Bioenergetikk og Krebs syklus Oksidativ fosforylering. Bioenergetikk, IA 2015
|
|
|
- Magnus Ulriksen
- 7 år siden
- Visninger:
Transkript
1 1 Bioenergetikk og Krebs syklus Oksidativ fosforylering Bioenergetikk, IA 2015
2 2 Relevante læringsmål, Biokjemi: beskrive den generelle strukturen til karbohydrater, lipider, aminosyrer og nukleinsyrer, og forklare stoffenes fysikalsk-kjemiske egenskaper ut fra dette gjøre rede for hovedfunksjonene til karbohydrater, lipider, nukleinsyrer og proteiner beskrive nedbrytingen av glykogen, disakkarider, monosakkarider og fettsyrer, og forklare hvordan slik nedbryting er koplet til dannelse av ATP beskrive syntesen av glukose og glykogen definere begrepene vitamin og mineral, angi de viktigste mineraler og vitaminer og beskrive deres biologiske funksjoner Relevant studiemateriell: Lehninger; Principles of biochemistry, 6 th Ed., Kapittel 13 Lippincott s; Biochemistry, 5 th Ed., Kapittel 6 Lippincott s Illstrated Reviews of Biochemistry, 6th ed., kap 6, 9 Baynes; Medical biochemistry, 3 rd Ed., Kapittel 9, 14 Bioenergetikk
3 3 Det store bildet Bioenergetikk
4 4 Bioenergetikk og termodynamikk Bioenergetikk er applikasjon av termodynamiske lover til biokjemiske systemer. Beskriver energioverføringer i celler og hvordan de kjemiske prosessene kan kvantifiseres. Dagens tema: 1. Termodynamikk 2. Fosforylgruppeoverføringer og energioverføringer 3. Oksidasjon og reduksjonsreaksjoner i celler 4. Massevirkningsloven Bioenergetikk

5 5 Energi kan anta ulike former Potensiell energi kan omformes til bevegelsesenergi, som igjen kan omformes til varmeenergi Noe av bevegelsesenergien kan benyttes til å løfte en bøtte vann, som derved får økt potensiell energi. Tilsvarende mindre energi avgis som varme Potensiell bevegelsesenergi lagret i vannbøtta kan benyttes til å drive ulike hydrauliske maskiner, og f. eks. omformes til elektrisk energi, som igjen kan omformes til kjemisk energi (eks. kunstgjødselproduksjon) Bioenergetikk

6 6 Massevirkningsloven I en kjemisk reaksjon: aa + bb cc + dd vil reaksjonen forløpe inntil en likevekt er innstilt seg mellom produktene og reaktantene. Denne likevekten er uttrykt som følger: K eq = [C] c [D] d [A] a [B] b Cato M Guldberg og Peter Waage Hastigheten av de to reaksjonene er proposjonal med konsentrasjonen av reaktantene som inngår. Hvis mengden av en av reaktantene eller produktene endres, vil konsentrasjonene av de andre komponentene også endres for å opprettholde likevekten. Dette har stor betydning for biologiske reaksjoner. Bioenergetikk
7 7 Viktige termodynamiske definisjoner H H S G Entalpi: Varmeenergien, eller den samlede energien i et system (feks i en bestemt mengde av en kjemisk forbindelse) Forandring i entalpi i løpet av en reaksjon. Bestemmes av antallet og typen av kjemiske bindinger i reaktanter og produkt. Er H positiv, sier vi at reaksjonen er endoterm (varme tas opp). Er H negativ, sier vi den er eksoterm (varme avgis) Entropi: Grad av uorden i et system. Den delen av energien som ikke kan utføre arbeid Gibbs fri energi: Den delen av energien i et system som kan utføre arbeid. Er G positiv, sier vi at reaksjonen er endergon. Er G negativ, sier vi den er eksergon. Disse verdiene vil være relatert til hverandre som følger: G = H - T S Bioenergetikk
8 8 Massevirkningsloven / likevekt I en kjemisk reaksjon A B vil alltid likevekten være forskjøvet mot den siden som har lavest innhold av fri energi, G. Hvis G for reaksjonen er negativ, vil altså likevekten være forskjøvet mot høyre. Standard fri energiendring G o : 25 o C (=298 K) og 1 M reaktanter. G o = -RT ln [B]/[A] = -RT ln K eq R = gasskonstanten = J/mol x K Merk at en kan utføre arbeid fra Gibbs fri energi bare når en kjemisk reaksjon går mot likevekt. Når likevekten først er nådd, kan ikke reaksjonen lenger produsere energi til arbeid. Bioenergetikk
9 9 Endringer i fri energi I en levende celle vil de lokale konsentrasjoner av reaktanter og produkt så godt som aldri være 1M. Så den virkelige G vil ikke være lik G o. Forholdet mellom de to verdiene er G= G o +RT ln [B]/[A] Reaksjon: A B I biokjemiske spor i cellen vil sjelden et produkt (B) bli liggende ubrukt, men vil omsettes videre. Hvis den videre omsetningen er effektiv, kan den cellulære konsentrasjonen av B bli svært lav. Det betyr at ln [B]/[A] kan få en høy negativ verdi, og at G kan bli negativ selv om G o er positiv. Reaksjonen vil derfor kunne gå mot høyre. Bioenergetikk
 ATP + H 2 0 ADP + P i G 0 = 30.5 kj/mol (2) ( G 0 verdiene av sekvensielle reaksjoner er additive.) Glukose + ATP glukose 6-fosfat + ADP G 0 = 16.")
10 10 Endringer i G o er additive Glukose + P i glukose 6-fosfat + H 2 0 G 0 = 13.8 kj/mol (1) (Den omvendte reaksjonen, hydrolyse av G-6-P er spontan.) ATP + H 2 0 ADP + P i G 0 = 30.5 kj/mol (2) ( G 0 verdiene av sekvensielle reaksjoner er additive.) Glukose + ATP glukose 6-fosfat + ADP G 0 = 16.7 kj/mol (3)

11 11 Energien i næringsmolekyl trekkes ut til energibærere som ATP, NADH, NADPH og FADH 2 Exergon / katabolsk reaksjon: systemet frigir fri energi G < 0 Endergon / anabolsk reaksjon: systemet bli tilført fri energi G > 0 Bioenergetikk

12 12 Bindingene mellom fosfatgruppene i adenosin trifosfat (ATP) er meget energirike. ATP er derfor den viktigste bæreren av raskt tilgjengelig energi i cellene. ATP Adenin Energirik binding ATP forbrukes nesten straks det er dannet, slik at det kontinuerlig må erstattes i cellene. Hydrolyse av den terminale fosfatbindingen frigjør 30.5 kj/mol. O C Ribose-sukker N P H Fosfat-grupper Bioenergetikk
 4.")
13 13 Energien som frigjøres ved hydrolyse av ATP skyldes: 1. Redusert ladningsrepulsjon mellom de negative ladningene på ATP. 2. Resonansstabilisering av avspaltet fosfat (P i ) 3. Ionisering av produktet ADP 2- til ADP 3- frigjør H + i miljø med veldig lav [H + ] (~10-7 M / ph~7) 4. Bedre solvatisering av ADP og P i relativt til ATP Fosfat: Fosforyl: -OPO 2-3 -PO 2-3 Det er fosforyl som fjernes fra ATP. Det dannes fosfat etter at O fra H 2 O hektes på. Bioenergetikk

14 14 ATP Den mengden ATP som brytes ned og dannes pr dag hos mennesket, tilsvarer omtrent kroppsvekten Pris på ATP fra Sigma Chemicals: 5g = $296 Det vil si at en aktiv person på 100 kg produserer ATP for ~6 mill $/dag Bioenergetikk

15 15 Energioverføringene i biokjemiske reaksjoner følger de samme termodynamiske lover som ellers i naturen 1. Lov: I enhver fysisk og kjemisk forandring vil den totale energimengden i universet forbli konstant, selv om energiens form kan forandres. 2. Lov: I alle naturlige prosesser vil universets entropi (grad av uorden) øke. Bioenergetikk
16 16 Termodynamikk 1. Termodynamikkens første lov (Konservering av E): Energi kan ikke oppstå eller forsvinne, men den kan forandre form (kjemisk, ) til (fysikalsk, termisk, ). E = q + w 2. Termodynamikkens andre lov (Entropi, S): Entropien i alle naturlige prosesser i universet (men ikke nødvendigvis systemet) vil alltid øke. Total S i universet øker kontinuerlig. S universe = S system + S omg > 0 Bioenergetikk

17 17 Termodynamikkens andre lov (Entropi, S):
18 18 Eks.:Termodynamikkens andre lov (Entropi, S): Oksidasjon av glukose C 6 H 12 O O 2 6CO H 2 O + energi I denne prosessen blir 7 molekyler konvertert til (og tilfeldig delt inn i) 12 nye molekyler. økning i entropi (molekylær uorden). Bioenergetikk

19 19 Oksidasjon Oksidasjon innebærer fjerning av elektroner - enten alene eller i form av hydrogenatomer. Oksidasjoner er nedbrytningseaksjoner (katabolske) som frigjør energi. H H Redusert molekyl Frigitt energi Oksydert molekyl + H H Hydrogenatomer For karbon i karbonforbindelser, vil økt grad av oksidasjon i praksis tilsvare en økning i antall C- O-bindinger Bioenergetikk


20 20 Reduksjon I syntesereaksjoner (anabolske) skjer det motsatte. Forbindelser reduseres ved at elektroner eller hydrogenatomer tas opp. Energi + H H H H Oksydert molekyl Hydrogenatomer Redusert molekyl I cellene skjer oksidasjons- og reduksjonsprosesser samtidig. Når næringsmolekyler oksideres, tas ofte elektronene opp av bestemte elektronbærere, som blir redusert. Disse elektronbærerne kan deretter avgi elektronene til syntesereaksjoner i cellene, blant annet til dannelse av ATP. Bioenergetikk
21 21 Transport av elektroner kan utføre arbeid Når vi kobler et batteri til en motor, vil strømmen av elektroner fra den ene polen til den andre polen kunne få motoren til å utføre arbeid. Tilsvarende vil elektroner som doneres fra ett molekyl til et molekyl med høyere elektronaffinitet (mer stabilt), avgi energi. Denne energien kan benyttes av cellene til å utføre arbeid. Det er dette prinsippet som benyttes for å syntetisere ATP fra de reduserte elektronbærerne NADH og FADH 2. Bioenergetikk
22 22 De viktigste elektronbærerne er NADH, NADPH og FADH 2 Koenzymer: Nikotinamid adenin dinukleotid (NAD/NAD + ) Nikotinamid adenin dinukleotid fosfat (NADP/NADP + ) Fra vitaminet Niacin (Nikotinsyre / Vitamin B 3 ) Prostetisk gruppe: Flavin adenin dinukleotid (FAD/FAD + ) Fra vitamin riboflavin (Vitamin B 2 ) NAD + + 2e - + 2H + NADH + H + NADP + + 2e - + 2H + NADPH + H + (NAD + reduseres til NADH) (NADP + reduseres til NADPH) FAD + + 2e - + 2H + FADH 2 (FAD + reduseres til FADH 2 )
23 23 De viktigste elektronbærerne er NADH, NADPH og FADH 2 (Framside) (Bakside) 1. Substratet donerer 2 H atomer 2. Hvert oksiderte nukleotid aksepterer et hydrid ion (proton + 2 e-) 3. H + frigis til medium Niacinmangel : Pellagra = DDD Bioenergetikk
24 24 De viktigste elektronbærerne er NADH, NADPH og FADH 2 FADH 2 er mer allsidig enn NAD(P)H: 1. Aksepterer 1 eller 2 e- med 1 eller 2 H+ (begge medfører hydrogen overføring) 2. Reduksjonspotensial (-0.4 V til V) avhengig av bindingsstyrke
25 25 Sammendrag Termodynamikkens lover kan anvendes på systemer av levende organismer. Kjemiske reaksjoner er gunstige når den frie energien til produktene er lavere enn den frie energien til reaktantene. Energimessig ugunstige reaksjoner kan gjøres mulig ved å sammenkoble en ugunstig reaksjon med svært gunstige reaksjoner. Oksidasjon/reduksjonsreaksjoner involverer overføring av elektroner fra reduserte organiske molekyler til spesialiserte red/oks kofaktorer. De reduserte kofaktorene kan brukes i biosyntetiske spor eller brukes som energi til ATP syntese. Bioenergetikk
26 26 Stikkord Energi ATP Oksidasjon og reduksjon Massevirkningsloven Elektronbærere
27 27 Det store bildet TCA er en «hub» (sentralstasjon) hvor nedbrytningsreaksjoner (katabolske) konvergerer med oppbygningsreaksjoner (anabolske).

28 28 SIR HANS KREBS Professor i biokjemi (University of Sheffield, ). Vant nobelprisen i medisin/fysiologi i Oppdaget mekanismen for hvordan energi omdannes fra oksidasjon av næringsstoffer i levende celler. Oppdagelsen fikk navnet Krebs syklus eller sitronsyresyklus.
29 29 Sitronsyresyklus, elektrontransportkjeden og oksidativ fosforylering Oksideres og mister e - Når cellene har tilstrekkelig tilgang på oksygen, vil ikke pyruvat dannet i glykolysen omdannes til laktat, men vil i stedet omdannes til Acetyl-CoA og kanaliseres inn i sitronsyresyklus (Krebs syklus). Dette gir en fullstendig nedbrytning av pyruvat til H 2 O og CO 2. O 2 er den endelige elektronakseptoren. e - (energien) konserveres som: Denne aerobe prosessen kalles cellulær respirasjon.


30 30 Oksidativ nedbrytning av pyruvat skjer i mitokondriene
.")
31 31 1 Første trinn er omdanning av pyruvat til Acetyl-CoA Aerobe forhold : det meste av pyruvat dannet i glykolysen transporteres (aktivt) inn i mitokondriene, og oksideres videre gjennom sitronsyresyklus. Bare acetylgruppen i pyruvat går inn i TCA. Denne må først kobles på Coenzym A, og danne acetyl- CoA (=aktivert acetat). Acetylgrupper fra fettsyrer og de fleste aminosyrer kanaliseres også inn i sitronsyresyklus via acetyl-coa. Totalreaksjon for omdanning av pyruvat til Acetyl- CoA ved pyruvat dehydrogenase
 (x60) Dihydrolipoamid dehydrogenase (E3) (x12) I tillegg inneholder komplekset 5 kofaktorer (prostetisk gruppe/koenzym) Fire av kofaktorene stammer fra")
32 32 Pyruvat dehydrogenase (PDH) PDH er det største multienzymkomplekset som er kjent, og består av: Pyruvat dekarboksylase (E1) (x30) (E1 kalles i noen lærebøker for pyruvat dehydrogenase) Dihydrolipoamid acetyltransferase (E2) (x60) Dihydrolipoamid dehydrogenase (E3) (x12) I tillegg inneholder komplekset 5 kofaktorer (prostetisk gruppe/koenzym) Fire av kofaktorene stammer fra vitaminer. Subenhetene er ordnet slik at mellomproduktene i katalysen kan dirigeres fra reaksjon til reaksjon, uten å forlate enzymets overflate Gunstig for regulering Svært godt konservert mellom ulike arter. Elektronmikrografi av pyruvat dehydrogenasekompleks fra E. coli
 Periferi av kompleks: 30")
 12 kopier of E 3 (homodimer - gul) E1 =")

33 33 Pyruvat dehydrogenase (PDH) Kjernekompleks: 60 kopier av E 2 (rød) Periferi av kompleks: 30 kopier av E 1 (tetramer med 2 2 subenheter lilla) 12 kopier of E 3 (homodimer - gul) E1 = Pyruvate dekarboksylase (dehydrogenase) E2 = Dihydrolipoyl transacetylase E3 = Dihydrolipoyl dehydrogenase
34 34 Pyruvat dehydrogenase (PDH) Dekarboksylering Reaktiv thiol-gruppe (-SH) Bindingen mellom CoA og acetylgruppen fra pyruvat kalles en tioester-binding, og er svært energirik - vi sier at den er aktivert. Dette gjør at acetylgruppen kan doneres til en rekke ulike akseptormolekyler. Både arvelige defekter i enzymkomplekset, og vitaminmangel (f. eks fra høyt alkoholinntak) kan føre til tap av PDH funksjon. Enzym Prostetisk gruppe Koenzym Vitamin prekursor E1 Tiamin pyrofosfat (TPP) Tiamin (Vit. B 1 ) E2 Lipoat (lipoamid) CoA-SH Pantotensyre (Vit. B 5 ) E3 FAD Riboflavin (Vit. B 2 ) NAD + Niacin (Vit. B 3 )
 form av lipoat og e - går via FAD til FADH 2 og videre til NADH.")
35 35 Pyruvat dehydrogenase (PDH) oppsummert 1. E1 katalyserer dekarboksylering av pyruvat hydroksyetyl-tpp E2 katalyserer overføring av acetylgruppe til Acetyl-CoA E3 katalyserer regenerering av disulfid (oksidert) form av lipoat og e - går via FAD til FADH 2 og videre til NADH. Den lange lipoyllysin armen svinger fra aktivt sete i E1 til E2 og videre til E3 og knytter intermediatene til enzymkomplekset substratkanalisering
36 36 Acetyl-Coenzym A er et nøkkelintermediat i metabolismen Acetyl-CoA er et energirikt tioester som produseres fra nedbrytning av både glukose, fettsyrer og andre lipider, aminosyrer, karbohydrater mm. Det er derfor det viktigste skjæringspunktet i katabolske og anabolske reaksjoner. I animalske celler, i motsetning til planter, kan ikke acetyl-coa omdannes til karbohydrater. Glukose CO 2 Kolesterol og steroider CoA-SH Heme Acetyl- CoA Fettsyrer Ketogene aminosyrer Ketonlegemer Aktivert gruppe for overføring av energi

37 37 Regulering av pyruvat dehydrogenase Begge produktene Acetyl-CoA og NADH, virker inhiberende på PDH enzymaktiviteten. Aktiviteten reguleres imidlertid også via fosforylering av enzymkomplekset. Fosforyleringen skjer i E1-komplekset, og styres av: Pyruvat dehydrogenase kinase - hemmer PDH -Aktiveres av acetyl-coa, ATP og NADH -Hemmes av pyruvat, CoA og NAD + Pyruvat dehydrogenase fosfatase - aktiverer PDH -Aktiveres av Ca 2+. I skjelettmuskulatur frigjøres Ca 2+ ved trening 5 reaksjoner Intermediatene forlater aldri komplekset (substratkanalisering) Motvirker at andre enzymer stjeler den aktiverte acetylgruppen
38 38 1 Lage Acetyl-CoA: Pyruvat AcetylCoA + CO 2 + NADH 2 Oksidere Acetyl-CoA: Acetyl-CoA 2 CO 2 + FADH NADH + GTP
39 39 2 Sitronsyresyklus (TCA-syklus, Krebs syklus) Acetyl-CoA donerer acetylgruppen inn i sitronsyresyklus, hvor energien i molekylet trekkes ut via en rekke oksidasjons- og dekarboksyleringsreaksjoner (frigjør CO 2 ). Energien som frigjøres, fanges hovedsakelig opp av kofaktorene NAD og FAD. Redusert NADH og FADH 2 benyttes til dannelse av ATP via såkalt oksidativ fosforylering.
40 40 Sitronsyresyklus I sitronsyresyklus doneres 2 karbonatomer inn fra acetyl-coa, og to karbonatomer spaltes av i form av CO 2 i hver runde. Karbonatomene som frigjøres i form av CO 2 er ikke de samme som blir donert av acetyl-coa. I 4 av 8 reaksjoner er energien fra oksidasjon effektivt konservert i form av reduserte kofaktorer (NADH og FADH 2 )
.")

 gjør at dette")
41 41 1 Sitrat syntase starter syklusen Binding av oksaleddiksyre induserer en stor konformasjonsendring i enzymet (grønn=før binding). Dette åpner et bindingssete for acetyl- CoA. Begge molekylene begraves inne i enzymet, og beskyttes mot den ytre løsningen. Dette favoriserer kondensasjonen. Sterkt negativ G 0 (hydrolyse av høyenergi thioesterbinding) gjør at dette er en eksergon reaksjon som dras mot dannelse av sitrat. Dette er viktig for flyten i syklusen.
42 42
 binder til et jernsvovelkompleks som sitter dypt nede i")

.")
43 43 2 Aconitase omorganiserer sitratmolekylet Sitronsyren (sitrat) binder til et jernsvovelkompleks som sitter dypt nede i en kløft på enzymet. Ved substratbinding lukkes enzymet, og Fe 4 S 4 -komplekset deltar i a) fjerning av vann b) tilbakeføring av vann for å danne isositronsyre (isositrat). Reaksjonen er spontan til tross for positiv G 0 pga at isositrat dras inn i neste reaksjon.
44 44

45 45 3 Isositrat dehydrogenase fjerner første CO 2 NAD + Dette trinnet er en oksidativ dekarboksylering. Isositrat oksideres til -ketoglutarat, samtidig som CO 2 spaltes av. Når isositrat oksideres, blir samtidig kofaktoren NAD + redusert til NADH + H +. Det finnes to former av dette enzymet i alle celler, og den ene av disse krever NAD + som e - akseptor og den andre formen krever NADP +. (Hovedfunksjonen til enzymet som foretrekker NADP + er å generere NADPH til reduktive anabolske prosesser.)
46 46
47 47 4 -ketoglutarat dehydrogenase fjerner andre CO 2 -ketoglutarat dehydrogenase er svært likt pyruvat dehydrogenase, og består at tre subenheter E1, E2 og E3. Det har også samme kofaktorer, TPP, lipoat, FAD, NAD + og CoA. Selve reaksjonen er nesten identisk bortsett fra at subenheten E1 binder -ketoglutarat i stedet for pyruvat. Dette trinnet er også en oksidativ dekarboksylering, hvor NAD + reduseres til NADH. Energirik tioesterbinding
48 48

49 49 5 Ravsyre-CoA syntetase danner GTP Energien i tioesterbindingen dannet i forrige trinn er tilstrekkelig til å danne ett molekyl GTP fra GDP. Reaksjonen er reversibel, og enzymet har sitt navn fra den omvendte reaksjonen. Merk at en syntetase foretar kondensasjonsreaksjoner ved bruk av ATP eller GTP, mens en syntase (eks. sitrat syntase) ikke trenger en slik energikilde. Substratnivå fosforylering (Ravsyre thiokinase) GTP + ADP GDP + ATP G ~ 0 kj/mol Effektiv produksjon av 1 GTP/ATP
50 50
. Indre mitokondriemembran Det består av 4 subenheter.")
.")
51 51 6 Ravsyre dehydrogenase danner FADH 2 Ravsyre dehydrogenase er det eneste enzymet i Krebs-syklus som ligger bundet i den indre mitokondriemembranen viktig i elektrontransportkjeden (kompleks II). Indre mitokondriemembran Det består av 4 subenheter. A inneholder FAD som oksiderer ravsyre, B inneholder 3 FeSkompleks, som transporterer elektronene fra FADH 2 til Ubiquinon (Coenzym Q 10 ). Ubiquinon transporterer så e - videre til den endelige e - akseptoren - O 2 (vil bli behandlet under oksidativ fosforylering). Subenhet C og D forankrer enzymet i den indre Kontroversielt i klinisk mitokondriemembranen. sammenheng.

52 52
 til L-malat.")
53 53 7 Fumarase adderer H 2 O Enzymet består av to like subenheter (homodimer), som hver kan binde ett molekyl fumarat. Fumarat hydreres (vann adderes) til L-malat. Stereospesifikt Fumarat
54 54

55 55 8 Malat dehydrogenase danner NADH NAD + Enzymet katalyserer det siste trinnet i sitronsyresyklus. Oksaleddiksyre som dannes blir imidlertid fort omsatt i en ny runde i TCA, og nivået av oksaleddiksyre i cellen er derfor svært lavt ( likevekten dras mot høyre)
56 56 TCA oppsummert Merk: De to karbonatomene som kommer ut som CO 2 er ikke de samme som kom inn via Acetyl-CoA. (I ravsyre og fumarat kan ikke lengre de opprinnelige C-atomene fra acetyl-coa identifiseres pga at de er symmetriske molekyler.) Utbytte av 1 runde: 1 FADH 2, 3 NADH (4 medregnet pyruvat Acetyl-CoA) og 1 GTP (ATP)
57 57 Viktigste reguleringsmåter: 1. Tilgjengelighet av substrat 2. Produktinhibering 3. Allosterisk feedback inhibering Regulering av TCA * * 3 (4) eksergone rx:rxn 1, 3 og 4 a) Alle dehydrogenaser [NADH]/[NAD + ] kan inhiberes av massevirkning : NADH i konkurranse med NAD + for binding til enzym (NAD + er substrat for 3 ulike enzymer) b) Sitrat syntase & -ketoglutarat dehydrogenase Produkt inhibering av sitrat og ravsyre-coa, c) Sitrat syntase, Isositrat & -Ketoglutarat dehydrogenase Allosterisk feedback inhibering av NADH og/eller ATP d) Pyruvat dehydrogenase Acetyl CoA konkurrerer med CoA for å binde seg til E2 * * *

58 58 Mellomprodukter i sitronsyresyklus brukes som utgangspunkt for mange syntesereaksjoner. For at syklusen ikke skal stoppe opp, kan mellomprodukter fylles på på to steder - malat og oksaloacetat. Slike påfyllingsreaksjoner kalles anaplerotiske reaksjoner. TCA: Cellens sentralstasjon Anaplerotiske reaksjoner

59 59 Det store bildet
60 60 1 Lage Acetyl-CoA: Pyruvat AcetylCoA + CO 2 + NADH 2 Oksidere Acetyl-CoA: Acetyl-CoA 2 CO 2 + FADH NADH + GTP 3 Elektrontransport og oksidativ fosforylering: O 2 + e- + ADP + Pi ATP + H 2 O
61 61 Cellulær respirasjon I oksidativ fosforylering konvergerer degradering av karbohydrater, aminosyrer og fett. Oksideres og mister e - Mesteparten av all ATP som dannes av organismer blir syntetisert via: Oksidativ fosforylering (dyr, mikroorganismer) Fotosyntese (planter) Ved cellulær respirasjon blir O 2 redusert til H 2 O vha elektroner donert fra NADH og FADH 2. e - (energien) konserveres som:
62 62 Transport av elektroner kan utføre arbeid Elektroner som doneres fra ett molekyl til et molekyl med høyere elektronaffinitet (mer stabilt), avgir energi. Denne energien kan benyttes av cellene til å utføre arbeid. Det er dette prinsippet som benyttes for å syntetisere ATP fra de reduserte elektronbærerne NADH og FADH 2. Elektronene vandrer mot forbindelser som har høyere standard reduksjonspotensiale. Den frigitte energien benyttes til å transportere protoner (H + ) over indre mitokondriemembran.
 og ytre. Indre membran er ikke permeabel for andre forbindelser enn de som har egne transportører.")
63 63 Mitokondriet Mitokondriet er omgitt av to membraner. Ytre membran er fritt permeabel for små molekyler og ioner (< 5000 Da) via porin. Indre membran har mye større areal enn ytre, foldet i kristae. To funksjonelle rom: indre (matriks) og ytre. Indre membran er ikke permeabel for andre forbindelser enn de som har egne transportører. Proteinene som utfører oksidativ fosforylering sitter i indre membran a) Elektrontransportkjeden b) ATP syntasen

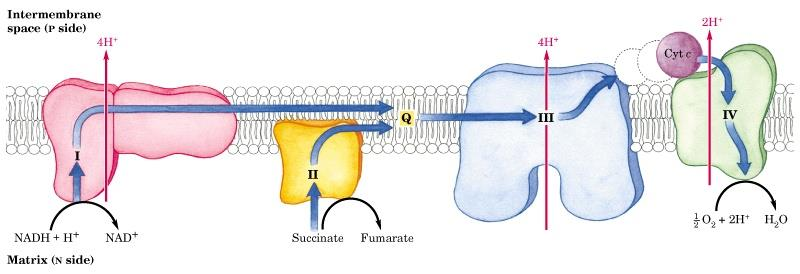
64 64 Elektrontransportkjeden P side = positiv side N side = negativ side
65 65 Elektrontransportkjeden e - overføres fra NADH til flavin mononukleotid (FMN), og via FeS bærere til coenzymq (Q) (Kompleks I) e - fra FADH 2 går inn i elektrontransportkjeden på et lavere energinivå (i forhold til kompleks I). e - overføres til cytokromc i kompleks III, og overføres til kompleks IV. e - brukes til slutt til å redusere O 2 til H 2 O H + kan bare nå inn i matrix via proton-spesifikke kanaler (F o ). Protongradienten som oppstår tilfører nødvendig energi for å danne ATP, og er katalysert av F 1 komplekset som er assosiert med F o.
. 2. Bruke energien som frigjøres til å pumpe 4 protoner ut til intermembranrommet.")
66 66 Kompleks I: NADH-ubiquinon oksidoreduktase (Også kalt NADH dehydrogenase) Dette er et stort proteinkompleks med 42 subenheter. Enzymet har to oppgaver: 1. Overføre ett hydridion fra NADH og ett proton fra matrix til ubiquinon dannelse av ubiquinol (QH 2 ). 2. Bruke energien som frigjøres til å pumpe 4 protoner ut til intermembranrommet. Inneholder flavin mononukleotid (FMN), Fe-S senter, jern-svovel proteinet N-2 og ubiquinon (Q). NADH + H + + Q NAD + + QH 2 (Eksergon)
67 67 Fra ubiquinon (Q) til Ubiquinol (QH 2 ) Komplett reduksjon av ubiquinon til ubiquinol krever to elektroner og to protoner. Ubiquinon er et lite, fettløselig molekyl, som kan flyte fritt inne i mitokondriemembranen, og transportere elektroner mellom de ulike kompleksene.
.")
68 68 Kompleks II: Ravsyre dehydrogenase Det eneste proteinet i TCA syklus som sitter i mitokondriemembranen. 4 ulike subenheter (A-D) og 5 prostetiske grupper (FAD, 3x FeS og Heme b). Elektroner tatt opp fra ravsyre passerer via FADH 2 og 3 FeS-kompleks til ubiquinon ubiquinol (QH 2 ). Heme b beskytter mot reaktive oksygen species (ROS) ved å hindre «lekkasje» av e - (H 2 O 2 O2 - ). Merk at elektroner overført til kompleks II ikke kan bidra til protonpumping i kompleks I. A/B: Forlenging til matrix C/D: Transmembran enhet

69 69 Kompleks III: Cytokrom bc 1 -komplekset Komplekset har 2 bindingsseter for ubiquinol: Q N og Q P 2e - overføres fra: QH 2 2Fe-2S Cytokrom b cytokrom c 1 cytokrom c Redusert cytokrom flyter løst rundt i intermembranrommet og forflytter seg til kompleks IV. Under denne prosessen pumper kompleks III 4 protoner ut til intermembranrommet.

70 70 Kompleks IV: Cytokrom oksidase Dette er det siste trinnet i elektrontransportkjeden. Her overføres to e - fra cytokrom c til to kopperioner bundet til proteinet. Deretter passerer elektronene cytokrom a, og til slutt cytokrom a3 bundet til enda et kopperion. Elektronene tas derfra opp av molekylært oksygen. For hver 4 elektroner som passerer kompleks IV, brukes 4 protoner fra matrix til å danne 2 molekyler vann, samtidig som 4 protoner pumpes ut i intermembranrommet.

71 71 Oppsummering av elektrontransportkjeden For hvert elektronpar som doneres inn via NADH, pumpes 10 protoner ut. For elektronpar som doneres inn direkte til kompleks II fra ravsyre-trinnet i Krebssyklus, pumpes 6 protoner ut.

72 72 Elektrontransportkjeden H + kan bare nå inn i matrix via proton-spesifikke kanaler (F o ). Protongradienten som oppstår tilfører nødvendig energi for å danne ATP, og er katalysert av F 1 komplekset som er assosiert med F o.

73 73 Kjemiosmotisk modell : gradienten er koblet til ATP syntese H + ute = 10X H + inne Elektronenes vandring gjennom elektrontransportkjeden har medført: Frigjøring av energi ved overføring av elektroner til O 2 Transport av protoner ut av matriks Overskudd av elektrokjemisk energi til å drive syntese av ATP.
74 74 ATP syntase (kompleks V) Stort membranintegrert protein. Består av to deler; F o og F 1. F o er en transmembran kanal hvor protoner passerer gjennom. F 1 er et perifert protein på membranens innside. F 1 utfører ATP syntesen. F 1 F 0

. Det dannes 3 ATP pr runde, ca. 100 rotasjoner per sekund!!!")
75 75 F 0 F 1 er en kompleks ATPase Når protoner strømmer gjennom F 0 -delen, roterer denne og hele - skaftet. Dette får de tre aktive setene på -enhetene til å endre konformasjon etter tur. F 1 ADP + P i ATP + H 2 O G 0 0 kj/mol (!) F 0 1.Høy ADP/Pi-binding. 2.Kraftig binding av ATP (forskyver likevekten mot ATP). 3.Svak binding av ATP (ATP frigjøres). Det dannes 3 ATP pr runde, ca. 100 rotasjoner per sekund!!!
76 76 Regulering av oksidativ fosforylering ATP produksjonen reguleres hele tiden for å møte cellens behov. Akseptor kontroll: respirasjonen reguleres av tilgangen på ADP. Massevirkningsforholdet dvs forholdet ATP / (ADP + Pi) regulerer. Dette forholdet er normalt svært høyt dvs nesten alt er ATP. Når cellens aktivitet medfører forbruk av ATP økes tilgangen på ADP og dermed øker respirasjonen. Svært rask regulering sørger for nærmest konstante nivåer av ATP i enhver celle.
77 77 ATP kontrollerer sin egen syntese ATP/ADP ratioen kontrollerer alle viktige reguleringspunkter i katabolismen. ATP kontrollerer dermed sin egen syntese. Når ATP forbruket øker: 1. Øker e - transport og oksidativ fosforylering 2. Øker oksidering av pyruvat (via TCA) 3. Øker hastigheten av glykolysen slik at det dannes mer pyruvat
78 78 Case En bevisstløs 25-årig kvinne blir båret inn på akutten av kjæresten sin. Hun har tatt to slankepiller. Før hun besvimte klagde hun over hodepine, feber, brystsmerter, svette og slapphet. Funn: Temperatur på 40.8⁰C, puls på 151, respirasjonsrate på 56/min, blodtrykk 40/10. Kommentar: Kvinnen døde etter 15 min og kunne ikke gjenopplives. Temp økte til 46 ⁰C 10 min etter at døden hadde inntruffet. Kvinnen drev med fitness og hadde fått kjøpt pillene av en «venn». Disse pillene viste seg å inneholde 2,4 dinitrofenol. 2,4-dinitrofenol er en inhibitor av effektiv energi (ATP) produksjon i celler. Kobler ut oksidativ fosforylering ved å frakte protoner over mitokondriemembranen. Dette fører til rask omsetning av metabolitter og oksygen uten at det genereres noe ATP. All energien brukes til å generere varme. Cellene dør pga høy temperatur og mangel på ATP.

79 79 Mange stoffer kan koble ut elektrontransporten (Inhiberer overføring av e - til jern-svovel senter i kompleks I) Insektmiddel, brukes bl.a mot lakseparasitter (Hemmer oksidasjon av ubiquinol i kompleks III) (Bindes til redusert Fe 2+ i komplex IV)
80 80 Energiutbytte i TCA/elektrontransportkjeden 1 glukosemolekyl: NADH 2 e- via kompleks I, III, IV 10 H + pumpes ut ~ 2.5 ATP FADH 2 2 e- via kompleks II, III, IV 6 H + pumpes ut ~ 1.5 ATP 1 NADH 2.5 ATP (2x4x ATP) 1 FADH ATP (2x1x1.5 3 ATP) 1 GTP 1 ATP (2x1x1 2 ATP) 25 ATP
81 81 Hexose katabolisme - Glykolyse Nettoreaksjon: Glukose + 2NAD + + 2ADP + 2P i 2 Pyruvat + 2NADH + 2H + + 2ATP + 2H 2 O Totalt energiutbytte fra glukosenedbrytning varierer etter hvilken måte redusert NADH fra glykolysen transporteres inn i mitokondriene. To mulige shuttle -mekanismer: 1. Malat-aspartat 2. Glycerol-fosfat

82 82 Malat-aspartat shuttle (Lever, nyre og hjerte) Netto resultat: Overføring av to e - og ett proton til mitokondriematriks, bundet til NADH og klar til videreførsel til elektrontransportkjeden Sitronsyresyklus, elektrontransportkjeden : 2,5 ATP. og oksidativ fosforylering
.")
83 83 Glycerol-fosfat shuttle (Muskel og hjerne) Elektroner fra NADH i cytosol overføres til glycerol-3-fosfat vha glycerol-3-fosfat dehydrogenase (cytoplasma). Glycerol-3-fosfat reoksideres til dihydroksyacetonfosfat av samme enzym (mitokondriemembran). I denne reaksjonen overføres elektronene til FADH 2 og taes med inn i elektrontransportkjeden (kompleks III) : 1,5 ATP.
84 84 Nettoreaksjon fra NADH/FADH 2 til O 2 NADH + H + + ½ O 2 H 2 O + NAD + G o = kj/mol FADH 2 + ½ O 2 H 2 O + FAD G o = kj/mol Sterkt eksergone reaksjoner Energien brukes til å pumpe protoner ut av matriks Overføring av to elektroner fra NADH til O 2 medfører utpumping av 10 protoner => dannelse av 2,5 ATP (lever, nyre, hjerte) Overføring av to elektroner fra FADH 2 til O 2 medfører utpumping av 6 protoner => dannelse av 1,5 ATP (muskel, hjerne)

85 85 Reduserte koenzymer og ATP fra glykolysen, PDH, TCA og oksidativ fosforylering 5-7 ATP 25 ATP
86 86 Glykolysen omsetter bare litt av energien som fins i glukose Glykolyse G = 146 kj/mol 2 GLUKOSE Full oksidasjon (+ 6 O 2 ) G = 2,840 kj/mol 6 CO H 2 O
87 87 Energiutbytte anaerob glykolyse vs aerob TCA/Oksidativ fosforylering Ved anaerobe forhold vil det bli dannet 2 ATP via glykolysen (NADH blir ikke omsatt i TCA). Ved aerobe forhold vil det bli dannet ATP via TCA/oksidativ fosforylering. Mye mer energieffektivt å bryte ned glukose fullstendig til CO 2 og H 2 O fremfor til pyruvat/laktat. Hva er da grunnen til at kreftceller foretrekker å bryte ned glukose til laktat uavhengig av oksygentilgang?
. Hva er grunnen til at kreftceller foretrekker å drive aerob glykolyse?")
88 88 Warburg effekten Kreftceller omdanner glukose til laktat uavhengig av O 2 -tilgang (Otto Warburg i 1924). Hva er grunnen til at kreftceller foretrekker å drive aerob glykolyse? Van der Heiden, Science, 2009
89 89 Warburg effekten Forklaring: Glukoseopptak/glykolyse ~10x raskere i kreftceller sammenlignet med normale celler. En del glykolytiske enzymer er overprodusert i kreftceller. Mange kreftceller har økt evne til å kvitte seg med laktat. Kreftceller har evne til å vokse under hypoksiske forhold (før angiogenese).
 b) Hemmer av")
 2) Diagnostikk: Positron")

90 90 Klinisk betydning: Warburg effekten 1) Mål for terapi: a) Hemmere av hexokinase (2- deoxyglukose, lonidamin og 3- brompyruvat) b) Hemmer av tyrosin kinase (Imatinib) lavere syntese av hexokinase CT PET m/isotop merket glukoseanalog Fusjonert bilde (m/ farge) 2) Diagnostikk: Positron emisjons tomografi (PET) kan brukes til å detektere tumorer vha isotopmerket glukoseanalog PET scanning kan detektere områder med høy glykolyseaktivitet.


91 91 Fedon Lindberg, sukker og kreft "Jo mer sukker og karhohydrater man spiser, desto mer fores kreftcellene." "Kroppen vår er en kompleks biokjemisk suppe. Det er vanskelig å forutsi resultatet av alle komplekse prosesser i menneskekroppen når man utfører tester bare i cellekulturer eller i små gnagere." Viktig å skille mellom årsak og virkning: Ja, kreftceller har høy glykolytisk aktivitet, men det er ikke nødvendigvis årsaken til kreften.
92 92 Oppsummering Relevante læringsmål, Biokjemi: beskrive den generelle strukturen til karbohydrater, lipider, aminosyrer og nukleinsyrer, og forklare stoffenes fysikalsk-kjemiske egenskaper ut fra dette gjøre rede for hovedfunksjonene til karbohydrater, lipider, nukleinsyrer og proteiner beskrive nedbrytingen av glykogen, disakkarider, monosakkarider og fettsyrer, og forklare hvordan slik nedbryting er koplet til dannelse av ATP beskrive syntesen av glukose og glykogen definere begrepene vitamin og mineral, angi de viktigste mineraler og vitaminer og beskrive deres biologiske funksjoner Karbohydrater/Glykolyse/Glukoneogenese/Glykogensyntese/ Glykogenolyse/TCA/Oksidativ fosforylering 1. Fokuser på å forstå hva som skjer og hvorfor Tips: Det er lettere å forstå når man vet litt detaljer 2. Regulering av disse prosessene 3. Sett kunnskapen inn i et klinisk perspektiv
93 93 Youtube; ++
94 94 dimethylisoalloxazine H 3 C C H C C FAD: prostetisk gruppe i E3 i PDH N C O C NH 2 e + 2 H + H 3 C C H C C H N C O C NH H 3 C C C H C N CH 2 C N C O H 3 C C C H C N CH 2 C N H C O HC OH HC OH HC HC OH FAD FADH 2 OH O O Adenine HC HC OH OH O O Adenine H 2 C O P O P O Ribose H 2 C O P O P O Ribose O- O- FAD (Flavin Adenin Dinukleotid) er derivert fra vitaminet riboflavin (Vit. B 2 ). Det er ringsystemet i dimetylisoalloxazin som gjennomgår oksidasjon/reduksjon. FAD er en prostetisk gruppe, permanent bundet til E3. Redusert FADH 2 overfører et hydridion (:H - ) til NAD + NADH som går inn i elektrontransportkjeden. O- O-
Bioenergetikk og Krebs syklus Oksidativ fosforylering
 Bioenergetikk og Krebs syklus Oksidativ fosforylering Bioenergetikk, IA 2015 Det store bildet Bioenergetikk ATP Den mengden ATP som brytes ned og dannes pr dag hos mennesket, tilsvarer omtrent kroppsvekten
Bioenergetikk og Krebs syklus Oksidativ fosforylering Bioenergetikk, IA 2015 Det store bildet Bioenergetikk ATP Den mengden ATP som brytes ned og dannes pr dag hos mennesket, tilsvarer omtrent kroppsvekten
Krebs syklus og Oksidativ fosforylering. Krebs syklus og oksidativ fosforylering, IA 2016
 1 Krebs syklus og Oksidativ fosforylering Krebs syklus og oksidativ fosforylering, IA 2016 2 Relevante læringsmål, Biokjemi: 2.1.1.6 gjøre rede for hovedfunksjonene til karbohydrater, lipider, nukleinsyrer
1 Krebs syklus og Oksidativ fosforylering Krebs syklus og oksidativ fosforylering, IA 2016 2 Relevante læringsmål, Biokjemi: 2.1.1.6 gjøre rede for hovedfunksjonene til karbohydrater, lipider, nukleinsyrer
1 J = cal = energi som trengs for å løfte 1 kg 1m mot en 1N kraft, eller 100 g 1meter mot tyngdekraften (10N) (ett eple en meter)
 (ett eple en meter)") 1 1 J = 0.239 cal = energi som trengs for å løfte 1 kg 1m mot en 1N kraft, eller 100 g 1meter mot tyngdekraften (10N) (ett eple en meter) 2 Energioverføringene i biokjemiske reaksjoner følger de samme
1 1 J = 0.239 cal = energi som trengs for å løfte 1 kg 1m mot en 1N kraft, eller 100 g 1meter mot tyngdekraften (10N) (ett eple en meter) 2 Energioverføringene i biokjemiske reaksjoner følger de samme
4 Viktige termodynamiske definisjoner ΔG = ΔH - T ΔS
 1 2 1 J = 0.239 cal = energi som trengs for å løfte 1 kg 1m mot en 1N kraft, eller 100 g 1meter mot tyngdekraften (10N) (ett eple en meter) Energioverføringene i biokjemiske reaksjoner følger de samme
1 2 1 J = 0.239 cal = energi som trengs for å løfte 1 kg 1m mot en 1N kraft, eller 100 g 1meter mot tyngdekraften (10N) (ett eple en meter) Energioverføringene i biokjemiske reaksjoner følger de samme
Sitronsyresyklus (TCA), elektrontransportkjeden og oksidativ fosforylering. Medisin stadium IA, Tonje S. Steigedal
, elektrontransportkjeden og oksidativ fosforylering. Medisin stadium IA, Tonje S. Steigedal") 1 Sitronsyresyklus (TCA), elektrontransportkjeden og oksidativ fosforylering Medisin stadium IA, Tonje S. Steigedal 2 SIR HANS KREBS and OTTO WARBURG Lindau, 1966 Nobel Prize Awards SIR HANS KREBS Professor
1 Sitronsyresyklus (TCA), elektrontransportkjeden og oksidativ fosforylering Medisin stadium IA, Tonje S. Steigedal 2 SIR HANS KREBS and OTTO WARBURG Lindau, 1966 Nobel Prize Awards SIR HANS KREBS Professor
Pyruvat dehydrogenase er et multienzymkompleks. Oksydativ nebrytning av pyrodruesyre skjer i mitokondriene
 Medisin, stadium 1A, Geir Slupphaug, IKM Sitronsyresyklus Når cellene har tilstrekkelig tilgang på oksygen, vil ikke pyruvat dannet i glykolysen omdannes til laktat, men vil i stedet omdannes til Acetyl-
Medisin, stadium 1A, Geir Slupphaug, IKM Sitronsyresyklus Når cellene har tilstrekkelig tilgang på oksygen, vil ikke pyruvat dannet i glykolysen omdannes til laktat, men vil i stedet omdannes til Acetyl-
Sitronsyresyklus (TCA), elektrontransportkjeden og oksidativ fosforylering
, elektrontransportkjeden og oksidativ fosforylering") 1 Sitronsyresyklus (TA), elektrontransportkjeden og oksidativ fosforylering Medisin stadium IA, Tonje S. Steigedal 2 Relevante læringsmål, Biokjemi: 2.1.1.6 gjøre rede for hovedfunksjonene til karbohydrater,
1 Sitronsyresyklus (TA), elektrontransportkjeden og oksidativ fosforylering Medisin stadium IA, Tonje S. Steigedal 2 Relevante læringsmål, Biokjemi: 2.1.1.6 gjøre rede for hovedfunksjonene til karbohydrater,
Medisin, stadium 1A, Geir Slupphaug, IKM Sitronsyresyklus
 Medisin, stadium 1A, Geir Slupphaug, IKM Sitronsyresyklus Når cellene har tilstrekkelig tilgang på oksygen, vil ikke pyruvat dannet i glykolysen omdannes til laktat, men vil i stedet omdannes til Acetyl-
Medisin, stadium 1A, Geir Slupphaug, IKM Sitronsyresyklus Når cellene har tilstrekkelig tilgang på oksygen, vil ikke pyruvat dannet i glykolysen omdannes til laktat, men vil i stedet omdannes til Acetyl-
Sitronsyresyklus (TCA), elektrontransportkjeden og oksidativ fosforylering
, elektrontransportkjeden og oksidativ fosforylering") 1 Sitronsyresyklus (TA), elektrontransportkjeden og oksidativ fosforylering Medisin stadium IA, Tonje S. Steigedal 2 SIR HANS KREBS and OTTO WARBURG Lindau, 1966 Nobel Prize Awards SIR HANS KREBS Professor
1 Sitronsyresyklus (TA), elektrontransportkjeden og oksidativ fosforylering Medisin stadium IA, Tonje S. Steigedal 2 SIR HANS KREBS and OTTO WARBURG Lindau, 1966 Nobel Prize Awards SIR HANS KREBS Professor
LEHNINGER PRINCIPLES OF BIOCHEMISTRY
 David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 19 Oxidative Phosphorylation 2008 W. H. Freeman and Company Cellulær respirasjon: siste trinn Elektronoverføring
David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 19 Oxidative Phosphorylation 2008 W. H. Freeman and Company Cellulær respirasjon: siste trinn Elektronoverføring
Bioenerge(kk. Bioenerge(kk, IA 2014
 1, IA 2014 2 Store Norske Leksikon: Bioenergetikk, den gren av biokjemien som beskjeftiger seg med de mekanismene som organismene bruker for å skaffe den energien de trenger for å holde livsprosessene
1, IA 2014 2 Store Norske Leksikon: Bioenergetikk, den gren av biokjemien som beskjeftiger seg med de mekanismene som organismene bruker for å skaffe den energien de trenger for å holde livsprosessene
Obligatorisk oppgave 2 MBV1030 Høst 2005
 Obligatorisk oppgave 2 MBV1030 Høst 2005 Levert av (navn): Første del: Flervalgsspørsmål. Angi det svaralternativet (ett) du mener er korrekt. I-1: Ved anaerob glykolyse dannes det laktat. Dersom glukosen
Obligatorisk oppgave 2 MBV1030 Høst 2005 Levert av (navn): Første del: Flervalgsspørsmål. Angi det svaralternativet (ett) du mener er korrekt. I-1: Ved anaerob glykolyse dannes det laktat. Dersom glukosen
Flervalgsoppgaver: celleånding
 Flervalgsoppgaver - celleånding Hver oppgave har ett riktig svaralternativ. Celleånding 1 Nettoutbyttet av glykolysen er pyruvat, 2 ATP og 2 NADH + H + B) 2 pyruvat, 6 ATP og 2 NADH + H + C) 4 pyruvat,
Flervalgsoppgaver - celleånding Hver oppgave har ett riktig svaralternativ. Celleånding 1 Nettoutbyttet av glykolysen er pyruvat, 2 ATP og 2 NADH + H + B) 2 pyruvat, 6 ATP og 2 NADH + H + C) 4 pyruvat,
FLERVALGSOPPGAVER ENERGIOMSETNING
 FLERVALGSOPPGAVER ENERGIOMSETNING FLERVALGSOPPGAVER FRA EKSAMEN I BIOLOGI 2 V2008 - V2011 Disse flervalgsoppgavene er hentet fra eksamen i Biologi 2 del 1. Det er fire (eller fem) svaralternativer i hver
FLERVALGSOPPGAVER ENERGIOMSETNING FLERVALGSOPPGAVER FRA EKSAMEN I BIOLOGI 2 V2008 - V2011 Disse flervalgsoppgavene er hentet fra eksamen i Biologi 2 del 1. Det er fire (eller fem) svaralternativer i hver
Kapittel 7: Cellulære spor for høsting av kjemisk energi
 Kapittel 7: Cellulære spor for høsting av kjemisk energi Glykolyse og cellulær respirasjon Glykolyse Sitronsyresyklus Elektrontransportkjeden med oksydativ fosforylering Aerobisk respirasjon: benyttes
Kapittel 7: Cellulære spor for høsting av kjemisk energi Glykolyse og cellulær respirasjon Glykolyse Sitronsyresyklus Elektrontransportkjeden med oksydativ fosforylering Aerobisk respirasjon: benyttes
Cellular Energetics- Kap. 16
 Forelesninger i BI 212 - Cellebiologi - Våren 2002 Cellular Energetics- Kap. 16 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio chembio.ntnu.no
Forelesninger i BI 212 - Cellebiologi - Våren 2002 Cellular Energetics- Kap. 16 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio chembio.ntnu.no
BIOS 2 Biologi
 BIOS 2 Biologi 2 Figurer kapittel 4: elleåndingen Figur s 107 8 essensielle aminosyrer Tryptofan Metionin Maischips Valin Treonin Fenylalanin Leucin Isoleucin Lysin Bønnedipp Mais og bønner inneholder
BIOS 2 Biologi 2 Figurer kapittel 4: elleåndingen Figur s 107 8 essensielle aminosyrer Tryptofan Metionin Maischips Valin Treonin Fenylalanin Leucin Isoleucin Lysin Bønnedipp Mais og bønner inneholder
SENSORVEILEDNING. Dato: Eventuelt:
 SENSORVEILEDNING Emnekode: HSB1309 Emnenavn: Biokjemi Eksamensform: Skriftlig Dato: 03.06.19 Faglærer(e): Norunn Storbakk Eventuelt: Oppgave 1 a) Tegn og beskriv byggesteiner, oppbygging og strukturen
SENSORVEILEDNING Emnekode: HSB1309 Emnenavn: Biokjemi Eksamensform: Skriftlig Dato: 03.06.19 Faglærer(e): Norunn Storbakk Eventuelt: Oppgave 1 a) Tegn og beskriv byggesteiner, oppbygging og strukturen
Idrett og energiomsetning
 1 Medisin stadium IA, Tonje S. Steigedal 2 ATP er den eneste forbindelsen som kan drive kontraksjon av musklene. ATPnivået i muskelcellene er imidlertid begrenset, og må etterfylles kontinuerlig. Ved ulike
1 Medisin stadium IA, Tonje S. Steigedal 2 ATP er den eneste forbindelsen som kan drive kontraksjon av musklene. ATPnivået i muskelcellene er imidlertid begrenset, og må etterfylles kontinuerlig. Ved ulike
BIOS 2 Biologi
 . Figurer kapittel 2: Energi Figur s. 48 Solenergi Økosystem CO 2 + 2 O Fotosyntese i kloroplaster Organiske molekyler + O 2 Celleånding i mitokondrier Energi til arbeid Varme rodusentene i økosystemet
. Figurer kapittel 2: Energi Figur s. 48 Solenergi Økosystem CO 2 + 2 O Fotosyntese i kloroplaster Organiske molekyler + O 2 Celleånding i mitokondrier Energi til arbeid Varme rodusentene i økosystemet
Biokjemiske reaksjonsruter
 Forelesning i glykolyse. Medisin stadium 1A, Geir Slupphaug, IKM Biokjemiske reaksjonsruter Kan virke svært forvirrende ved første øyekast. Slike oversikter er imidlertid ikke annet enn en form for kart,
Forelesning i glykolyse. Medisin stadium 1A, Geir Slupphaug, IKM Biokjemiske reaksjonsruter Kan virke svært forvirrende ved første øyekast. Slike oversikter er imidlertid ikke annet enn en form for kart,
Kjemisk likevekt. La oss bruke denne reaksjonen som et eksempel når vi belyser likevekt.
 Kjemisk likevekt Dersom vi lar mol H-atomer reager med 1 mol O-atomer så vil vi få 1 mol H O molekyler (som vi har diskutert tidligere). H + 1 O 1 H O Denne reaksjonen er irreversibel, dvs reaksjonen er
Kjemisk likevekt Dersom vi lar mol H-atomer reager med 1 mol O-atomer så vil vi få 1 mol H O molekyler (som vi har diskutert tidligere). H + 1 O 1 H O Denne reaksjonen er irreversibel, dvs reaksjonen er
Kjemien stemmer KJEMI 2
 Figur s. 167 energi aktiveringsenergi uten enzym aktiveringsenergi med enzym substrat produkt reaksjonsforløp Aktiveringsenergien for en reaksjon med enzym er lavere enn for reaksjonen uten enzym. S P
Figur s. 167 energi aktiveringsenergi uten enzym aktiveringsenergi med enzym substrat produkt reaksjonsforløp Aktiveringsenergien for en reaksjon med enzym er lavere enn for reaksjonen uten enzym. S P
Kjemi 2. Figur s Figurer kapittel 8: Biokjemi. Aktiveringsenergien for en reaksjon med enzym er lavere enn for reaksjonen uten enzym.
 Figur s. 177 energi aktiveringsenergi uten enzym aktiveringsenergi med enzym substrat produkt reaksjonsforløp Aktiveringsenergien for en reaksjon med enzym er lavere enn for reaksjonen uten enzym. S P
Figur s. 177 energi aktiveringsenergi uten enzym aktiveringsenergi med enzym substrat produkt reaksjonsforløp Aktiveringsenergien for en reaksjon med enzym er lavere enn for reaksjonen uten enzym. S P
Oksydasjon av glukose og fettsyrer til karbondioksid Dannelse av acetylcoa og sitronsyresyklusen (forts.)
") Oksydasjon av glukose og fettsyrer til karbondioksid Dannelse av acetylcoa og sitronsyresyklusen (forts.) * Det er av betydning for forståelsen av oksydativ fosforylering at man vet hvilke omdannelser
Oksydasjon av glukose og fettsyrer til karbondioksid Dannelse av acetylcoa og sitronsyresyklusen (forts.) * Det er av betydning for forståelsen av oksydativ fosforylering at man vet hvilke omdannelser
Forelesninger i BI Cellebiologi. Enzymer : senker aktiveringsenergien. Figure 6.13
 Enzymer : senker aktiveringsenergien Figure 6.13 Aktive seter : camp-avhengig protein kinase *For å illustrere hvordan det aktive setet binder et spesifikt substrat er valgt som eksempel camp-avhengig
Enzymer : senker aktiveringsenergien Figure 6.13 Aktive seter : camp-avhengig protein kinase *For å illustrere hvordan det aktive setet binder et spesifikt substrat er valgt som eksempel camp-avhengig
UNIVERSITETET I OSLO
 UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen i MBV 1030 Generell biokjemi Eksamensdag: Mandag 6. desember 2004 Tid for eksamen: kl. 09.00 12.00 Oppgavesettet er på 9 sider Vedlegg:
UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen i MBV 1030 Generell biokjemi Eksamensdag: Mandag 6. desember 2004 Tid for eksamen: kl. 09.00 12.00 Oppgavesettet er på 9 sider Vedlegg:
UNIVERSITETET I OSLO. Det matematisk-naturvitenskapelige fakultet
 UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen i MBV 1030 Generell biokjemi Eksamensdag: 6. /7. januar 2005 Tid for eksamen: Oppgavesettet er på 6 sider Vedlegg: 1 Tillatte hjelpemidler:
UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen i MBV 1030 Generell biokjemi Eksamensdag: 6. /7. januar 2005 Tid for eksamen: Oppgavesettet er på 6 sider Vedlegg: 1 Tillatte hjelpemidler:
Vcu. ( K"nto ev-e<ne* - fil, H-oS) UNIVERSITETET I OSLO. Det matemati sk-n aturviten skapelige fakultet. Eksamen i MBV 1030 Generell biokjemi
 UNIVERSITETET I OSLO. Det matemati sk-n aturviten skapelige fakultet. Eksamen i MBV 1030 Generell biokjemi") Vcu UNIVERSITETET I OSLO ( K"nto ev-e
Vcu UNIVERSITETET I OSLO ( K"nto ev-e
Flervalgsoppgaver: Enzymer
 Flervalgsoppgaver - Enzymer Hver oppgave har ett riktig svaralternativ Enzym 1 Et enzym ekstraheres fra Sulfolobus acidocaldarius (en bakterie som finnes i sure, varme kilder med temperaturer opp til 90
Flervalgsoppgaver - Enzymer Hver oppgave har ett riktig svaralternativ Enzym 1 Et enzym ekstraheres fra Sulfolobus acidocaldarius (en bakterie som finnes i sure, varme kilder med temperaturer opp til 90
Biokjemiske reaksjonsruter. Glykolyse. Nedbrytning av glukose til cellulær energi
 Forelesning i glykolyse. Medisin stadium 1A, Geir Slupphaug, IKM Biokjemiske reaksjonsruter Kan virke svært forvirrende ved første øyekast. Slike oversikter er imidlertid ikke annet enn en form for kart,
Forelesning i glykolyse. Medisin stadium 1A, Geir Slupphaug, IKM Biokjemiske reaksjonsruter Kan virke svært forvirrende ved første øyekast. Slike oversikter er imidlertid ikke annet enn en form for kart,
Flervalgsoppgaver: fotosyntese
 Flervalgsoppgaver fotosyntese Hver oppgave har ett riktig svaralternativ. Fotosyntese 1 Hvilke to lysfarger blir best absorbert av klorofyll-a-molekylene? A) rødt og gult B) grønt og blått C) rødt og grønt
Flervalgsoppgaver fotosyntese Hver oppgave har ett riktig svaralternativ. Fotosyntese 1 Hvilke to lysfarger blir best absorbert av klorofyll-a-molekylene? A) rødt og gult B) grønt og blått C) rødt og grønt
FLERVALGSOPPGAVER ENERGIOMSETNING
 FLERVALGSOPPGAVER ENERGIOMSETNING FLERVALGSOPPGAVER FRA EKSAMEN I BIOLOGI 2 Disse flervalgsoppgavene er hentet fra eksamen i Biologi 2 del 1. Det er fire (eller fem) svaralternativer i hver oppgave, og
FLERVALGSOPPGAVER ENERGIOMSETNING FLERVALGSOPPGAVER FRA EKSAMEN I BIOLOGI 2 Disse flervalgsoppgavene er hentet fra eksamen i Biologi 2 del 1. Det er fire (eller fem) svaralternativer i hver oppgave, og
- Kinetisk og potensiell energi Kinetisk energi: Bevegelses energi. Kinetiske energi er avhengig av masse og fart. E kin = ½ mv 2
 Kapittel 6 Termokjemi (repetisjon 1 23.10.03) 1. Energi - Definisjon Energi: Evnen til å utføre arbeid eller produsere varme Energi kan ikke bli dannet eller ødelagt, bare overført mellom ulike former
Kapittel 6 Termokjemi (repetisjon 1 23.10.03) 1. Energi - Definisjon Energi: Evnen til å utføre arbeid eller produsere varme Energi kan ikke bli dannet eller ødelagt, bare overført mellom ulike former
l-l oco UNIVERSITETET IOSLO Det matematisk-naturvitenskapelige fakuftet fi t
 UNIVERSITETET IOSLO Det matematisk-naturvitenskapelige fakuftet Eksamen i: MBV1030 Genercll biokjemi Eksamensdag: S. desember 2006 Tid for eksamen: 15.30 - {9.30 Oppgavesettet er pi 7 side(r) Vedlegg:
UNIVERSITETET IOSLO Det matematisk-naturvitenskapelige fakuftet Eksamen i: MBV1030 Genercll biokjemi Eksamensdag: S. desember 2006 Tid for eksamen: 15.30 - {9.30 Oppgavesettet er pi 7 side(r) Vedlegg:
EKSAMEN I EMNE TBT4102 BIOKJEMI I. 10. desember 2010 kl
 NRGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FR BITEKNLGI Faglig kontakt under eksamen: Institutt for bioteknologi, Gløshaugen Hanne Jørgensen, tlf. 591685 EKSAMEN I EMNE TBT4102 BIKJEMI I 10.
NRGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FR BITEKNLGI Faglig kontakt under eksamen: Institutt for bioteknologi, Gløshaugen Hanne Jørgensen, tlf. 591685 EKSAMEN I EMNE TBT4102 BIKJEMI I 10.
EKSAMEN I EMNE TBT4102 BIOKJEMI I. 2. desember 2011 kl
 NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR BIOTEKNOLOGI Faglig kontakt under eksamen: Institutt for bioteknologi, Gløshaugen Hanne Jørgensen, tlf. 591685 EKSAMEN I EMNE TBT4102 BIOKJEMI
NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR BIOTEKNOLOGI Faglig kontakt under eksamen: Institutt for bioteknologi, Gløshaugen Hanne Jørgensen, tlf. 591685 EKSAMEN I EMNE TBT4102 BIOKJEMI
Velkommen til kjemikurs juni Programfag i studiespesialiserende utdanningsprogram på Vg3
 Velkommen til kjemikurs juni 2008 Programfag i studiespesialiserende utdanningsprogram på Vg3 Program for dagen Presentasjon av Kjemien stemmer for kjemi 2 ved Vivi Ringnes Forsøk for kjemi 2 med utradisjonelt
Velkommen til kjemikurs juni 2008 Programfag i studiespesialiserende utdanningsprogram på Vg3 Program for dagen Presentasjon av Kjemien stemmer for kjemi 2 ved Vivi Ringnes Forsøk for kjemi 2 med utradisjonelt
Glukoneogenese og glykogensyntese
 1 Medisin stadium IA, Tonje S. Steigedal, IKM 2 Relevante læringsmål, Biokjemi: 2.1.1.6 gjøre rede for hovedfunksjonene til karbohydrater, lipider, nukleinsyrer og proteiner 2.1.1.8 beskrive nedbrytingen
1 Medisin stadium IA, Tonje S. Steigedal, IKM 2 Relevante læringsmål, Biokjemi: 2.1.1.6 gjøre rede for hovedfunksjonene til karbohydrater, lipider, nukleinsyrer og proteiner 2.1.1.8 beskrive nedbrytingen
TRANSPORT GJENNOM CELLEMEMBRANEN
 TRANSPORT GJENNOM CELLEMEMBRANEN MÅL: Forklare transport gjennom cellemembranen ved å bruke kunnskap om passive og aktive transportmekanismer Cellemembranen - funksjon - beskytte innholdet i cellen kontroll
TRANSPORT GJENNOM CELLEMEMBRANEN MÅL: Forklare transport gjennom cellemembranen ved å bruke kunnskap om passive og aktive transportmekanismer Cellemembranen - funksjon - beskytte innholdet i cellen kontroll
Glukoneogenese - kroppens egen syntese av glukose
 1 Medisin stadium IA, Tonje S. Steigedal, IKM 2 Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak 1 3 Glukoneogenese Glukoneogenese = dannelse av
1 Medisin stadium IA, Tonje S. Steigedal, IKM 2 Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak 1 3 Glukoneogenese Glukoneogenese = dannelse av
Universitetet i Oslo
 Universitetet i Oslo Det matematisk-naturvitenskapelige fakultet Eksamen i: MBV1030 - Generell biokjemi Eksamenstype: Midtterminseksamen Eksamensdag: Mandag 11. oktober 2004 Tid for eksamen: kl 14.30 17.30
Universitetet i Oslo Det matematisk-naturvitenskapelige fakultet Eksamen i: MBV1030 - Generell biokjemi Eksamenstype: Midtterminseksamen Eksamensdag: Mandag 11. oktober 2004 Tid for eksamen: kl 14.30 17.30
Oppgave 4: Fermentering av karbohydrater og identifisering av disakkaridhydrolaser i gjær
 TBT4107 Biokjemi 2 Oppgave 4: Fermentering av karbohydrater og identifisering av disakkaridhydrolaser i gjær Gruppe 20 Katrine Bringe, Lene Brattsti Dypås og Ove Øyås NTNU, 15. mars 2012 Innhold Sammendrag
TBT4107 Biokjemi 2 Oppgave 4: Fermentering av karbohydrater og identifisering av disakkaridhydrolaser i gjær Gruppe 20 Katrine Bringe, Lene Brattsti Dypås og Ove Øyås NTNU, 15. mars 2012 Innhold Sammendrag
Studie av overføring av kjemisk energi til elektrisk energi og omvendt. Vi snakker om redoks reaksjoner
 Kapittel 19 Elektrokjemi Repetisjon 1 (14.10.02) 1. Kort repetisjon redoks Reduksjon: Når et stoff tar opp elektron Oksidasjon: Når et stoff avgir elektron 2. Elektrokjemiske celler Studie av overføring
Kapittel 19 Elektrokjemi Repetisjon 1 (14.10.02) 1. Kort repetisjon redoks Reduksjon: Når et stoff tar opp elektron Oksidasjon: Når et stoff avgir elektron 2. Elektrokjemiske celler Studie av overføring
... Proteiner og enzymer. kofaktor. polypeptid
 30 Proteiner og enzymer Proteiner er bygd opp av rekker av aminosyrer som er kveilet sammen ved hjelp av bindinger på kryss og tvers, såkalte peptidbindinger. Slike oppkveilete rekker av aminosyrer kaller
30 Proteiner og enzymer Proteiner er bygd opp av rekker av aminosyrer som er kveilet sammen ved hjelp av bindinger på kryss og tvers, såkalte peptidbindinger. Slike oppkveilete rekker av aminosyrer kaller
BIOKJEMI MED BIOTEKNOLOGI
 EKSAMEN BIOKJEMI MED BIOTEKNOLOGI Dato: 22.05.06 Tid: Kl. 09.00-13.00 Antall timer: 4 Antall studiepoeng: 6 Antall sider: 5 (herav 2 vedlegg) Fagansvarlig: Sven Olav Aastad Tillatte hjelpemidler: Kalkulator
EKSAMEN BIOKJEMI MED BIOTEKNOLOGI Dato: 22.05.06 Tid: Kl. 09.00-13.00 Antall timer: 4 Antall studiepoeng: 6 Antall sider: 5 (herav 2 vedlegg) Fagansvarlig: Sven Olav Aastad Tillatte hjelpemidler: Kalkulator
EKSAMENSOPPGAVER uke 3 og 4 fra 2007 til 2001. Noen oppgaver kan gå over i pensum fra uke 13 (integrert metabolisme).
.") EKSAMENSOPPGAVER uke 3 og 4 fra 2007 til 2001. Noen oppgaver kan gå over i pensum fra uke 13 (integrert metabolisme). Kontinuasjonseksamen, MEDSEM2/ODSEM2/ERNSEM2 høst 2007 Onsdag 20. februar 2008 kl.
EKSAMENSOPPGAVER uke 3 og 4 fra 2007 til 2001. Noen oppgaver kan gå over i pensum fra uke 13 (integrert metabolisme). Kontinuasjonseksamen, MEDSEM2/ODSEM2/ERNSEM2 høst 2007 Onsdag 20. februar 2008 kl.
Fasit til oppgavene. K-skallet L-skallet M-skallet
 Kapittel 1 1. Tegn atomet til grunnstoffet svovel (S), og få med antall protoner, nøytroner, elektroner, elektronskall og antall valenselektroner. K-skallet L-skallet M-skallet Svovel har, som vi kan se
Kapittel 1 1. Tegn atomet til grunnstoffet svovel (S), og få med antall protoner, nøytroner, elektroner, elektronskall og antall valenselektroner. K-skallet L-skallet M-skallet Svovel har, som vi kan se
EKSAMEN I EMNE TBT4100 BIOKJEMI GRUNNKURS. 29. november 2007 kl
 NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR BIOTEKNOLOGI Faglig kontakt under eksamen: Institutt for bioteknologi, Gløshaugen Professor Kjell M. Vårum, tlf. 93324 (mob. 930 22165) EKSAMEN
NORGES TEKNISK-NATURVITENSKAPELIGE UNIVERSITET INSTITUTT FOR BIOTEKNOLOGI Faglig kontakt under eksamen: Institutt for bioteknologi, Gløshaugen Professor Kjell M. Vårum, tlf. 93324 (mob. 930 22165) EKSAMEN
Regulering av karbohydratstoffskiftet
 Regulering av karbohydratstoffskiftet DEL I : Oppsummering og oversikt sentrale hormoner og organer i regulering av karbohydratstoffskiftet: Bukspyttkjertel : insulin, glukagon Binyrebark/marg : kortisol/adrenalin
Regulering av karbohydratstoffskiftet DEL I : Oppsummering og oversikt sentrale hormoner og organer i regulering av karbohydratstoffskiftet: Bukspyttkjertel : insulin, glukagon Binyrebark/marg : kortisol/adrenalin
Kjemi og miljø. Elektrokjemi Dette kompendiet dekker følgende kapittel i Rystad & Lauritzen: 10.1, 10.2, 10.3, 10.4 og 10.5
 1 Kjemi og miljø Elektrokjemi Dette kompendiet dekker følgende kapittel i Rystad & Lauritzen: 10.1, 10.2, 10.3, 10.4 og 10.5 Kapittel 10 Elektrokjemi 2 10.1 Repetisjon av viktige begreper: 2 10.2 Elektrokjemiske
1 Kjemi og miljø Elektrokjemi Dette kompendiet dekker følgende kapittel i Rystad & Lauritzen: 10.1, 10.2, 10.3, 10.4 og 10.5 Kapittel 10 Elektrokjemi 2 10.1 Repetisjon av viktige begreper: 2 10.2 Elektrokjemiske
Glukoneogenese. kroppens egen syntese av glukose
 Medisin, stadium A Geir Slupphaug, IKM Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak Glukoneogenese Glukoneogenese = dannelse av nytt sukker Mange
Medisin, stadium A Geir Slupphaug, IKM Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak Glukoneogenese Glukoneogenese = dannelse av nytt sukker Mange
BIOS 1 Biologi
 BIS 1 Biologi..... 1.................... Figurer kapittel 5: Transport gjennom cellemembranen Figur s. 123 glyserol organisk molekyl fosfat glyserol 2 2 2 2 3 R P 2 2 2 2 3 2 2 2 2 3 2 2 2 2 3 2 2 2 2
BIS 1 Biologi..... 1.................... Figurer kapittel 5: Transport gjennom cellemembranen Figur s. 123 glyserol organisk molekyl fosfat glyserol 2 2 2 2 3 R P 2 2 2 2 3 2 2 2 2 3 2 2 2 2 3 2 2 2 2
Forelesninger i BI Cellebiologi. Protein struktur og funksjon - Kap. 3
 Forelesninger i BI 212 - Cellebiologi Protein struktur og funksjon - Kap. 3 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio.ntnu.no Tlf. 73 59
Forelesninger i BI 212 - Cellebiologi Protein struktur og funksjon - Kap. 3 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor-Henning.Iversen@chembio.ntnu.no Tlf. 73 59
Flervalgsoppgaver: proteinsyntese
 Flervalgsoppgaver - proteinsyntese Hver oppgave har ett riktig svaralternativ. Proteinsyntese 1 Hva blir transkribert fra denne DNA sekvensen: 3'-C-C-G-A-A-T-G-T-C-5'? A) 3'-G-G-C-U-U-A-C-A-G-5' B) 3'-G-G-C-T-T-A-C-A-G-5'
Flervalgsoppgaver - proteinsyntese Hver oppgave har ett riktig svaralternativ. Proteinsyntese 1 Hva blir transkribert fra denne DNA sekvensen: 3'-C-C-G-A-A-T-G-T-C-5'? A) 3'-G-G-C-U-U-A-C-A-G-5' B) 3'-G-G-C-T-T-A-C-A-G-5'
FLERVALGSOPPGAVER - CELLEMEMBRANEN
 FLERVALGSOPPGAVER - CELLEMEMBRANEN Hvert spørsmål har ett riktig svaralternativ. Transport cellemembranen 1 På hvilken måte er ulike membraner i en celle forskjellige? A) Fosfolipider finnes bare i enkelte
FLERVALGSOPPGAVER - CELLEMEMBRANEN Hvert spørsmål har ett riktig svaralternativ. Transport cellemembranen 1 På hvilken måte er ulike membraner i en celle forskjellige? A) Fosfolipider finnes bare i enkelte
Oppgave 10 V2008 Hvilket av følgende mineraler er en viktig byggestein i kroppens beinbygning?
 Hovedområde: Ernæring og helse Eksamensoppgaver fra skriftlig eksamen Naturfag (NAT1002). Oppgave 10 V2008 Hvilket av følgende mineraler er en viktig byggestein i kroppens beinbygning? A) natrium B) kalsium
Hovedområde: Ernæring og helse Eksamensoppgaver fra skriftlig eksamen Naturfag (NAT1002). Oppgave 10 V2008 Hvilket av følgende mineraler er en viktig byggestein i kroppens beinbygning? A) natrium B) kalsium
Glukoneogenese og glykogensyntese
 1 Medisin stadium IA, Tonje S. Steigedal, IKM 2 Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak 3 Glukoneogenese = dannelse av nytt sukker Glukoneogenese
1 Medisin stadium IA, Tonje S. Steigedal, IKM 2 Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak 3 Glukoneogenese = dannelse av nytt sukker Glukoneogenese
Lourdes Hydrogenvann maskin. Vann Gården As
 Lourdes Hydrogenvann maskin Vann Gården As S Hydrogen S S S S Er det minste atomet som finner (1 proton 1 elektron) Komponerer 90% av universet En transportør for elektron (bevegelser mellom elektroner
Lourdes Hydrogenvann maskin Vann Gården As S Hydrogen S S S S Er det minste atomet som finner (1 proton 1 elektron) Komponerer 90% av universet En transportør for elektron (bevegelser mellom elektroner
8. Ulike typer korrosjonsvern. Kapittel 10 Elektrokjemi. 1. Repetisjon av noen viktige begreper. 2. Elektrolytiske celler
 1 Kapittel 10 Elektrokjemi 1. Repetisjon av noen viktige begreper 2. Elektrolytiske celler 3. Galvaniske celler (i) Cellepotensial (ii) Reduksjonspotensialet (halvreaksjonspotensial) (iii) Standardhydrogen
1 Kapittel 10 Elektrokjemi 1. Repetisjon av noen viktige begreper 2. Elektrolytiske celler 3. Galvaniske celler (i) Cellepotensial (ii) Reduksjonspotensialet (halvreaksjonspotensial) (iii) Standardhydrogen
Enzymer og enzymkinetikk
 1, IA 2013 2 Relevante læringsmål, Biokjemi: 2.1.1.17 forklare enzymers hovedfunksjon, og definere begrepene K m og V max 2.1.1.18 forklare hvordan enzymers aktivitet kan reguleres i cellene Relevant studiemateriell:
1, IA 2013 2 Relevante læringsmål, Biokjemi: 2.1.1.17 forklare enzymers hovedfunksjon, og definere begrepene K m og V max 2.1.1.18 forklare hvordan enzymers aktivitet kan reguleres i cellene Relevant studiemateriell:
Kapittel 2: Næringsstoffene
 Kapittel 2: Næringsstoffene Tid: 2 skoletimer Oppgave 1 Flervalgsoppgaver a) Hvilke hovedgrupper næringsstoffer gir oss energi? Vann Mineraler Karbohydrater Proteiner Vitaminer Fett b) Hvilket organisk
Kapittel 2: Næringsstoffene Tid: 2 skoletimer Oppgave 1 Flervalgsoppgaver a) Hvilke hovedgrupper næringsstoffer gir oss energi? Vann Mineraler Karbohydrater Proteiner Vitaminer Fett b) Hvilket organisk
FASIT TIL BIOKJEMIEKSAMEN 30. MAI 2005
 FASIT TIL BIOKJEMIEKSAMEN 30. MAI 2005 Oppgave 1 a) Obligat aerobe bakterier må ha oksygen. De utfører aerob respirasjon hvor den endelige elektronakseptoren er oksygen. Fakultativt anaerobe bakterier
FASIT TIL BIOKJEMIEKSAMEN 30. MAI 2005 Oppgave 1 a) Obligat aerobe bakterier må ha oksygen. De utfører aerob respirasjon hvor den endelige elektronakseptoren er oksygen. Fakultativt anaerobe bakterier
Transport av ioner og næringsstoffer over cellemembraner. Læringsmål IA: Beskrive ulike mekanismer for transport over membraner
 Transport av ioner og næringsstoffer over cellemembraner Læringsmål IA: 2.1.2.6 Beskrive ulike mekanismer for transport over membraner Transport Passiv Aktiv Diffusjon Fasilitert diffusjon Ionekanaler
Transport av ioner og næringsstoffer over cellemembraner Læringsmål IA: 2.1.2.6 Beskrive ulike mekanismer for transport over membraner Transport Passiv Aktiv Diffusjon Fasilitert diffusjon Ionekanaler
Oppgave 2b V1983 Hva er et enzym? Forklar hvordan enzymer virker inn på nedbrytningsprosessene.
 Bi2 «Energiomsetning» [2B] Målet for opplæringa er at elevane skal kunne forklare korleis enzym, ATP og andre kofaktorar verkar, og korleis aktiviteten til enzym blir regulert i celler og vev. Oppgave
Bi2 «Energiomsetning» [2B] Målet for opplæringa er at elevane skal kunne forklare korleis enzym, ATP og andre kofaktorar verkar, og korleis aktiviteten til enzym blir regulert i celler og vev. Oppgave
Universitetet i Oslo Det matematisk-naturvitenskapelige fakultet
 Universitetet i Oslo Det matematisk-naturvitenskapelige fakultet Eksamen i KJM1100 Generell kjemi Eksamensdag: Fredag 15. januar 2016 Oppgavesettet består av 17 oppgaver med følgende vekt (også gitt i
Universitetet i Oslo Det matematisk-naturvitenskapelige fakultet Eksamen i KJM1100 Generell kjemi Eksamensdag: Fredag 15. januar 2016 Oppgavesettet består av 17 oppgaver med følgende vekt (også gitt i
Klassifisering av enzymer. Litt historikk. Generell mekanisme for enzymkatalyse:
 Enzymologi De aller fleste enzymer består helt eller delvis av proteiner (unntak: ribozymer) Enzymer har evnen til å katalysere kjemiske reaksjoner i kroppen ved lav temperatur. Selv kjemiske reaksjoner
Enzymologi De aller fleste enzymer består helt eller delvis av proteiner (unntak: ribozymer) Enzymer har evnen til å katalysere kjemiske reaksjoner i kroppen ved lav temperatur. Selv kjemiske reaksjoner
1. UTTAKSPRØVE. til den 44. Internasjonale Kjemiolympiaden 2012. i Washington DC, USA. Oppgaveheftet skal leveres inn sammen med svararket
 Kjemi OL 1 UTTAKSPRØVE til den 44 Internasjonale Kjemiolympiaden 2012 i Washington DC, USA Dag: En dag i ukene 40-42 Varighet: 90 minutter Hjelpemidler: Lommeregner og Tabeller og formler i kjemi Maksimal
Kjemi OL 1 UTTAKSPRØVE til den 44 Internasjonale Kjemiolympiaden 2012 i Washington DC, USA Dag: En dag i ukene 40-42 Varighet: 90 minutter Hjelpemidler: Lommeregner og Tabeller og formler i kjemi Maksimal
BI 212- Protein Sorting - Kap. 17 Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.)
") Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.) Veiene for opptak fra cytosol av kloroplast-proteiner Opptak av proteiner fra cytosol til kloroplaster ligner mye på mitokondrie-importen
Syntese og mål for mitokondrie- og kloroplast-proteiner (forts.) Veiene for opptak fra cytosol av kloroplast-proteiner Opptak av proteiner fra cytosol til kloroplaster ligner mye på mitokondrie-importen
Kapittel 20, introduksjon
 Kapittel 20, introduksjon Ekstracellulær signalisering Syntese Frigjøring Transport Forandring av cellulær metabolisme, funksjon, utvikling (trigga av reseptor-signal komplekset) Fjerning av signalet Signalisering
Kapittel 20, introduksjon Ekstracellulær signalisering Syntese Frigjøring Transport Forandring av cellulær metabolisme, funksjon, utvikling (trigga av reseptor-signal komplekset) Fjerning av signalet Signalisering
Protein Sorting- Kap. 17
 Forelesninger i BI 212 - Cellebiologi - Våren 2002 Protein Sorting- Kap. 17 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor- Henning.Iversen@chembio chembio.ntnu.no Tlf.
Forelesninger i BI 212 - Cellebiologi - Våren 2002 Protein Sorting- Kap. 17 Tor-Henning Iversen, Plantebiosenteret (PBS),Botanisk institutt,ntnu e-mail : Tor- Henning.Iversen@chembio chembio.ntnu.no Tlf.
Faglig kontaktperson under eksamen: Jens Rohloff (mob 97608994)
") Side 1 av 6 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson under eksamen: Jens Rohloff (mob 97608994) EKSAMEN I: BI1001
Side 1 av 6 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson under eksamen: Jens Rohloff (mob 97608994) EKSAMEN I: BI1001
Transport av ioner og næringsstoffer over cellemembranen
 Transport av ioner og næringsstoffer over cellemembranen Terje Espevik, IKM Innhold: Diffusjonshastighet av molekyler over et lipiddobbeltlag Ionekonsentrasjoner innenfor og utenfor en typisk celle Transportere
Transport av ioner og næringsstoffer over cellemembranen Terje Espevik, IKM Innhold: Diffusjonshastighet av molekyler over et lipiddobbeltlag Ionekonsentrasjoner innenfor og utenfor en typisk celle Transportere
3. Balansering av redoksreaksjoner (halvreaksjons metoden)
") Kapittel 4 Oksidasjon og reduksjons reaksjoner (redoks reaksjoner) 1. Definisjon av oksidasjon og reduksjon 2. Oksidasjonstall og regler 3. Balansering av redoksreaksjoner (halvreaksjons metoden) Kapittel
Kapittel 4 Oksidasjon og reduksjons reaksjoner (redoks reaksjoner) 1. Definisjon av oksidasjon og reduksjon 2. Oksidasjonstall og regler 3. Balansering av redoksreaksjoner (halvreaksjons metoden) Kapittel
KJ1042 Øving 3: Varme, arbeid og termodynamikkens første lov
 KJ1042 Øving 3: arme, arbeid og termodynamikkens første lov Ove Øyås Sist endret: 17. mai 2011 Repetisjonsspørsmål 1. Hvordan ser Ideell gasslov ut? Ideell gasslov kan skrives P nrt der P er trykket, volumet,
KJ1042 Øving 3: arme, arbeid og termodynamikkens første lov Ove Øyås Sist endret: 17. mai 2011 Repetisjonsspørsmål 1. Hvordan ser Ideell gasslov ut? Ideell gasslov kan skrives P nrt der P er trykket, volumet,
2. Kjemisk likevekt Vi har kjemisk likevekt når reaksjonen mot høgre og venstre går like fort i en reversibel reaksjon.
 Repetisjon (.09.0) apittel 5 jemisk likevekt. Reversible reaksjoner En reaksjon som kan gå begge veier: H (g) + I (g) HI (g). jemisk likevekt i har kjemisk likevekt når reaksjonen mot høgre og venstre
Repetisjon (.09.0) apittel 5 jemisk likevekt. Reversible reaksjoner En reaksjon som kan gå begge veier: H (g) + I (g) HI (g). jemisk likevekt i har kjemisk likevekt når reaksjonen mot høgre og venstre
De aller fleste enzymer består helt eller delvis av proteiner (unntak: ribozymer) Enzymer har evnen til å katalysere kjemiske reaksjoner i kroppen
 Enzymer har evnen til å katalysere kjemiske reaksjoner i kroppen") Enzymologi De aller fleste enzymer består helt eller delvis av proteiner (unntak: ribozymer) Enzymer har evnen til å katalysere kjemiske reaksjoner i kroppen ved lav temperatur. Selv kjemiske reaksjoner
Enzymologi De aller fleste enzymer består helt eller delvis av proteiner (unntak: ribozymer) Enzymer har evnen til å katalysere kjemiske reaksjoner i kroppen ved lav temperatur. Selv kjemiske reaksjoner
FLERVALGSOPPGAVER ENERGIOMSETNING
 FLERVALGSOPPGAVER ENERGIOMSETNING FLERVALGSOPPGAVER FRA EKSAMEN I BIOLOGI 2 Disse flervalgsoppgavene er hentet fra eksamen i Biologi 2 del 1. Det er fire (eller fem) svaralternativer i hver oppgave, og
FLERVALGSOPPGAVER ENERGIOMSETNING FLERVALGSOPPGAVER FRA EKSAMEN I BIOLOGI 2 Disse flervalgsoppgavene er hentet fra eksamen i Biologi 2 del 1. Det er fire (eller fem) svaralternativer i hver oppgave, og
Introduksjon til Biokjemi. Ingar Leiros, Institutt for Kjemi, UiT
 Introduksjon til Biokjemi Ingar Leiros, Institutt for Kjemi, UiT Biokjemi Biokjemi (Wikipedia): -Studien av de kjemiske prosesser i levende organismer, eller sagt på en annen måte; det molekylære grunnlaget
Introduksjon til Biokjemi Ingar Leiros, Institutt for Kjemi, UiT Biokjemi Biokjemi (Wikipedia): -Studien av de kjemiske prosesser i levende organismer, eller sagt på en annen måte; det molekylære grunnlaget
MED2200-2_OPPGAVE3_V17_ORD DEL 1
 MED2200-2_OPPGAVE3_V17_ORD DEL 1 OVERVEKT OG TYPE 2 DIABETES Kari Larsen er 50 år og har en BMI på 32. Hun har gradvis økt i vekt over de siste 20 årene og har nå merket økt trettbarhet og hyppig vannlating.
MED2200-2_OPPGAVE3_V17_ORD DEL 1 OVERVEKT OG TYPE 2 DIABETES Kari Larsen er 50 år og har en BMI på 32. Hun har gradvis økt i vekt over de siste 20 årene og har nå merket økt trettbarhet og hyppig vannlating.
Auditorieoppgave nr. 1 Svar 45 minutter
 Auditorieoppgave nr. 1 Svar 45 minutter 1 Hvilken ladning har et proton? +1 2 Hvor mange protoner inneholder element nr. 11 Natrium? 11 3 En isotop inneholder 17 protoner og 18 nøytroner. Hva er massetallet?
Auditorieoppgave nr. 1 Svar 45 minutter 1 Hvilken ladning har et proton? +1 2 Hvor mange protoner inneholder element nr. 11 Natrium? 11 3 En isotop inneholder 17 protoner og 18 nøytroner. Hva er massetallet?
Elektrokjemi: Studie av overføring av kjemisk energi til elektrisk energi og omvendt.
 1 Kapittel 10 Elektrokjemi Elektrokjemi: Studie av overføring av kjemisk energi til elektrisk energi og omvendt. 1. Repetisjon av viktige begreper: Reduksjon: Når et stoff tar opp elektron: Cu 2+ + 2e
1 Kapittel 10 Elektrokjemi Elektrokjemi: Studie av overføring av kjemisk energi til elektrisk energi og omvendt. 1. Repetisjon av viktige begreper: Reduksjon: Når et stoff tar opp elektron: Cu 2+ + 2e
Kapittel 12. Brannkjemi. 12.1 Brannfirkanten
 Kapittel 12 Brannkjemi I forbrenningssonen til en brann må det være tilstede en riktig blanding av brensel, oksygen og energi. Videre har forskning vist at dersom det skal kunne skje en forbrenning, må
Kapittel 12 Brannkjemi I forbrenningssonen til en brann må det være tilstede en riktig blanding av brensel, oksygen og energi. Videre har forskning vist at dersom det skal kunne skje en forbrenning, må
Fra alkymi til kjemi. 2.1 Grunnstoffene blir oppdaget
 Fra alkymi til kjemi 2.1 Grunnstoffene blir oppdaget 2.1 Grunnstoffene blir oppdaget GRUNNSTOFF hva er det? År 300 1800: Alkymi læren om å lage gull av andre stoffer Ingen klarte dette. Hvorfor? Teori
Fra alkymi til kjemi 2.1 Grunnstoffene blir oppdaget 2.1 Grunnstoffene blir oppdaget GRUNNSTOFF hva er det? År 300 1800: Alkymi læren om å lage gull av andre stoffer Ingen klarte dette. Hvorfor? Teori
Grunnleggende cellebiologi
 Grunnleggende cellebiologi Ann Kristin Sjaastad Sert. yrkeshygieniker, Dr. Philos HMS-seksjonen, NTNU Tema Cellens oppbygning Transportmekanismer Arvestoff og proteinsyntese Mutasjoner og genotoksisitet
Grunnleggende cellebiologi Ann Kristin Sjaastad Sert. yrkeshygieniker, Dr. Philos HMS-seksjonen, NTNU Tema Cellens oppbygning Transportmekanismer Arvestoff og proteinsyntese Mutasjoner og genotoksisitet
Institutt for biologi Faglig kontaktperson under eksamen: Berit Johansen (91897000) EKSAMEN I: BI1001 Celle- og molekylærbiologi BOKMÅL
 EKSAMEN I: BI1001 Celle- og molekylærbiologi BOKMÅL") 1 av 7 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson under eksamen: Berit Johansen (91897000) EKSAMEN I: BI1001 Celle-
1 av 7 Norges teknisknaturvitenskapelige universitet Fakultet for naturvitenskap og teknologi Institutt for biologi Faglig kontaktperson under eksamen: Berit Johansen (91897000) EKSAMEN I: BI1001 Celle-
Kapittel 4 Ulike kjemiske reaksjoner og støkiometri i løsninger
 Kapittel 4 Ulike kjemiske reaksjoner og støkiometri i løsninger 1. Vann som løsningsmiddel 2. Elektrolytter Sterke elektrolytter Svake elektrolytter Ikke-eletrolytter 3. Sammensetning av løsning Molaritet
Kapittel 4 Ulike kjemiske reaksjoner og støkiometri i løsninger 1. Vann som løsningsmiddel 2. Elektrolytter Sterke elektrolytter Svake elektrolytter Ikke-eletrolytter 3. Sammensetning av løsning Molaritet
1. Oppgaver til atomteori.
 1. Oppgaver til atomteori. 1. Hva er elektronkonfigurasjonen til hydrogen (H)?. Fyll elektroner inn i energidiagrammet slik at du får elektronkonfigurasjonen til hydrogen. p 3. Hva er elektronkonfigurasjonen
1. Oppgaver til atomteori. 1. Hva er elektronkonfigurasjonen til hydrogen (H)?. Fyll elektroner inn i energidiagrammet slik at du får elektronkonfigurasjonen til hydrogen. p 3. Hva er elektronkonfigurasjonen
UNIVERSITETET I OSLO
 Side 1 UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen (utsatt prøve) i: KJM 1110 Organisk kjemi I Eksamensdag: 19. august 2010 Tid for eksamen: 14:30-17:30 Oppgavesettet er på
Side 1 UNIVERSITETET I OSLO Det matematisk-naturvitenskapelige fakultet Eksamen (utsatt prøve) i: KJM 1110 Organisk kjemi I Eksamensdag: 19. august 2010 Tid for eksamen: 14:30-17:30 Oppgavesettet er på
NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK. EKSAMEN I FAG CELLEBIOLOGI 1 august 1997 Tid: kl
 NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK Faglig kontakt under eksamen: Navn: Professor Tore Lindmo Tlf.:93432 EKSAMEN I FAG 74618 CELLEBIOLOGI 1 august 1997 Tid: kl
NORGES TEKNISK-NATURVITENSKAPELIG UNIVERSITET Side 1 av 5 INSTITUTT FOR FYSIKK Faglig kontakt under eksamen: Navn: Professor Tore Lindmo Tlf.:93432 EKSAMEN I FAG 74618 CELLEBIOLOGI 1 august 1997 Tid: kl
3. Massevirkningsloven eller likevektsuttrykk for en likevekt
 apittel 8 jemisk likevekt 1. Reversible reaksjoner. Hva er likevekt? 3. Massevirkningsloven eller likevektsuttrykk for en likevekt 4. Likevektskonstanten (i) Hva sier verdien oss? (ii) Sammenhengen mellom
apittel 8 jemisk likevekt 1. Reversible reaksjoner. Hva er likevekt? 3. Massevirkningsloven eller likevektsuttrykk for en likevekt 4. Likevektskonstanten (i) Hva sier verdien oss? (ii) Sammenhengen mellom
Andre studieår: Institutt Delemneansvarlig
 Innholdsfortegnelse Ex.phil MED-100 Første studieår: Institutt Delemneansvarlig 10 stp 50 stp E-post HEL-FEL (10 studiepoeng) ISM Ellen Pedersen ellen.b.pedersen@uit.no Introduksjon IMB Karen Sørensen
Innholdsfortegnelse Ex.phil MED-100 Første studieår: Institutt Delemneansvarlig 10 stp 50 stp E-post HEL-FEL (10 studiepoeng) ISM Ellen Pedersen ellen.b.pedersen@uit.no Introduksjon IMB Karen Sørensen
Innhold. Forord... 11
 Innhold Forord... 11 Kapittel 1 Atomet og periodesystemet... 13 1.1 Kjemi og atomet... 13 Atomet består av protoner, nøytroner og elektroner... 14 Grunnstoffer... 14 Atomnummer og massenummer... 15 Isotoper...
Innhold Forord... 11 Kapittel 1 Atomet og periodesystemet... 13 1.1 Kjemi og atomet... 13 Atomet består av protoner, nøytroner og elektroner... 14 Grunnstoffer... 14 Atomnummer og massenummer... 15 Isotoper...
+ - 2.1 ELEKTRISK STRØM 2.1 ELEKTRISK STRØM ATOMER
 1 2.1 ELEKTRISK STRØM ATOMER Molekyler er den minste delen av et stoff som har alt som kjennetegner det enkelte stoffet. Vannmolekylet H 2 O består av 2 hydrogenatomer og et oksygenatom. Deles molekylet,
1 2.1 ELEKTRISK STRØM ATOMER Molekyler er den minste delen av et stoff som har alt som kjennetegner det enkelte stoffet. Vannmolekylet H 2 O består av 2 hydrogenatomer og et oksygenatom. Deles molekylet,
Kap 4. Typer av kjemiske reaksjoner og løsningsstøkiometri
 1 Kap 4. Typer av kjemiske reaksjoner og løsningsstøkiometri Vandige løsninger; sterke og svake elektrolytter Sammensetning av løsninger Typer av kjemiske reaksjoner Fellingsreaksjoner (krystallisasjon)
1 Kap 4. Typer av kjemiske reaksjoner og løsningsstøkiometri Vandige løsninger; sterke og svake elektrolytter Sammensetning av løsninger Typer av kjemiske reaksjoner Fellingsreaksjoner (krystallisasjon)
Glukoneogenese - kroppens egen syntese av glukose
 Medisin, stadium A Geir Slupphaug, IKM Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak Glukoneogenese Glukoneogenese = dannelse av nytt sukker Mange
Medisin, stadium A Geir Slupphaug, IKM Glukoneogenese - kroppens egen syntese av glukose Kilder til glukose ved ulike tidspunkt etter matinntak Glukoneogenese Glukoneogenese = dannelse av nytt sukker Mange