NORGE (1) Int Cl. F17D 1/18 (06.01) F16L 3/00 (06.01) Patentstyret (21) Oversettelse publisert 1.04. (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2093737 B1 (19) NO NORGE (51) Int Cl. G08B 29/06 (2006.01) G08B 29/12 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.10 (80) Dato for
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2093737 B1 (19) NO NORGE (51) Int Cl. G08B 29/06 (2006.01) G08B 29/12 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.10 (80) Dato for
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2131860 B1 (19) NO NORGE (51) Int Cl. A61K 39/395 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2131860 B1 (19) NO NORGE (51) Int Cl. A61K 39/395 (2006.01) Patentstyret (21) Oversettelse publisert 2014.03.24 (80) Dato for Den Europeiske Patentmyndighets
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 2113323 B1 (19) NO NORGE (1) Int Cl. B23B 31/02 (2006.01) B23B 31/20 (2006.01) Patentstyret (21) Oversettelse publisert 2012.11.19 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift (11) NO/EP 2113323 B1 (19) NO NORGE (1) Int Cl. B23B 31/02 (2006.01) B23B 31/20 (2006.01) Patentstyret (21) Oversettelse publisert 2012.11.19 (80) Dato for Den
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") (12) Oversettelse av europeisk patentskrift (11) NO/EP 229688 B1 (19) NO NORGE (1) Int Cl. B29B 17/02 (06.01) D21B 1/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.18 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift (11) NO/EP 229688 B1 (19) NO NORGE (1) Int Cl. B29B 17/02 (06.01) D21B 1/02 (06.01) Patentstyret (21) Oversettelse publisert 13.11.18 (80) Dato for Den Europeiske
(12) Oversettelse av europeisk patentskrift
 Oversettelse av europeisk patentskrift") NO/EP918 (12) Oversettelse av europeisk patentskrift (11) NO/EP 918 B1 (19) NO NORGE (1) Int Cl. H02J 7/00 (06.01) Patentstyret (21) Oversettelse publisert 14.02.03 (80) Dato for Den Europeiske Patentmyndighets
NO/EP918 (12) Oversettelse av europeisk patentskrift (11) NO/EP 918 B1 (19) NO NORGE (1) Int Cl. H02J 7/00 (06.01) Patentstyret (21) Oversettelse publisert 14.02.03 (80) Dato for Den Europeiske Patentmyndighets
(12) Translation of european patent specification
 Translation of european patent specification") (12) Translation of european patent specification (11) NO/EP 2616307 B1 (19) NO NORWAY (51) Int Cl. B61L 29/28 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2016.03.07 (80)
(12) Translation of european patent specification (11) NO/EP 2616307 B1 (19) NO NORWAY (51) Int Cl. B61L 29/28 (2006.01) Norwegian Industrial Property Office (21) Translation Published 2016.03.07 (80)